Тероцефалы — Therocephalia
Класс †Синапсиды (Synapsida)
Подкласс †Терапсиды (Therapsida)
Надотряд †Терозавры (Therosauria)
Отряд †Тероцефалы (Therocephalia)
Тероцефалы (Therocephalia Broom, 1903) — отряд надотряда терозавров (Therosauria Kemp, 1982), происходящая из средней перми - среднего триаса Африки, средней перми - среднего триаса Европы, поздней перми - среднего триаса Азии, раннего триаса Южной Америки и раннего триаса Антарктики. Установлено около 21 семейства.
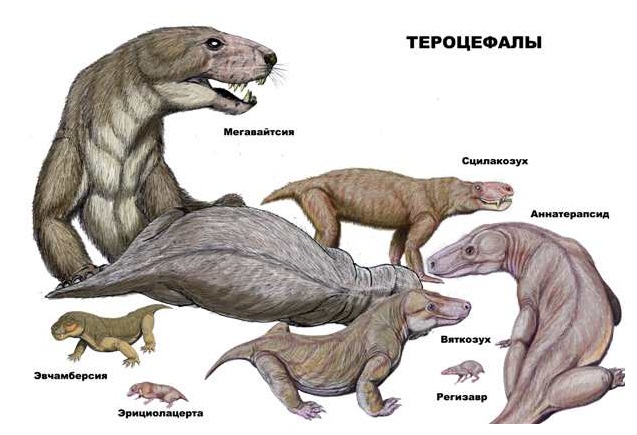
Разнообразие тероцефалов (Therocephalia). Автор — Д. Богданов.
С древнегреческого языка название группы переводится как "звероголовые".
Размеры небольшие и средние. Весьма разнообразная группа, во многом параллельная цинодонтам.
Череп массивный, обычно несколько уплощенный. Морда умеренно удлиненная, широкая, низкая. Скуловая и глазничные дуги обычно узкие. У мелких продвинутых тероцефалов — Bauriidae — глазница неполная (как и у млекопитающих, ее задний край редуцирован). Отросток чешуйчатой кости не заходит далеко вперед по скуловой дуге. Предтеменной кости нет, заднелобная обычно сохраняется. Теменные кости узкие, широко разделяющие заглазничные и чешуйчатые кости и образующие главную часть верхнего края височных окон. Теменные кости обычно образуют высокий сагиттальный гребень, который отсутствует только у примитивного семейства Scylacosauridae. У примитивных родов пинеальное отверстие маленькое, у более продвинутых исчезает. Затылок в большинстве случаев широкий, низкий и вогнутый. Дистальный конец околозатылочного отростка зачастую окостеневает не полностью. У большинства тероцефалов затылочный мыщелок непарный, образованный основными затылочными костями при участии боковых затылочных костей, у продвинутых Baurioidea — глубоко двухраздельный или отчетливо парный.
Квадратная и квадратно-скуловая кости маленькие. Высказываются предположения о подвижности у тероцефалов квадратной кости, что оспаривается другими исследователями. Действительно, наблюдается ослабление связи комплекса квадратной и квадратно-скуловой кости с черепом. Квадратно-скуловая кость теряет связь со скуловой, а весь комплекс входит в вогнутость на задней поверхности чешуйчатой кости. Но в известных случаях квадратно-скуловая кость по верхнему краю сочленяется зубчатым швом с прилежащим участком переднего края чешуйчатой кости, чего не наблюдалось у горгонопов с подвижной квадратной костью. Как и у горгонопов, у тероцефалов представлена лигаментозная связь квадратной кости с квадратными ветвями птеригоидов, а сочленение самой квадратной кости с чешуйчатой костью (по крайней мере, у Annatherapsidus), упрочнено вертикальным гребнем чешуйчатой кости, который не позволяет квадратной кости смещаться. В отличие от горгонопид, челюстная мускулатура ранних тероцефалов распространялась дорсально на мозговую коробку, оставляя лишь узкий сагиттальный гребень между аддукторными камерами.
Основание черепа короткое. Базисфеноидные бугры умеренно развиты. Рострум парасфеноида с вентральным килем, заходящим далеко назад, в область базиптеригоидного сочленения. Сфенэтмоид имеется. Проотика с передней пластиной, имеющей глубокую вырезку для прохождения челюстных ветвей тройничного нерва. В носовой полости развиты турбиналии.
У многих тероцефалов есть вторичное костное небо, но, в отличие от цинодонтов, в его образовании участвуют сошник, премаксиллы и максиллы. По сути, данное образование представляет собой прехоанальный мостик между максиллами и сошником. Небные кости остаются дорсальнее и в состав вторичного неба не входит. Вторичное небо имеется у Bauriodea, более ограниченный контакт между максиллой и сошником возникает также у Moschowhaitsia, Regisaurus и Urumchia. Обычно хорошо развиты парахоанальные гребни небных костей, ограничивающие постхоанальный небный носоглоточный желоб. Сошник парный, обычно суженный посередине и резко расширенный на переднем конце. На небе почти всегда развиты большие подглазничные окна, утраченные лишь у Whatsiidae и Euchambersia. Высказано предположение, что именно наличие больших окон не позволило тероцефалам развить ушной звукопроводящий аппарат по типу млекопитающих.
Обычно имеется щелевидная межптеригоидная яма, находящаяся позади уровня соприкасающихся поперечных отростков птеригоидов. С основанием черепа птеригоиды сочленяются посредством треугольных пластинок, расположенных на квадратных ветвях. Верхняя крыловидная кость расширенная, но не образует шовного соединения с передней ушной костью. Как и у цинодонтов, надкрыловидная кость простирается в виде костной пластины латеральнее мозговой коробки. Вырезка для челюстной мускулатуры на вентральной поверхности черепа заходит далеко вперед кнаружи от эктоптеригоида.
Как и у горгонопид, квадратные ветви птеригоидов отходят непосредственно сзади от базиптеригоидного сочленения, которое у известных в этом отношении тероцефалов неподвижно. Квадратная ветвь птеригоида (и эпиптеригоида) обычно несколько обособлена от собственно базиптеригоидного отростка. Этот отросток образован базикраниальной пластиной, широко соединяющейся с основанием квадратной ветви и сзади обособленной от последней вырезкой. Эпиптеригоиды у низших тероцефалов Scylacosauridae с узким, как у горгонопид, восходящим отростком. У остальных тероцефалов восходящая пластина эпиптеригоидов расширенная.
Небные зубы развиты только на птеригоидах, тогда как зубы на небных костях всегда отсутствуют. У примитивных тероцелафов небные зубы представлены 3 основными рядами: антеролатеральным и антеромедиальным на птеригоидах, а также зубами на поперечных птеригоидных флангах. Антеромедиальная группа обычно замещается парасагиттальным костным гребнем, зубы на поперечных отростках в подавляющем большинстве случаях также утрачиваются, но зубы антеролатеральной группы исчезают лишь у специализированных форм.
Нижняя челюсть высокая, но со слабым симфизом. Зубная кость значительно увеличена, но прочие кости нижней челюсти также хорошо развиты. Зубная кость с небольшим венечным отростком, напоминающим горгонопидный, но имеющим не закругленный, а почти прямоугольный верхний край. Между зубной и угловой костями в большинстве случаев имеется щелевидное нижнечелюстное окно. Задний край ангулярного лепестка почти вплотную приближен к заднему краю челюсти, его наружная поверхность с крестообразным гребнем. Ретроартикулярный отросток нижней челюсти направлен вниз, но нижний его конец не загнут вперед. Стремя не прободенное, во всяком случае, заметного отверстия в нем нет.
В верхней челюсти имеются 7 сильно развитых резцов, 1-2 больших клыка и не более 9 слабых заклыковых зубов. 2 пары верхних клыков характерны для примитивных форм (Lycosuchidae, Trochosuchidae). Впрочем, их наличие в настоящее время подвергается сомнению — не исключено, что за дополнительный функционирующий клык принимается замещающий клык, обычно скрытый в толще верхнечелюстной кости. Замещающий клык полностью вырастал заранее, еще до выпадения старого, и какое-то время в пасти имелось сразу 4 верхних клыка. Затем старый клык выпадал, а рядом с оставшимся начинал расти новый сменный клык. Среди челюстных костей Lycosuchidae почти 40 % находок — с удвоенными клыками. Ранние тероцефалы (например, Gorynychus) имеют тонкие клыки с режущими гранями, которые требовались для вырезания из добычи кусков плоти. У типичных тероцефалов клыки становятся округлыми в сечении и служили для разрывания жертвы.
Заклыковые зубы простые конические, без придаточных бугорков, иногда с продольными гранями на поверхности у примитивных тероцефалов, у Ictidosuchidae они могут становиться трехзубчатыми, а у Baurioidea приобретают уплощенные коронки жующего типа. У примитивных тероцефалов может сохраняться до 9 заклыковых зубов, но в типичных случаях их число уменьшено, а у Whaitsiidae и Euchambersia они полностью исчезают.
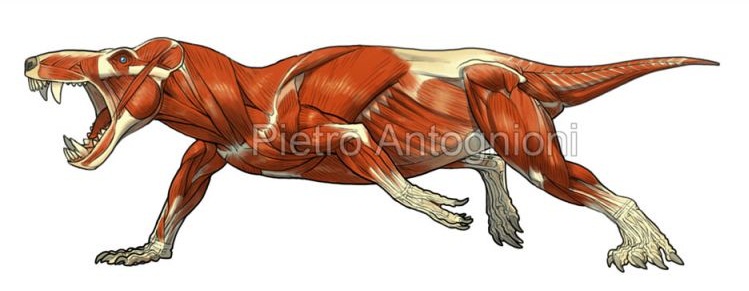
Реконструкция скелетной мускулатуры Pristerognathus.
Телесные пропорции зачастую сходны с таковыми у млекопитающих. Посткраниальный скелет у некоторых видов массивный. Как у горгонопид, атлант примитивного темноспондильного типа состоит из парных невральных дуг и самостоятельных интер- и плевроцентра. Невральные дуги атланта тесно контактируют с плевроцентром последнего.У бауриоидов плевроцентр атланта тесно связан с невральными дугами, что маскирует темноспондильность его состава. Нет никакой тенденции к объединению плевроцентра атланта с эпистрофеем, так что эпистрофеевый комплекс в шее не сформирован. Поясничные ребра уменьшены.
В плечевом поясе имеется клейтрум. Лопатка со слабым акромиальным отростком. Тироидное отверстие мало или отсутствует. Плечевая кость с энтэпикондилярным отверстием. Тазовый пояс пластинчатый, но с глубокими вырезками между подвздошной костью и обеими нижними костями таза. У ряда форм (Regisaurus, Ericiolacerta и др.) седалищная кость удлинена назад, что может предполагать тенденцию перехода к парасагиттальной постановке задней пары конечностей. Конечности зачастую длинные. Передняя их пара обычно широко расставлена. В изученных случаях фаланговая формула маммального типа: 2 - 3 - 3 - 3 - 3. Хвост обычно в большей или меньшей степени укорочен.

Портретные реконструкции тероцефалов: 1 — Lycosuchus; 2 — Scylacosaurus; 3 — Moschorhinus; 4 — Ictidosuchoides. Автор — Zero Smany.
Вероятно, у тероцефалов имелась влажная мочка носа, обостряющая обоняние и позволяющая определять направление ветра. Об этом свидетельствуют отпечатки желез возле ноздрей. Наличие турбиналей также может свидетельствовать о хорошем обонянии. Отпечатки сосудов и мелкие ямки на черепах показывают, что эти животные имели вибриссы, хотя мягкие губы не были достаточно развиты. Таким образом, сосать молоко в младенчестве они не могли. Тем не менее, детеныши вполне могли слизывать молоко с железистых полей на брюхе матери. Наружные ушные раковины, судя по всему, отсутствовали.
Тело почти наверняка покрывал шерстный покров. В отложениях терминальной перми местонахождения Вязники (Владимирская область, Россия) обнаружены копролиты каких-то терапсид — скорее всего, тероцефалов. В них содержатся шерстинки — то ли тероцефалы себя вылизывали и глотали шерсть, то ли поедали добычу покрытую шерстью.
Основная масса тероцефалов описана из Южной Африки, но довольно обильные остатки происходят также из Восточной Африки, Европейской части России и Западного Китая.

Портретные реконструкции тероцефалов. Автор — Г. Угуэто.
Большинство тероцефалов — хищники. Их добычей могло быть любое животное мельче их самих или близкое по размерам. Некоторые тероцефалы обладали ядовитым укусом. Существовали также насекомоядные, рыбоядные, падалеядные, всеядные и, возможно, сугубо растительноядные формы.
Судя по строению склеротикальных колец, большинство тероцефалов предпочитало сумерки, однако среди них также имелись дневные и ночные формы. Однако надо заметить, что мелкие тероцефалы жили в норах — поэтому их зрение могло быть приспособлено к подземному сумраку.
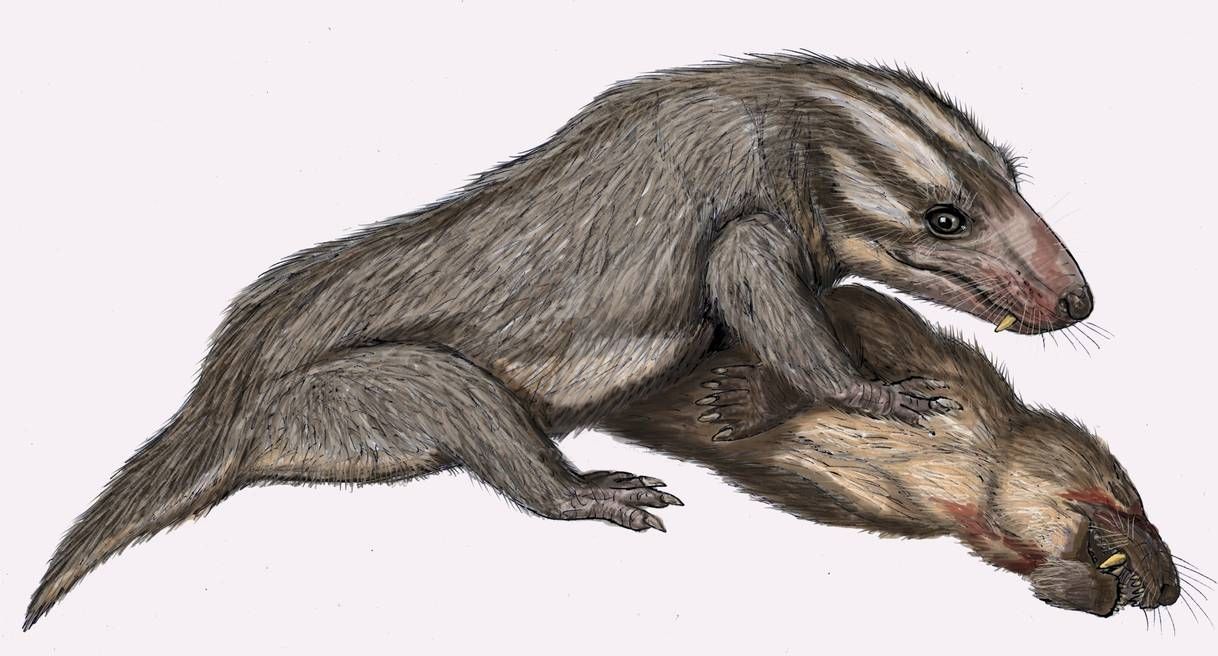
Плотоядный тероцефал мосховайтсия (Moschowhaitsia vjuschkovi) с добытым растительноядным тероцефалом пурловией (Purlovia maxima). Автор — Д. Богданов.
Вероятно, тероцефалы возникли в Африке, где известны уже из самых ранних фаун. В конце средней перми, после вымирания диноцефалов, они некоторое время играли роль доминирующих наземных хищников. Вскоре верширну трофической пирамиды заняли горгонопиды, зачастую достигавшие весьма значительной величины. Примечательно, что смена доминирующих хищников в Восточной Европе и Южной Африке развивалась схожим образом, несмотря на большое расстояние между регионами: в пермском периоде они располагались на разных краях Пангеи. Тероцефалам надолго пришлось довольствоваться нишами средне- и мелкоразмерных хищников. Все-таки, некоторые позднепермские представители группы продолжали довольно успешно конкурировать с горгонопидами. Как и последние, эти тероцефалы обладали мощными клыками и демонстрировали тенденцию к утрате заклыковых зубов.

Мегавайтсия (Megawhaitsia patrichae) возле тела амфибии Dvinosaurus. Автор — А. Атучин.
В терминальной перми, когда горгонопиды в ряде регионов планеты исчезли, вакантные ниши крупных хищников опять ненадолго заняли увеличившиеся в размерах тероцефалы (пример — Вязниковская фауна в Приуралье). В триасовом периоде их сместили уже принципиально другие хищники — рептилии архозавры. В начале триаса выделилась линия менее крупных тероцефалов, попытавшихся освоить растительный рацион. Последние специализированные представители группы вымерли в среднем триасе, возможно, не выдержав конкуренции с цинодонтами с одной стороны и бурно эволюционировавшими архозаврами с другой.

Тероцефал мосхорин (Moschorhinus kitchingi) возле убитого им дицинодонта Lystrosaurus и группа тероцефалов тетрацинодонов (Tetracynodon darti). Автор — Г. Угуэто.
Прежде тероцефалов считали возможными предками цинодонтов или даже предками млекопитающих. Однако несмотря на то, что поздние тероцефалы приобретают ряд маммальньх признаков (неполная заглазничная перегородка, высокий сагиттальный гребень на черепе, почти полностью сформированное вторичное костное небо, большая зубная кость), они стоят в стороне от линии, предковой для млекопитающих.
Первые тероцефалы были описаны в конце XIX ст. Р. Оуэном и Г. Сили. Группа была установлена Р. Брумом в 1903 г. В этом же году Р. Брум выделил из состава тероцефалов горгонопсов. В границах рассматриваемой здесь группы Л.П. Татаринов (2009 г.) разделяет тероцефалов и скалопозавров, указывая на существующие между ними различия в распределении небных зубов, развитии межптеригоидной ямы, а также на расширенную восходящую пластину эпиптеригоида у скалопозавров и неподвижное базиптеригоидное сочленение у тероцефалов.
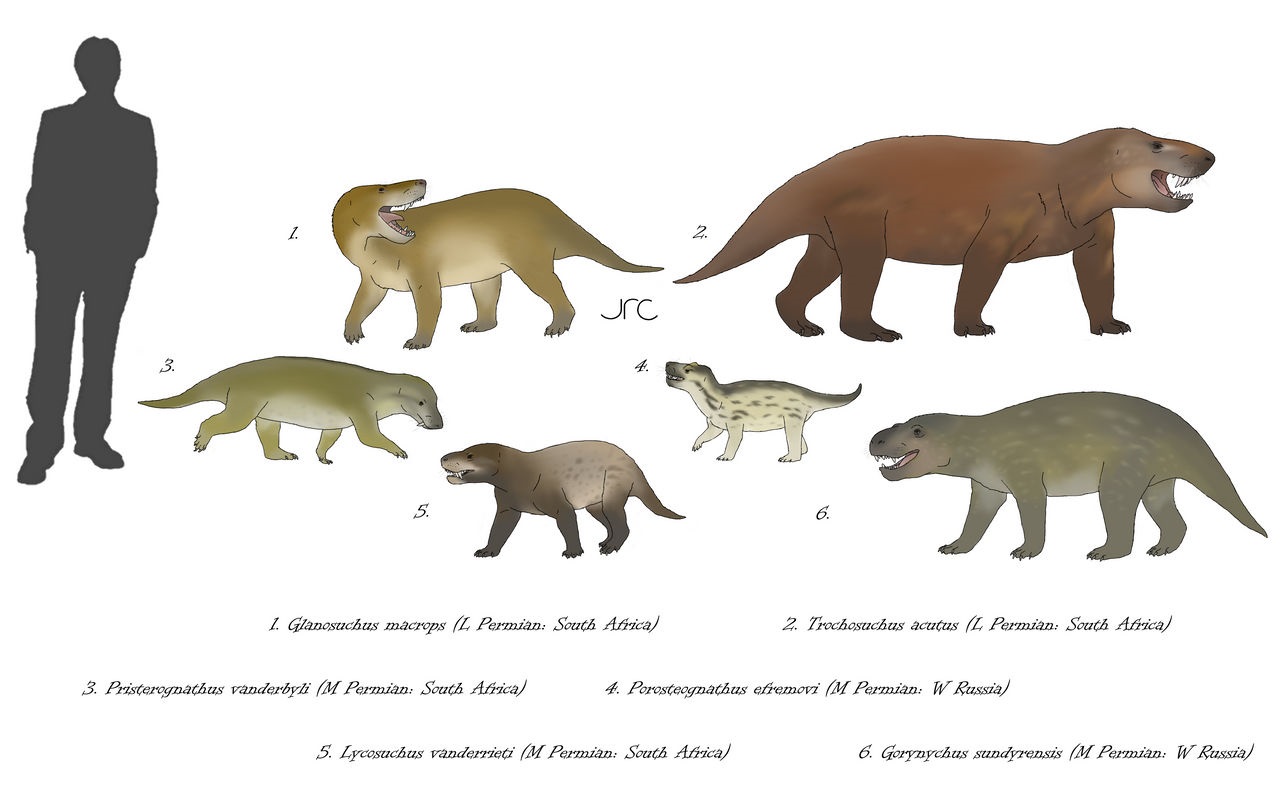
Разнообразие базальных тероцефалов. Автор — artbyjrc.
Дж. Хопсон и Г. Бархьюзен (1986 г.) объединили низших тероцефалов в группу Pristerognathosauria (Scylacosauridae, Alopecodontidae и Pristerognathidae), а высших тероцефалов — в группу Eutherocephalia (Lycosuchidae, Annatherapsididae, Whaitsiidae, Moschowhaitsiidae, Moschorhinidae, Euchambersiidae и Simorhinellidae).
А. Бринк (1986 г.) разделил скалопозавров на примитивных Lycideopsoidea (Lycideopsidae, Chthonosauridae) и типичных Ictidosuchoidea (Ictidosuchidae, Nanictidopsidae и Scaloposauridae). К последнему надсемейству Л.П. Татаринов (2009 г.) также относит Regisauridae и Bauriidae. Karenitidae и Scalopognathidae, по его мнению, можно сближать с другими семействами иктидозухоидей лишь условно, их целесообразно выделять в особые надсемейства.
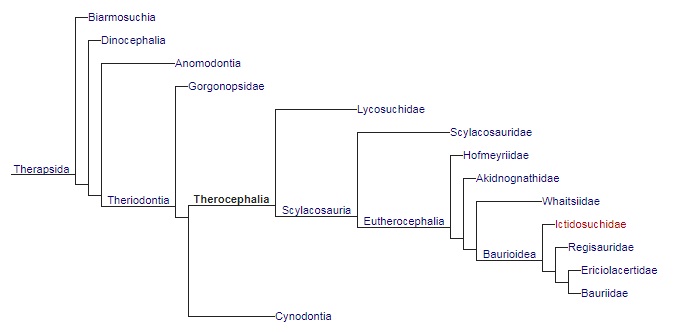
Кладограмма, иллюстрирующая родственные связи тероцефалов (по Сидору, 2001 и Гуттенлокеру, 2009).
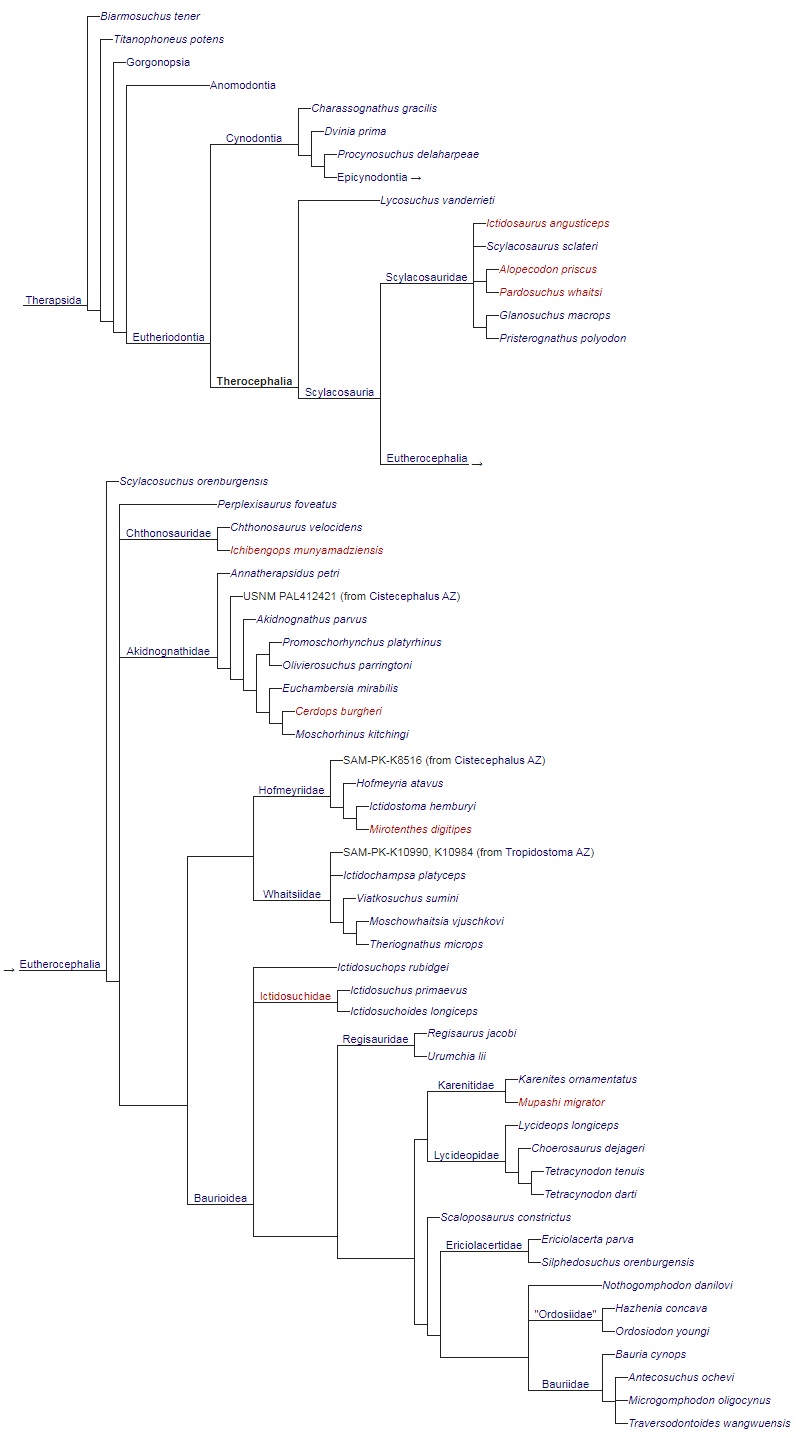
Кладограмма, иллюстрирующая родственные связи тероцефалов (по Адаму и Гуттенлокеру, 2014).
Ссылки
Тероцефалы (Википедия) перейти
Therocephalia (Paleobiology Database) перейти
Литература
Богданов Д. В. Звероящеры и другие пермские монстры. — М.: Фитон XXI, 2018
Иванов А. О., Черепанов Г. О. Ископаемые высшие позвоночные. — СПб.: Издательство Санкт-Петербургского университета, 2001
Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Ч. 1 / Отв. ред. Ивахненко М. Ф. и Курочкин Е. Н. — М.: ГЕОС, 2008
Кэрролл Р. Палеонтология и эволюция позвоночных. В 3 т. Т. 2 — М.: Мир, 1993
Основы палеонтологии. В 15 т. Т. 12: Земноводные, пресмыкающиеся, птицы / Гл. ред. Орлов Ю. А. — М.: Наука, 1964
Протеро Д. Отпечатки жизни. 25 шагов эволюции и вся история планеты. — СПб.: Питер, 2016
Ромер А. Ш. Палеонтология позвоночных. — М. — Л.: Государственное научно-техническое издательство нефтяной и горно-топливной литературы, 1939
Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М.: ГеОС, 2009