Тилакосмилы — Thylacosmilus
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Териеобразные (Theriiformes)
Клада Голотерии (Holotheria)
Клада Трехнотерии (Trechnotheria)
Клада Кладотерии (Cladotheria)
Клада Затерии (Zatheria)
Клада Трибосфениды (Tribosphenida)
Клада Терии (Theria)
Клада Метатерии (Metatheria)
Клада Сумчатообразные (Marsupialiformes)
Отряд †Спарассодонты (Sparassodonta)
Надсемейство †Боргиеноиды (Borhyaenoidea)
Клада †Тилакосмилообразные (Thylacosmiliformes)
Семейство †Тилакосмиловые (Thylacosmilidae)
Род †Тилакосмилы (Thylacosmilus)
Вид †Thylacosmilus atrox
Тилакосмилы (Thylacosmilus Riggs, 1933) — монотипический род семейства тилакосмиловых (Thylacosmilidae (Riggs, 1933)), происходящий из позднего миоцена - позднего плиоцена Южной Америки.

Реконструкция внешнего облика тилакосмила (Thylacosmilus atrox). Автор — Р. Евсеев.
Синонимы: Achlysictis Ameghino, 1891; Notosmilus Kraglievich, 1960.
Единственный установленный вид — Thylacosmilus atrox Riggs, 1933 (тип). Его остатки обнаружены на севере и востоке Аргентины (провинции Буэнос-Айрес, Катамарка, Кордова, Ла-Пампа и Энтре-Риос), а также в Уругвае. Они датируются поздним миоценом - поздним плиоценом (уайкерский - чападмалальский века). Синонимы: Achlysictis lelongi Ameghino, 1891; Thylacosmilus lentis Riggs, 1933; Notosmilus pattersoni Kraglievich, 1960; "Achlysictis atrox".

Неполные черепа с нижними челюстями тилакосмила (Thylacosmilus atrox). Слева: образец AMNH 32780, Американский музей естествознания, Нью-Йорк.
История изучения
Впервые ископаемые остатки тилакосмилов были обнаружены в 1926 г. в позднемиоценовых отложениях Арауканской формации в провинции Катамарка на севере Аргентины. Найденные остатки принадлежат 3 особям и представляют собой 2 неполных черепа, нижнюю челюсть, некоторые позвонки и кости конечностей. Эти первые находки одновременно являются и наиболее полными из когда-либо совершенных. Основываясь на них, Э. Риггс в 1933 г. описал 2 вида нового рода — типовой Thylacosmilus atrox и Thylacosmilus lentis, в дальнейшем синонимизированный с первым. В последующие годы был сделан ряд новых находок остатков тилакосмилов, как правило, весьма фрагментарных.

Неполные черепа тилакосмила (Thylacosmilus atrox).
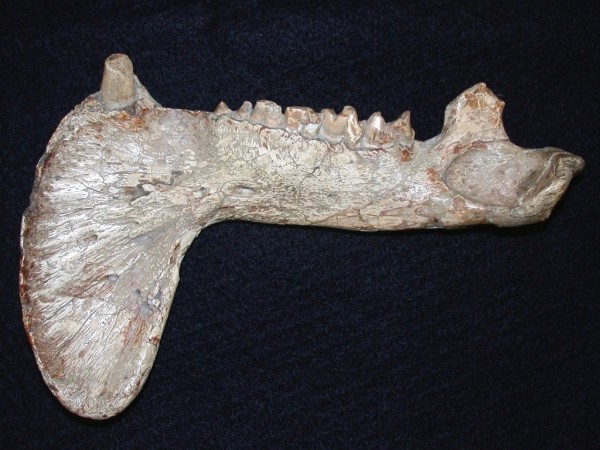
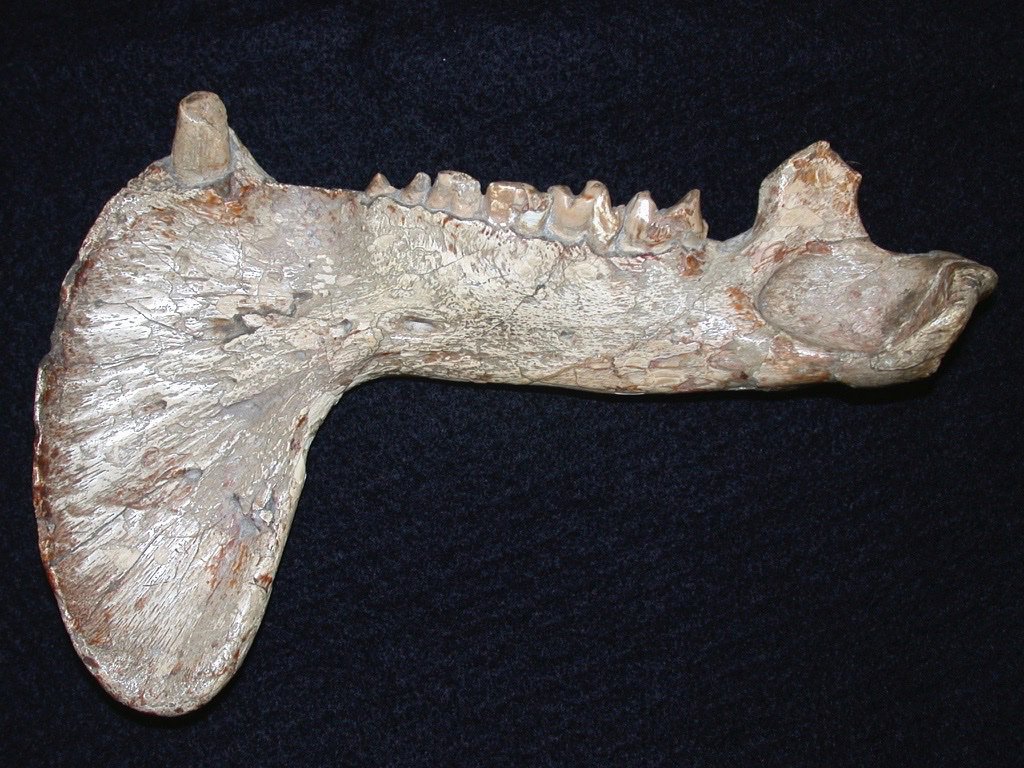
Ветвь нижней челюсти тилакосмила (Thylacosmilus atrox), снаружи.
Строение и функции
Размеры тилакосмила крупнейшие в его семействе и сопоставимы с таковыми современного ягуара (Panthera onca). Оценочная длина его тела без учета хвоста составляет 125-150 см, высота в холке — 55-70 см, вес — 60-120 кг. Таким образом, тилакосмил представлял собой довольно массивного, коренастого зверя. Несмотря на общее сходство с саблезубыми кошками (Machairodontinae), своим сложением он больше напоминал других хищных сумчатых, что особенно проявлялось в строении позвоночника, таза и лап.

Неполный череп с нижней челюстью тилакосмила (Thylacosmilus atrox).
Череп тилакосмила достигает 26 см в длину и характеризуется общей укороченностью. Его лицевой отдел очень массивный, а морда заметно сужается кпереди. Предчелюстные кости на найденных образцах не сохранились. Носовые кости длинные, узкие и сжатые с боков, они образуют небольшой клин и уходят под верхнечелюстные кости. Последние очень развиты, их верхние отростки проходят далеко позади глазниц и там расширяются; небо широкое. Глазницы небольшие и низко расположенные; в отличие от плацентарных хищников, они полностью замкнуты. Глаза были округлыми и сидели неглубоко. Имеется широкое подглазничное отверстие. Ушная капсула небольшая, наружный слуховой проход открывается латерально.
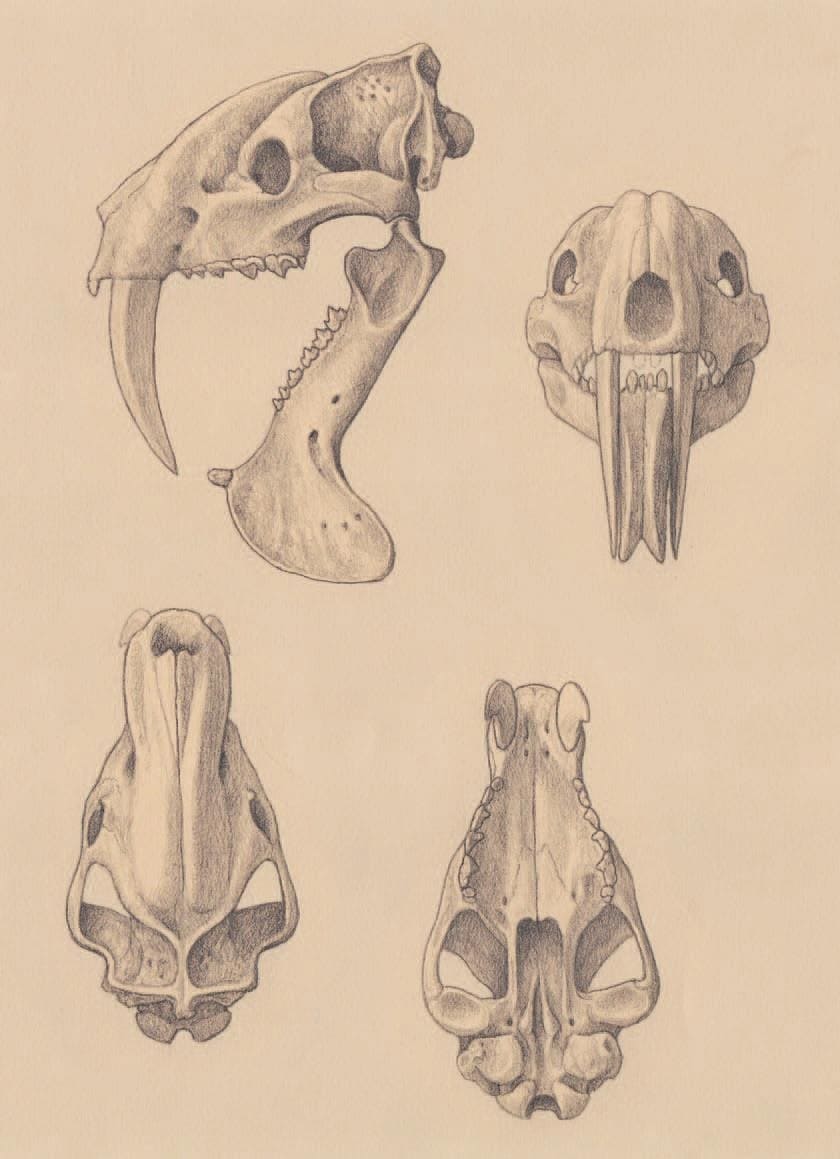
Прорисовки реконструированного черепа с нижней челюстью тилакосмила (Thylacosmilus atrox). Автор — М. Антон.
Вследствие разрастания верхнечелюстной кости лобная кость небольшая, наиболее широкая у верхнего края глазницы и далее суживающаяся, не участвующая у взрослой особи в образовании сагиттального гребня. Сагиттальный гребень короткий и высокий. Имелись мощные височные мышцы. Вся архитектура короткого и высокого черепа с круто спадающим лбом усиливает его к вертикальным нагрузкам при ударе клыками по кривой в направлении сверху вниз. Мозговой отдел черепа небольшой; головной мозг, как и у прочих сумчатых, был достаточно примитивным.
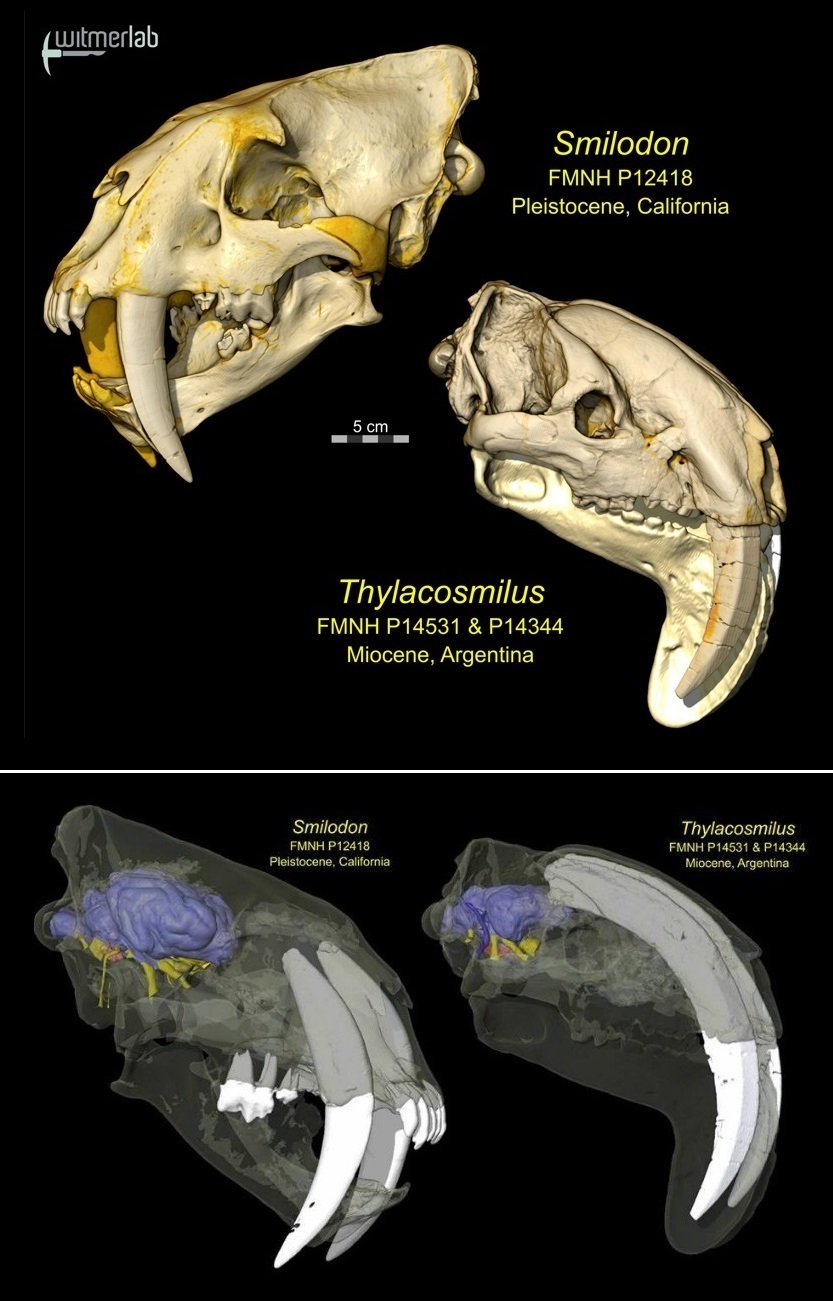
Трехмерные компьютерные модели черепа с нижней челюстью и головного мозга смилодона (Smilodon fatalis) и тилакосмила (Thylacosmilus atrox); масштаб не соблюден.
Затылочный гребень хорошо развит, что свидетельствует о наличии мощной шейной мускулатуры, крепившейся к затылку и обеспечивающей очень сильный удар клыками сверху вниз. Подобие в строении затылочного отдела черепа тилакосмила и саблезубых кошек позволяет предполагать, что группа мышц шеи, прикреплявшихся к черепу, у этих животных в анатомическом и функциональном отношениях была сходной.
Скуловые отростки небольшие, а нижнечелюстная мускулатура была довольно слабой и крепилась практически к самому концу челюсти. Ветвь нижней челюсти прямая и узкая, нижнечелюстной угол короткий. Все это, а также проведенный биомеханический анализ свидетельствует о том, что тилакосмил не обладал мощными укусом и хваткой. Сила его укуса составляла всего 80-100 кг, что уступает этому показателю даже у домашней кошки. Однако благодаря тому, что челюстной сустав сильно опущен вниз, этот хищник мог чрезвычайно широко открывать свою пасть, пуская в ход основное оружие убийства — саблевидные верхние клыки. При максимальном открывании пасти нижняя челюсть становилась к черепу под углом приблизительно в 100 °.
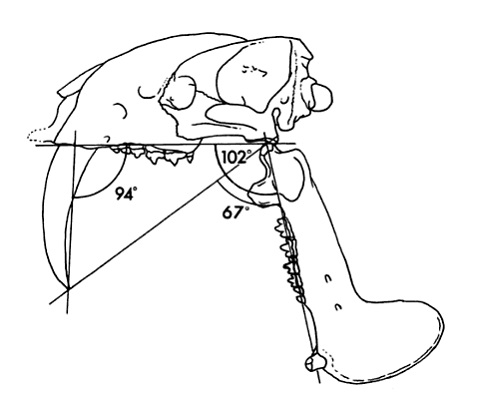
Схема черепа тилакосмила (Thylacosmilus atrox) с максимально отведенной нижней челюстью.
Зубная система тилакосмила демонстрирует целый ряд специфических особенностей. Его зубная формула — I1/i1, С1/с1, Р2/р2, М4/m4 = 32. Верхние резцы неизвестны, однако о их наличии можно судить по характерным следам износа на внутренних поверхностях нижних клыков. Размеры верхних резцов были совсем небольшими, что позволяло более эффективно использовать находящиеся за небольшими диастемами клыки — вонзать их в плоть жертвы максимально глубоко, практически на всю длину коронки. Нижние резцы развиты слабо и сильно сужены.
Верхние клыки очень мощные и длинные (длина их коронок около 15 см) — относительно длиннее, чем у всех других известных саблезубых хищников. Они располагаются параллельно, сплюснуты с боков и имеют трехгранную форму, что еще больше увеличивает их прочность при нагрузках. Боковые поверхности верхних клыков покрыты тонким слоем эмали. Очень длинные корни клыков, проходящие над глазницами и за ними, не замкнуты, что указывает на их постоянный рост. В отличие от саблезубых кошек, клыки находятся очень близко друг к другу. Несомненно, такие огромные клыки имели для выживания тилакосмила первостепенное значение, иначе они не увеличились бы до столь значительных размеров. Нижние клыки расположены вертикально, они мелкие, довольно слабые и обладают цилиндрическими корнями.
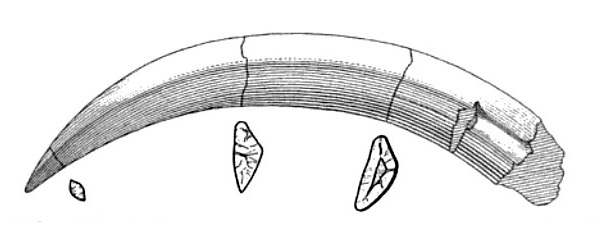
Прорисовки верхнего левого клыка тилакосмила (Thylacosmilus atrox), снаружи и в поперечном сечении.
Предкоренные зубы значительно редуцированы и уменьшены, Р1/р1 полностью отсутствуют. Между верхним клыком и предкоренными имеется небольшая диастема, очевидно возникшая на месте исчезнувшего предкоренного. Р2 конической формы, с 1 корнем; Р3 удлиненный, конический, с 2 корнями. На нижней челюсти между клыком и предкоренными лежит широкая диастема. Нижние предкоренные простой конической формы, имеют по 1 корню и несут выраженные следы износа.
Коренные зубы сравнительно небольшие. М1, М2 и М3 в большинстве случаев имеют сильно изношенные коронки. М3 продвинутого режущего типа, протокон и паракон на нем едва прослеживаются. М4 редуцирован до небольших размеров, его ширина больше длины; он обладает 1 корнем. Нижние коренные зубы сплюснуты с боков, с параконидом, протоконидом и талонидом, метаконид отсутствует. Как и верхние, они зачастую имеют сильно истертые коронки. m4 представляет собой хорошо развитый режущий зуб с уменьшенным талонидом.
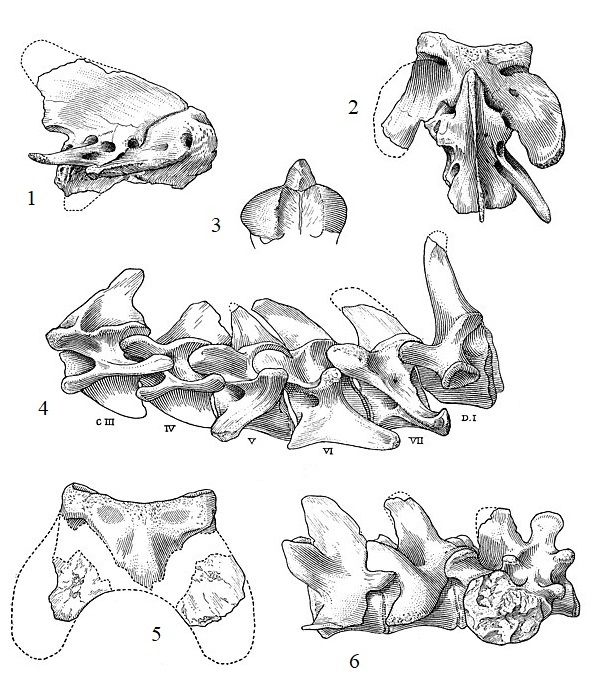
Прорисовки позвоночных элементов тилакосмила (Thylacosmilus atrox): 1, 2, 3 — эпистрофей сбоку, сверху и спереди; 4 — III-VII-е шейные и I-й грудной позвонки сбоку; 5 — атлант сверху; 6 — 2 последних поясничных и 2 крестцовых позвонка.
По обеим сторонам переднего конца нижней челюсти тилакосмила наличествуют характерные симфизные выступы — "лопасти", защищающие клыки при закрытой пасти. Они несколько загнуты назад и настолько велики, что даже при сомкнутых челюстях огромные верхние клыки как правило не достигают их нижнего края. Подобные отростки, выполнявшие ту же функцию, имеются у более древнего и примитивного представителя семейства — анахлизиктиса (Anachlysictis gracilis), а также у многих саблезубых кошек (Machairodontinae), барбурофеловых (Barbourofelidae), нимравовых (Nimravidae), части растительноядных (например, у диноцерат (Dinocerata)) и целого ряда хищных терапсид (Therapsida), однако ни у кого из них эти выступы не достигали столь же исключительных размеров относительно черепа, как у тилакосмила.
При жизни нижнечелюстные отростки были покрыты кожей и при закрывании рта плотно вдвигались между клыками. Существует предположение, что на данных выступах располагались не сохранившиеся в ископаемом состоянии специальные образования из мягких тканей, в которых верхние клыки прятались при закрывании пасти, словно в ножнах. Однако если учесть, что клыки могли расти на протяжении всей жизни животного, то тогда вполне возможно допустить наступление момента, когда они просто перерастали длину отростков и, соответственно, гипотетических ножен. Помимо этого, подобные образования наверняка служили бы потенциальным источником инфекции, так как в них непременно должны были скапливаться частички пищи хищника. Таким образом, "футляров" для клыков скорее всего не было.
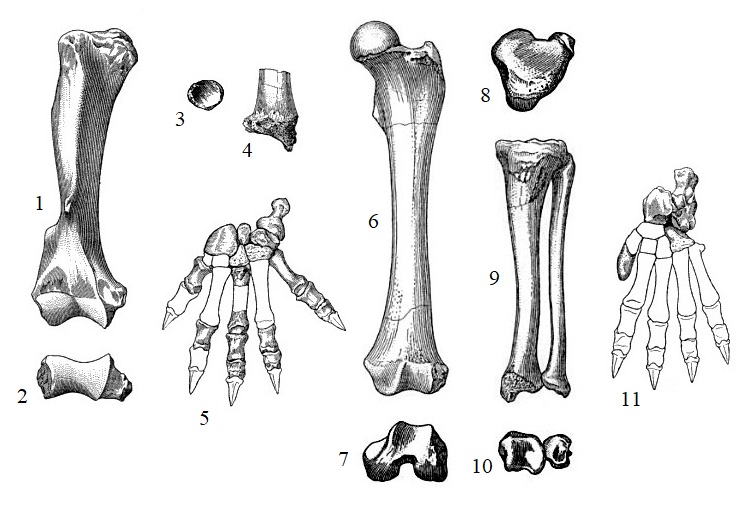
Прорисовки элементов конечностей тилакосмила (Thylacosmilus atrox): 1,2 — правое плечо спереди и дистально; 3, 4 — правая лучевая кость проксимально и дистально; 5 — левая кисть сверху; 6, 7 — левое бедро спереди и снизу; 8 — большая берцовая кость сверху; 9, 10 — большая и малая берцовые кости спереди и снизу; 11 — левая стопа сверху.
Шейные позвонки обладают выдающимся нижним килем и хорошо развитыми верхним и боковыми отростками для прикрепления сильных мышц, опускающих голову вниз и поворачивающих ее в стороны. Кроме того, шея тилакосмила, как и у многих других саблезубых хищников, была довольно длинной. Длинная, а не просто мускулистая шея была необходима этому животному для лучшего замаха, что обеспечивало большую скорость, а значит и силу удара клыками. Не менее важное значение при нанесении укуса имел быстрый и точный поворот головы. Кроме того, мощная шейная мускулатура компенсировала относительную слабость их челюстных мышц, что выражается у тилакосмила особенно ярко.
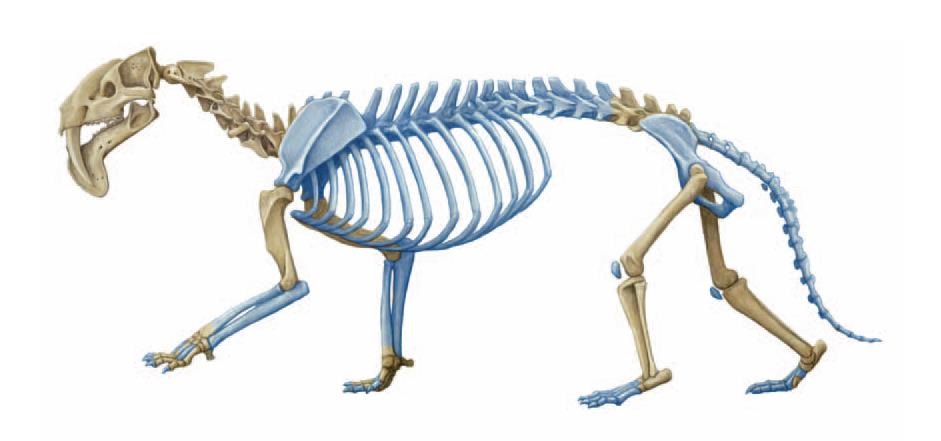
Реконструкция скелета тилакосмила (Thylacosmilus atrox). Синим цветом показаны отсутствующие кости, достроенные на основании изучения остатков других боргиеноидов. Автор — М. Антон.
Туловище тилакосмила было удлиненным и гибким, плечевой пояс мощным, а конечности — довольно короткими и сильными. Его лучевая кость массивная, короче плечевой. Локтевая и карпальные кости свободные. Кисть широкая, с 5 широко расставленными пальцами. Бедренная кость длиннее большеберцовой, малоберцовая кость свободная. Стопа широкая, пятипалая. I-й палец состоит из единственной редуцированной фаланги. Передние конечности полупальцеходящие, задние — стопоходящие. По всей вероятности, когти были массивными, короткими, довольно острыми, но не способными втягиваться. Крестец состоит из 2 сросшихся позвонков. Хвост предположительно являлся достаточно длинным, толстым и жестким.

Реконструированные скелеты тилакосмила (Thylacosmilus atrox).
Окраска шерсти тилакосмила с определенной степенью достоверности реконструируется благодаря изучению ныне живущих сумчатых. Если этот зверь был лесным обитателем, весьма вероятным для него являлся темный фон со светлыми пятнами — это стандартная для сумчатых схема окраса. Если животное населяло более открытые пространства, оно могло быть однотонным.
Образ жизни
Анатомическое строение тилакосмила свидетельствует о том, что он специализировался в охоте на крупных, хорошо защищенных, но при этом медлительных млекопитающих из засады или путем скрадывания. Несомненно, этот хищник практически не был приспособлен к быстрому бегу. Наиболее вероятной добычей тилакосмила являются токсодонтовые (Toxodontidae) и различные наземные ленивцы — например, мегатериевые (Megatheriidae). Что касается других южноамериканских копытных — прочих нотоунгулят (Notoungulata), а также литоптерн (Litopterna), то они в большинстве своем отличались исключительной верткостью и маневренностью и поэтому вряд ли часто становились добычей этого сравнительно медлительного хищника. Все же нельзя полностью исключать возможность того, что он мог нападать на подобных быстроногих животных, охотясь на них из засады и полагаясь на точность броска.

Тилакосмил (Thylacosmilus atrox) в естественном окружении. Автор — Р. Евсеев.
Потенциальные жертвы тилакосмила обычно имели толстую или даже бронированную шкуру и порой в десятки раз превосходили его по своему весу. Во избежание повреждения тонких клыков, укус этим хищником наносился преимущественно в область брюха, или сбоку в шею у относительно некрупных животных. Морфологический анализ лап и плечевого пояса, а также изучение способов охоты ряда современных сумчатых и плацентарных хищников позволяют утверждать, что тилакосмил активно использовал передние конечности при нападении на добычу для ее удержания и обездвиживания наподобие того, как это сейчас делают большие кошки. Рана от огромных и близко расположенных верхних клыков, нанесенная в наиболее уязвимую часть тела жертвы, вызывала у последней обильное кровотечение. Когда добыча ослабевала настолько, что уже не могла двигаться, хищник приступал к процессу питания.
Вопрос о том, каким образом тилакосмил разделывал туши убитых им животных, натыкается на некоторые трудности. Дело в том, что зубочелюстной аппарат тилакосмила, столь высоко специализированный к убийству крупной добычи, на первый взгляд кажется практически неприспособленным к дальнейшей ее утилизации. Почти полностью отсутствующие резцы, громадные верхние клыки (достаточно острые только на своих кончиках), небольшие щечные зубы, слабые челюстные мышцы — все это совсем не способствовало разрыванию плотных внешних покровов, разрезанию сухожилий и отделению кусков мяса поверженной добычи.

Этапы портретной реконструкции тилакосмила (Thylacosmilus atrox). Автор — Р. Евсеев.
Высказывались предположения, что тилакосмил питался преимущественно внутренними органами и даже кровью своих жертв, однако выглядят они неправдоподобно. Питание кровью не просто потребовало бы коренной перестройки зубочелюстного аппарата по сравнению с тем, что мы наблюдаем, оно попросту невозможно для столь крупного животного. Питание исключительно внутренностями выглядит нерационально с экологической точки зрения — маловероятно, что такой крупный хищник довольствовался настолько малой долей добычи и бросал нетронутой основную ее часть. Кроме того, длинные верхние клыки должны были сильно мешать при попытке извлечь из грудной клетки сердце и легкие.
Таким образом, остается предполагать, что тилакосмил потреблял более или менее существенную часть туши, включая значительную часть мяса и внутренние органы. Но чтобы добраться до них, хищнику сначала необходимо было вскрыть плотные кожные покровы добытого им животного. Производилось это именно зубами, так как помимо них, ничем другим зверь действовать не мог. Вероятно, тилакосмил поедал свою жертву, начиная с наиболее мягкого паха. Очевидно, ткани разрезались, откусывались и разжевывались щечными зубами, как это происходит у плацентарных хищников. Именно по причине активного использования щечных зубов они и доходят до нас преимущественно в сильно изношенном состоянии. Что же касается верхних клыков, то рвать ими добычу тилакосмил не мог — для этого они были слишком длинными и тонкими, и попросту не выдержали бы сильной боковой нагрузки. При этом с прямыми вытягивающими движениями при извлечении внутренностей клыки справлялись неплохо.

Портретные реконструкции тилакосмила (Thylacosmilus atrox). Автор — В. Симеоновски.
Редукция резцов говорит о том, что тилакосмил не был способен эффективно соскребывать мясо с костей и поэтому оставлял падальщикам довольно значительную часть добычи. Не исключено, впрочем, что язык тилакосмила являлся достаточно длинным и был покрыт специальными зубчиками, выраженными сильнее, чем у современных крупных кошек. Такой язык действовал подобно терке и при активном лизании отделял маленькие кусочки плоти. Кроме того, при разделывании туши тилакосмил наверняка помогал себе сильными передними лапами с острыми невтяжными когтями.
Так как тилакосмил съедал добычу не полностью, охотиться он вынужден был довольно часто. Вероятно, при возможности им охотно поедались и подвернувшиеся трупы крупных животных, однако рассчитывать на них особо не приходилось — падаль в первую очередь съедалась более мелкими хищниками, численность которых была явно выше и которые поэтому находили трупы быстрее.

Портретная реконструкция тилакосмила (Thylacosmilus atrox). Автор — М. Антон.
Своеобразная морфология зубной системы тилакосмила служит косвенным подтверждением сравнительно небольшой продолжительности его жизни. Сумчатые, как правило, вообще живут недолго, но в случае тилакосмила имеются также дополнительное свидетельство — очень значительная истертость большинства дошедших до нас щечных зубов. Так как они постепенно изнашивались (не восстанавливаясь, в отличие от клыков) практически при каждом приеме пищи, то за сравнительно небольшое количество лет достигали настолько плачевного состояния, что зверь уже не мог эффективно ими пользоваться и попросту умирал от голода.
О социальности тилакосмила достоверно ничего неизвестно, но об этом вопросе с определенной степенью уверенности можно судить благодаря исследованиям поведения современных сумчатых хищников. Очевидно, тилакосмил жил в одиночку. Каждая взрослая особь занимала свой охотничий участок и только в период размножения самка и самец могли некоторое время держаться рядом. В дальнейшем самка растила единственного или нескольких своих отпрысков. Наличие выводковой сумки у тилакосмила не установлено (она и сейчас имеется не у всех сумчатых). Если сумка была, то, вероятнее всего, открывалась назад, что обеспечивало лучшую защиту молодняка во время охоты матери. Не исключено также, что самка учила охотиться своих подросших детенышей.

Пара отдыхающих тилакосмилов (Thylacosmilus atrox). Автор — В. Симеоновски.
Тилакосмил жил рядом с очень крупными нелетающими хищными птицами семейства форораковых (Phorusrhacidae). Как и тилакосмил, форораки часто охотились на крупных млекопитающих, что могло порождать между этими хищниками конкуренцию. К тому же, форораки предположительно являлись стайными животными, тогда как тилакосмил вел одиночный или, в крайнем случае, парный (семейный) образ жизни. Однако эти птицы скорее всего населяли более-менее открытые ландшафты, в то время как пропорции тилакосмила указывают на то, что он отдавал предпочтение залесенным участкам. При беге форораки могли развивать значительную скорость и, судя по всему, были очень выносливыми. Вероятно, и тилакосмил в свою очередь являлся довольно выносливым животным (что в целом характерно для сумчатых), однако далеко не таким быстрым, как форораки. Очевидно, эти пернатые хищники в большей или меньшей мере создавали определенное давление на популяцию тилакосмила. Так или иначе, он является единственным действительно крупным аборигенным хищником Южной Америки, который на протяжении достаточно длительного времени смог сосуществовать с форораками.

Гигантский тераторн аргентавис (Argentavis magnificens) пытается отнять у тилакосмила (Thylacosmilus atrox) его добычу — броненосца вассаллию (Vassallia maxim). Автор — В. Симеоновски.
Вымирание
Причины исчезновения тилакосмила не выяснены. Одним из наиболее распространенных предположений, объясняющим его вымирание, является миграция крупных представителей семейства кошачьих (Felidae) из Северной Америки в Южную после образования Панамского перешейка. На первый взгляд, эта версия кажется весьма логичной, так как, являясь плацентарными, кошки находились на более высоком уровне физической организации, обладали лучше развитым интеллектом, а некоторые из них могли вести коллективный образ жизни. Однако у этой гипотезы имеется существенный недостаток. Дело в том, что, согласно имеющимся данным, тилакосмил вымирает порядка 3,5-3 млн. лет назад, т. е. задолго до прихода в Южную Америку больших кошек, которые появляются здесь не менее чем 1 млн. лет спустя.

Черепа с нижними челюстями тилакосмила (Thylacosmilus atrox) и смилодона (Smilodon fatalis), приведенные в общем масштабе.
Можно также предположить, что тилакосмила постепенно вытеснили форораки, но в таком случае остается непонятным, как он вообще мог эволюционировать, ведь этот сумчатый хищник появляется в эпоху, когда семейство форораковых как раз находилось в полном расцвете. Вероятно, исчезновение тилакосмила было сопряжено с целой совокупностью факторов. Например, к этому могла привести смена состава фауны растительноядных млекопитающих, происходившая на фоне постоянного натиска форораков в период глобального иссушения климата.
Ссылки
Сумчатый саблезубый тигр (Википедия) перейти
Сумчатый саблезубый тигр может лишиться звания хищника. — 2020 (ECOportal) перейти
Thylacosmilus (Paleobiology Database) перейти
Литература
Быстров А.П. Находка саблезубого сумчатого хищника. Природа 12, 1934
Кэрролл Р. Палеонтология и эволюция позвоночных. В 3 т. Т. 3 — М.: Мир, 1993
Симпсон Дж. Великолепная изоляция. История млекопитающих Южной Америки. — М.: Мир, 1983
Тернер А., Антон М. Большая энциклопедия доисторических животных. — М.: Оникс, 2006
Шпинар З., Буриан З. История жизни на Земле. — Прага: Артия, 1977
Croft D. A. Horned armadillos and rafting monkeys: the fascinating fossil mammals of South America. — Bloomington: Indiana University Press, 2016
Marshall L. G. Evolution of the Thylacosmilidae, extinct sabertooth marsupials of South America. PaleoBios 23, 1976
Riggs E. S. Preliminary description of a new marsupial sabertooth from the Pliocene of Argentina. Geological Series of Field Museum of Natural History 6, 1933
Riggs E. S. A new marsupial saber-tooth from the Pliocene of Argentina and its relationships to other South American predacious marsupials. Transactions of the American Philosophical Society 24 (1), 1934