Тейнолофы — Teinolophos
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Йинотерии (Yinotheria)
Клада Австралосфениды (Australosphenida)
Надотряд Прототерии (Prototheria)
Отряд Однопроходные (Monotremata)
Семейство †Тейнолофовые (Teinolophidae)
Род †Тейнолофы (Teinolophos)
Вид †Teinolophos trusleri
Тейнолофы (Teinolophos Rich, Vickers-Rich, Constantine, Flannery, Kool et van Klaveren, 1999) — монотипический род монотипического семейства тейнолофововых (Teinolophidae Flannery, Rich, Vickers-Rich, Ziegler, Veatch et Helgen, 2022), происходящий из раннего мела Австралии.
Спекулятивная реконструкция внешнего облика тейнолофа (Teinolophos trusleri).
Единственный установленный вид — Teinolophos trusleri Rich, Vickers-Rich, Constantine, Flannery, Kool et van Klaveren, 1999 (тип). Его остатки обнаружены в местонахождении Флэт-Рок, находящегося на побережье Национального парка Банеронг-Марин на юге штата Виктория, Австралия. Они датируются ранним мелом (формация Вонтагги, конец барремского яруса / низы аптского яруса). На сегодняшний день это древнейшее известное однопроходное.
Родовое название происходит от древнегреческих слов teino — "расширенный" и lophos — "гребень", что указывает на ярко выраженные особенности строения жевательной поверхности коренного зуба. Видовой эпитет присвоен в честь художника П. Траслера за его качественные иллюстрации остатков мезозойских млекопитающих Австралии.

Частичная левая половина нижней челюсти тейнолофа (Teinolophos trusleri).
Материалом для описания таксона послужила неполная левая половина нижней челюсти с сохранившимися мыщелком, угловым отростком и предпоследним коренным зубом. Помимо этого, в наличии имелось 3 нижнечелюстных фрагмента, приписываемые тейнолофу. Так как один из них по своему строению существенно отличается от типового образца, порой его принадлежность тейнолофу оспаривается. В дальнейшем были обнаружены более полные остатки, включая хорошо сохранившуюся левую ветвь нижней челюсти.
Расчетная длина черепа составляет 2,5 см, длина тела без учета хвоста — около 10 см, а вес оценивается в 40 г. Это делает тейнолофа самым мелким известным однопроходным.
В целом животное демонстрирует лишь немного более продвинутые черты, чем морганукодонты (Morganucodonta) и докодонты (Docodonta).
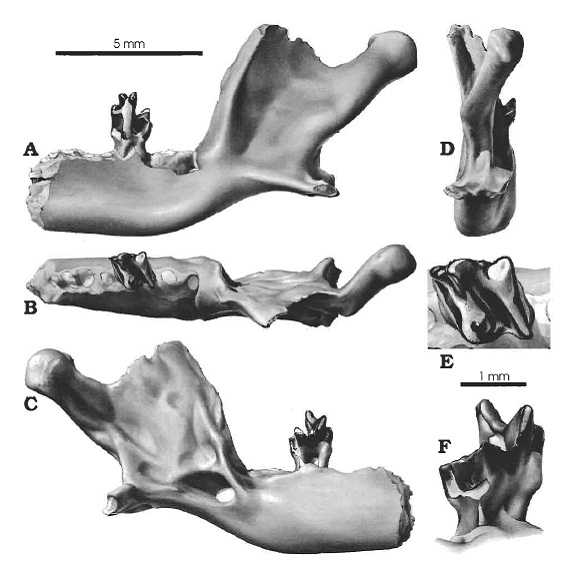
Прорисовки частичной левой половины нижней челюсти и коренного зуба тейнолофа (Teinolophos trusleri): A — снаружи; B — сверху; C — изнутри; D — сзади; E — вид сверху на предпоследний коренной зуб; F — наклонно-щечной вид предпоследнего коренного зуба.
По своему строению нижняя челюсть существенно отличается от таковой у других однопроходных. Среди ярко выраженных различий — суставный мыщелок, расположенный значительно выше горизонтальной ветви зубной кости (а не примерно на одном уровне с ней), высокая восходящая ветвь, а также гребни, расположенные впереди переднего края массетерной ямки с наружной и внутренней стороны. Хорошо развитый угловой отросток обладает внутренним и наружным выступами, тогда как у утконосовых и ехидновых маленький угловой отросток, когда он присутствует, несет выступ только с внутренней стороны. Все эти особенности предполагают, что жевательная и крыловидная мышцы были довольно развитыми, а укус являлся гораздо более мощным и имел иную механику, чем у современных однопроходных с их слабыми челюстями.
У базальных млекопитающих, в отличие о тейнолофа, наличествуют более низко расположенный мыщелок и менее развитый угол нижней челюсти, в совокупности с преобладанием действия височной мышцы. Впрочем, массетерный гребень и хорошо развитые места для прикрепления крыловидной мышцы имеются в угловой области челюсти и у некоторых других мезозойских млекопитающих — например, триконодонтовых (Triconodontidae) и спалакотериевых (Spalacotheriidae), несмотря на то, что их мыщелок располагается примерно на уровня зубного ряда.
Внутренний край нижнечелюстного отверстия тейнолофа следует за вершиной задневнутреннего челюстного угла, в связи с чем большая часть углового выступа в нижнечелюстной линии гипотетична. Кроме того, предполагаемые общие черты угловых отростков тейнолофа и тринаксодона (Thrinaxodon liorhinus) также проблематичны, поскольку у последнего все зазубные кости располагаются с внутренней стороны и нижнечелюстного отверстия на уровне углового отростка нет. Определение нижнечелюстной ложбинки для размещения зазубных костей у тейнолофа тоже проблематично. Глубокая впадина, направленная к мыщелку, с выраженным срединным гребнем — характерный для млекопитающих признак, который у тейнолофа неизвестен; отдельные выступы и гребни в ложбинке также отсутствуют.
Зубная кость по всей длине прободена относительно большим каналом для прохождения ветви тройничного нерва, которая у утконосов передает сигналы от рецепторов клюва в соматосенсорную кору головного мозга. Таким образом, у тейнолофа в определенной степени мог быть развит клюв.
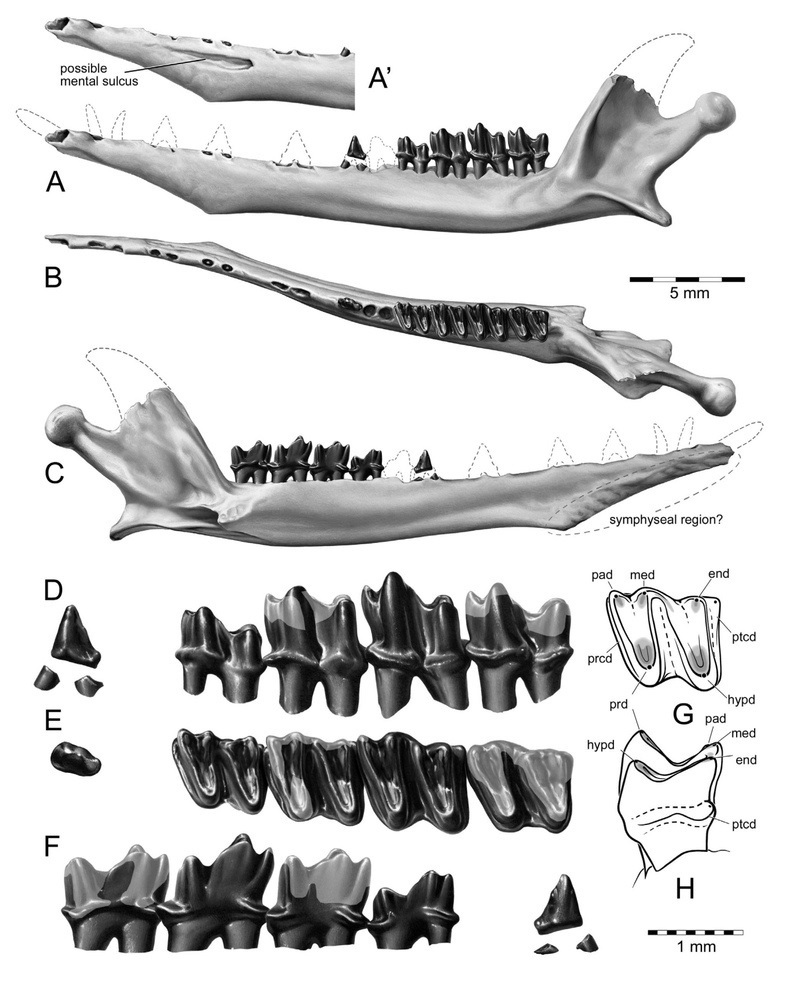
Прорисовки левой ветви нижней челюсти тейнолофа (Teinolophos trusleri) и ее зубов.
В наличном ископаемом материале по тейнолофу недостает подробностей, указывающих на строение косточек его среднего уха. Однако наличие длинной и изогнутой ложбинки на внутренней поверхности нижней челюсти может указывать на то, что, в отличие от современных однопроходных, у которых нижняя челюсть состоит из единственной зубной кости и цепь слуховых косточек в общем сходны с таковыми у сумчатых и плацентарных, у тейнолофа в состав нижней челюсти продолжали входить угловая и предсочленовная кости. Это свидетельствует о том, что сегодняшнее состояние среднего уха у однопроходных и териев сформировалось независимо друг от друга.
Поскольку зубную коронку голотипа покрывала твердая порода, бугорок в ее постеролингвальной области сначала приняли за метаконид, а не гипоконулид, вследствие чего тейнолоф был определен в качестве эвпантотерия. После расчистки зуба стало очевидным, что в передней части он несет триакидидрепанид (тригонид), а в задней — диакидрепанид (талонид). Это придает зубу сходство с m2 стероподона (Steropodon galmani) и, в меньшей степени, с коренными обдуродонов (Obdurodon). Дальнейшее исследование указало и на другие черты, общие со стероподоном. В связи с этим тейнолоф был предварительно зачислен в семейство стероподонтовых (Steropodontidae), что может быть оспорено в связи с появлением нового ископаемого материала и увеличения числа сравниваемых признаков.
Нижний коренной голотипа примерно в 6 раз меньше, чем m2 стероподона. Его отличие от последнего состоит в его значительно большей антеропостериорной сжатости, меньшей мезиодистальной удлиненности, приблизительно равной длине и ширине зуба (в отличие от длины, превышающей ширину), более мезиодистально сжатом тригониде, более косо расположенных обеих тригонидах и талониде, а также в значительно меньших мезиальном и дистальном цингулидах. Талонидный бассейн отсутствует. Уникальной особенностью тейнолофа и других озубленных однопроходных является то, что их тригонид высок, а талонид расположен гораздо ниже. У зуба имеется всего 2 корня, тогда как у утконосовых (в частности, у обдуродонов) они множественны.
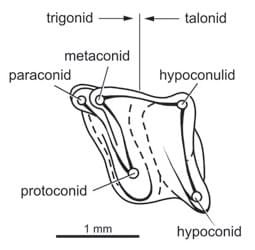
Схема жевательной поверхности нижнего коренного зуба тейнолофа (Teinolophos trusleri).
Износ тригонида и талонида подобен таковому у стероподона, что свидетельствует о нетрибосфеническом типе жевательной поверхности, так как отсутствуют признаки протокона, закрывавшего талонидный бассейн. Зоны износа талонида расположены вне центральной части вместо расположения внутри скошенных стенок бассейна, что указывает на "ортальное" смыкание с верхними коренными. У стероподона и обдуродонов главная ось тригонида почти перпендикулярна медиальным и лабиальным сторонам зуба, а также продольной оси челюсти. У тейнолофа, напротив, эта линия формирует сравнительно острый угол, что означает возможное отсутствие медиалатерального движения челюсти после смыкания поверхностей. Поэтому движение челюстей, скорее всего, осуществлялось почти строго прямо. Это подтверждает почти вертикальный износ зубов стероподона.
Остатки тейнолофа найдены в прибрежных морских отложениях. Судя по всему, он жил в условиях довольно прохладного климата (горные породы содержат следы низкотемпературных воздействий) и питался мелкими беспозвоночными животными.
Ссылки
Teinolophos (Paleobiology Database) перейти
Teinolophos (Wikipedia) перейти
Литература
Grant T. The platypus: a unique mammal. — 4nd ed. — Collingwood: CSIRO Publishing, 2007
Kielan-Jaworowska Z., Cifelli R. L., Luo Z-X. Mammals from the age of dinosaurs: origins, evolution, and structure. — New York: Columbia University Press, 2004
Long J., Archer M., Flannery T., Hand S. Prehistoric mammals of Australia and New Guinea: one hundred million years of evolution. — Baltimore: Johns Hopkins University Press, 2002
Musser A. M. Furry Egg-layers: Monotreme Relationships and Radiations. In Merrick J. R., Archer M., Hickey G. M., Lee M. S. Y. (eds). Evolution and Biogeography of Australasian Vertebrates. — Sydney: Australian Scientific Publishing, 2006
Nowak R. M. Walker’s Mammals of the World. Monotremes, Marsupials, Afrotherians, Xenarthrans, and Sundatherians. — Baltimore: Johns Hopkins University Press, 2018
Rich T. H., Vickers-Rich P., Trusler P., Flannery T. F., Cifelli R., Constantine A., Van Klaveren L. K. N. Monotreme nature of the Australian Early Cretaceous mammal Teinolophos. Acta Palaeontologica Polonica 46 (1), 2001
Rose K. D. The beginning of the age of mammals. — Baltimore: The Johns Hopkins University Press, 2006