Состав
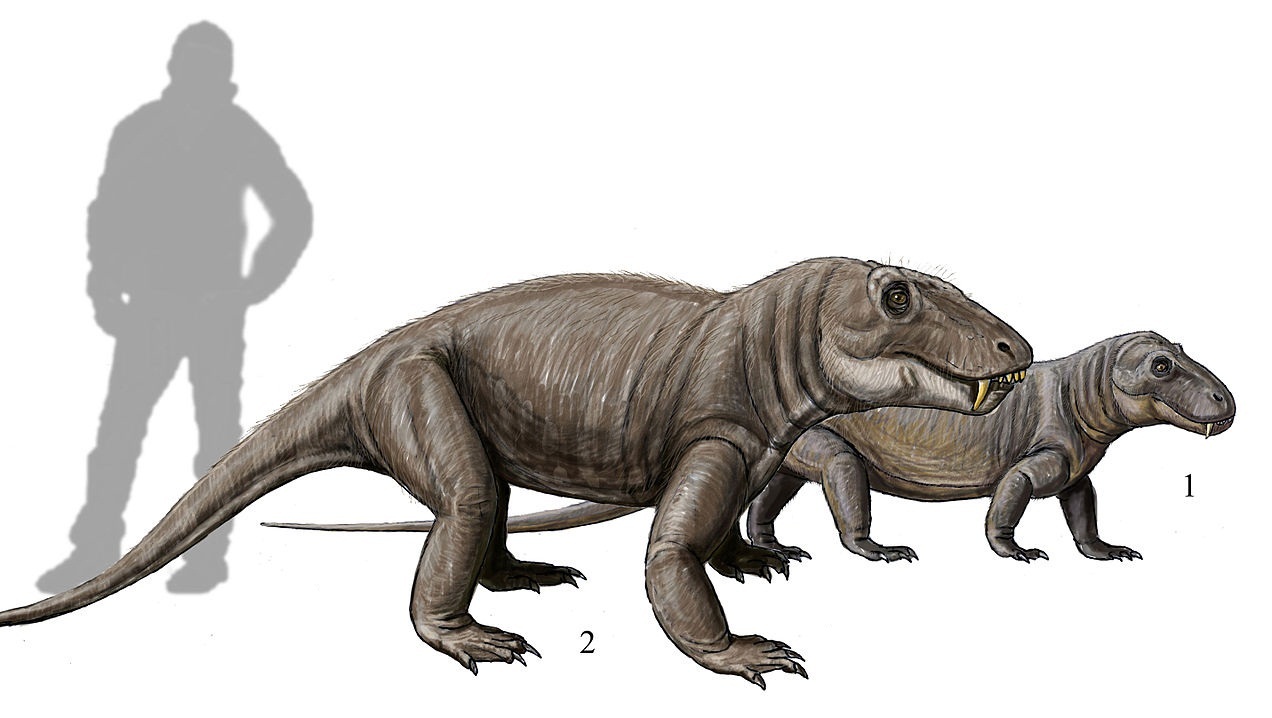
Реконструкция внешнего облика молодого (1) и взрослого (2) титанофонеев (Titanophoneus potens). Автор — Д. Богданов.
Titanophoneus potens Efremov, 1938 (тип) — типовой вид, известный из средней перми (зона Ulemosaurus, уржумский горизонт, уржумский ярус, биармийский отдел) местонахождения Ишеево (Апастовский район, Республика Татарстан, Россия). Синонимы: Doliosaurus yanshinovi Orlov, 1958; Doliosauriscus yanshinovi Kuhn, 1961. Это один из наиболее хорошо изученных диноцефалов.



Скелет молодой особи титанофонея (Titanophoneus potens) из местонахождения Ишеево, Республика Татарстан, Россия (расположен в позе обнаружения).
При первоописании вида И.А. Ефремов (1938 г.) в качестве голотипа указал 2 находки — почти полный скелет длиной около 3 м и разломанный череп длиной 53 см, принадлежавший гораздо более крупной особи ("большому старому самцу", по словам автора). При этом была специально подчеркнута их принадлежность одной форме. Ю.А. Орлов (1958 г.) принял указанный череп как голотип нового рода и вида диноцефалов — Doliosaurus yanschinovi. Поскольку вскоре выяснилось, что это родовое наименование преоккупировано и является синонимом ящерицы Phrynosoma, в 1961 г. оно было изменено на Doliosauriscus. М.Ф. Ивахненко (2008 г.), сравнивая диагнозы обеих форм, отмечает, что основные признаки, отличающие их, связаны с развитием у долиозавриска возрастного пахиостоза в лобно-теменной области. Имеются фрагменты черепных костей, по величине и степени развития пахиостоза промежуточные между типовыми экземплярами обеих форм. Таким образом, в данном случае мы имеем дело с остатками молодых и старых особей. Примечательно, что в Ишеево найдены обломки еще более крупных черепов того же диноцефала, длина которых оценивается в 80 см.
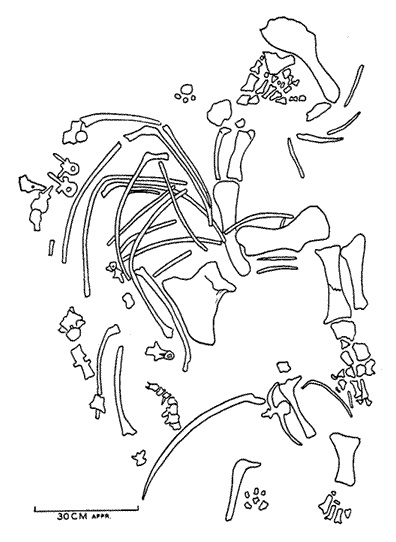
Прорисовка частичного скелета титанофонея (Titanophoneus potens) из местонахождения Ишеево, Республика Татарстан, Россия.
Некоторые отличия в абрисе черепов сбоку в старых работах связаны с тем, что типовой череп титанофонея разломан и передняя часть его резко вздернута вверх, а затем разлом загипсован и закрашен. На самом череп не являлся настолько бульдогообразным. Наличие у долиозавриска углублений на небе для заклыковых зубов нижней челюсти также может быть обусловлено сильным пахиостозом в этой области. С другой стороны, приводившееся раньше указание на отсутствие на небе ямы для нижнечелюстного клыка у титанофонея ошибочно. Некоторое расхождение в стороны задних краев дорсальных отростков предчелюстных костей на черепе долиозавриска связано с сильной вертикальной деформацией образца. Таким образом, по мнению М.Ф. Ивахненко, Doliosauriscus отходит в младшие синонимы Titanophoneus.

Частичный скелет взрослой особи титанофонея (Titanophoneus potens), или "долиозавриска" (Doliosauriscus yanshinovi), из местонахождения Ишеево, Республика Татарстан, Россия.
Череп сравнительно длинный, заглазничная длина укладывается в его общей длине 5 раз. С возрастом череп становится относительно выше и шире, особенно в скуловой области. Заостренные концы дорсальных отростков предчелюстных костей разобщены и вклиниваются в носовые. Глазницы средней величины, округлые. Они направленные косо в стороны и довольно сильно вперед, что важно при высматривании добычи. Глазное яблоко снабжено кольцом плоских, плотно сочлененных друг с другом косточек, увеличивавших устойчивость склеры. Глазничный край скуловой кости у молодых особей тонкий, с возрастом утолщается, равно как и заглазничная дуга. Височная дуга высокая и тонкая у молодых особей, низкая и толстая, с очень массивным заднем краем у взрослых. Височное окно крупное, открытое сверху; его край глубоко вклинен в отросток скуловой кости. Контакт скуловой и чешуйчатой костей с возрастом уменьшается. Надвисочная кость сильно редуцирована или отсутствует. У молодых особей предлобная кость и верхний край глазницы имеют слабобугристую поверхность, у взрослых задняя половина носовых костей, предлобные, лобные, заднелобные кости сильно утолщены и несут мощные бугристые разращения, испрещенные бороздами кровеносных сосудов.



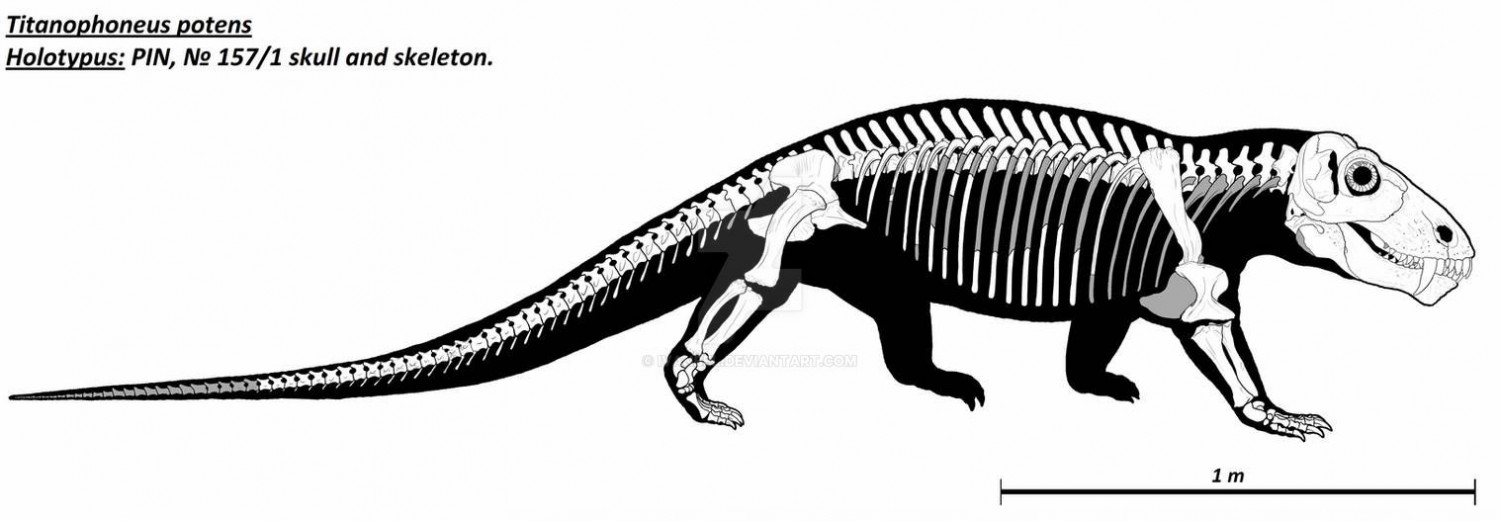
Реконструированный скелет взрослой особи титанофонея (Titanophoneus potens), или "долиозавриска" (Doliosauriscus yanshinovi), из местонахождения Ишеево, Республика Татарстан, Россия (смонтирован в нехарактерном для животного положении).
Межглазничный и теменной отделы сужены. Межтеменная кость очень высокая, сильно расширенная внизу, с мощным сагиттальным гребнем. Теменное отверстие расположено в задней части теменных костей и окружено высоким костным воротничком. Отверстие слабо удлиненно-овальное, относительно крупное у молодых особей и небольшое у взрослых. Затылок вертикальный, высокий и расширенный. Верхняя затылочная кость низкая и широкая. Небольшой затылочный мыщелок отогнут вниз — очевидно, голова обычно была наклонена мордой довольно сильно книзу. Вся затылочная область глубоко вогнутая, что говорит о мощной шейной мускулатуре. Большой срединный вертикальный затылочный гребень указывает на сильную срединную затылочную связку, что отчасти связано с тяжелым черепом (своей упругостью она облегчает приведение головы из опущенного положения в нормальное). Несмотря на большое височное окно и значительное развитие гребней на окружающих его костях, например на теменных, височная мускулатура была довольно слабой. Медиальный мыщелок квадратной кости несет отчетливый косой желоб.

Череп с нижней челюстью молодой особи титанофонея (Titanophoneus potens) из местонахождения Ишеево, Республика Татарстан, Россия.
Небо сводчатое. Хоанная область глубокая, разделенная парными, вогнутыми сошниками. Хоаны несколько смещены назад, так что их передние края располагаются на уровне клыков. Гребень на небе, образованный небной и крыловидной костями, выражен слабо. Каждая небная кость снабжена небольшим, четко обособленным низким бугром, поперечный диаметр которого равен 1/3 длины поперечной ветви крыловидной кости. Из небного бугра вниз торчат небольшие цилиндрические зубы с тупоконической вершиной — от 7-8 у молодых особей до 10-13 у взрослых. Массивные поперечные ветви крыловидных костей, примыкающих сзади к небным, внешними утолщенными концами опускаются далеко вниз. На их нижней стороне также имеются зубы, сходные с небными, но посаженные почти горизонтально, вершинами в стороны ветвей нижней челюсти. Крыловидная кость молодых особей несет лишь 2 сравнительно крупных зуба, тогда как у взрослых особей имеется 5-6 зубов: 3 мелких на поперечной ветви и 2-3 небольших на медиальном бугре. Озубленные зубы бугры на крыловидных костях не развиты.
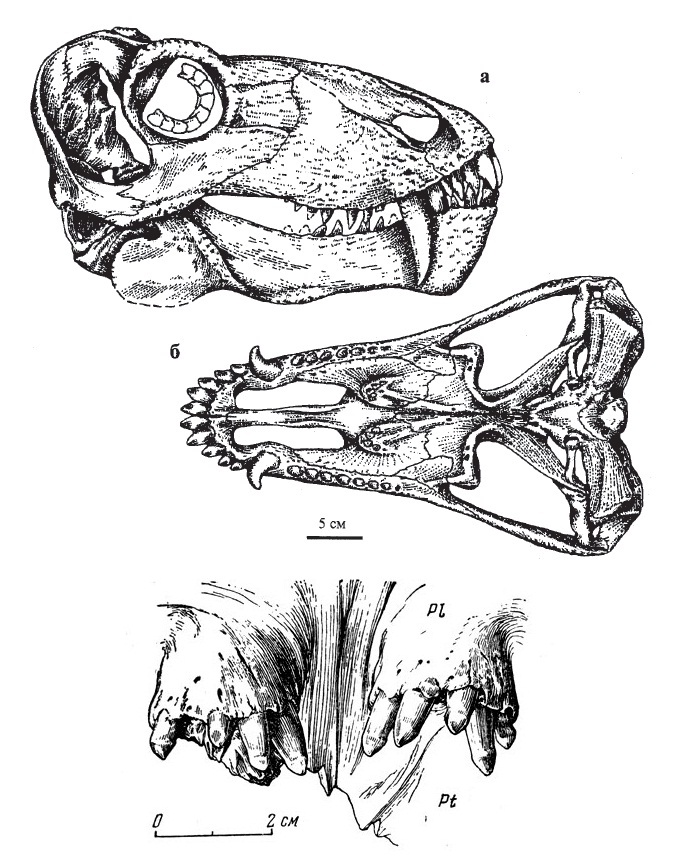
Прорисовки реконструированного черепа молодой особи титанофонея (Titanophoneus potens) из местонахождения Ишеево, Республика Татарстан, Россия; сбоку и сверху, а также прорисовка его небных бугров с зубами снизу и спереди.
На дорсальной стороне крыловидного комплекса позади поперечных ветвей имеется дополнительное окостенение в виде высоких и широких парасагиттальных выростов. Межптеригоидная щель очень узкая. На небной поверхности также присутствуют особые впадины для вхождения вершин нижних клыков (впереди верхних), а у взрослых особей — и для нескольких нижних заклыковых зубов. Высказывалось предположение, что наличие небных ямок для нижних зубов может указывать на сильное ороговение эпителия неба и ротовой полости в целом. Челюстное сочленение слабо опущено и лишь слегка смещено вперед по отношению к затылочному. Нижняя челюсть высокая и массивная, расширенная впереди клыков. Симфиз высокий и приподнят кверху, в соответствии с изгибом края верхней челюсти. Пахиостозный бугор на угловой кости низкий, изометрично-округлый.
Столбик уха, или стремя, представляет собою небольшую распорку в основании черепа между квадратной костью и мозговой коробкой, где он широким основанием закрывает овальное окно. Это положение столбика как будто исключает его подвижность и, на первый взгляд, приспособленность для передачи звуковых колебаний через овальное окно во внутреннее ухо, хотя по своему местоположению он и равнозначен стремечку среднего уха млекопитающих, птиц и пресмыкающихся, имеющих барабанную перепонку. Как воспринимали звук титанофонеи и другие синапсиды с подобным строением ушной области — не совсем ясно, но вряд ли они были глухими.

Нижняя челюсть молодой особи титанофонея (Titanophoneus potens) из местонахождения Ишеево, Республика Татарстан, Россия.
Зубная формула: I5/i4, C1/c1, Pc 6-8/pc6-9. Резцы крупные, остроконические, несколько скошенные вперед (особенно верхние), с режущими кантами и слабо выраженными пяточными выступами, в которые упираются вершины зубов противоположной челюсти. Клыки длинные, слабо изогнутые назад, резко сжатые с боков у молодых особей и почти округлые в сечении у взрослых, с килями по переднему и заднему краям и мощными, глубоко сидящими корнями. Задний край клыка снабжен мелко зазубренным режущим гребнем, отходящим от вершины зуба к его основанию. Заклыковые зубы не образующие сомкнутого ряда, однородные, низкие, слегка наклоненные назад, конические, каплевидные в сечении, уплощенные с лингвальной стороны, с зазубренными режущими кантами. Отмечена тенденция к уменьшению количества заклыковых зубов с возрастом.
Мощные резцы и особенно клыки приспособлены в основном для хватания добычи и ее разрывания на части. Гребни на краях верхних резцов, заходящие за края нижних, делают резцы одновременно режущими, а наличие на резцах зачаточных пяток указывает на их слабо выраженную измельчающую функцию (гораздо более совершенны в данном отношении резцы тапиноцефалоидов). Сравнительно слабые заклыковые зубы, хотя и режущие, но редкие и малоудобные для измельчения мышц, связок и грубой кожи, скорее всего, играли вспомогательную роль при удержании добычи. Небные зубы торчат вершиной вниз из бугров небных костей и при отсутствии зубов, противопоставленным им внизу, не могли использоваться для жевания. Вероятно, они представляют собой своеобразное раздавливающее, а частично и удерживающее добычу приспособление. Вполне вероятно, что для раздавливания отчасти служили и массивные поперечные отростки крыловидных костей.

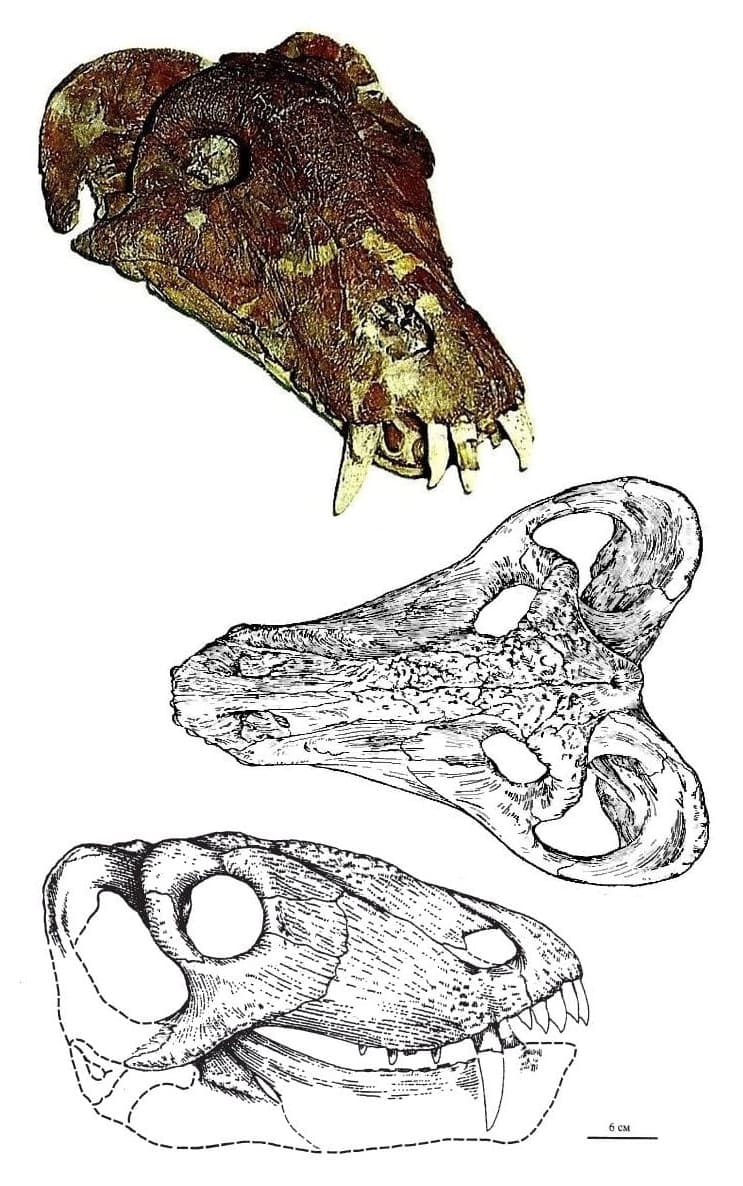
Череп взрослой особи титанофонея (Titanophoneus potens), или "долиозавриска" (Doliosauriscus yanshinovi), из местонахождения Ишеево, Республика Татарстан, Россия, а также его прорисовки сверху и сбоку.
При захлопывании пасти верхние резцы очень плотно вклиниваются в промежутки между нижними (как и нижние между верхними), а вершины нижних клыков и заклыковых зубов довольно точно входят в соответствующие им ямки на небе. Объединенность ямок в одном общем желобообразном углублении может указывать на возможность небольших передне-задних движений нижней челюсти; об этом же говорит и несколько уплощенная форма челюстного сустава. Нижние заклыковые зубы при этом ложились внутрь от верхних, не соприкасаясь с ними; это дополнительно свидетельствует о неприспособленности заклыковых зубов к измельчению пищи.
Точность в смыкании челюстей, без которой животное при стремительном хватании добычи могло бы сломать себе зубы, обеспечивалось строением челюстного сустава — глубокого двойного блока вверху и двойной впадины в нижней челюсти. Очевидно, с этим же связано и сильное развитие поперечных отростков крыловидных костей, вероятно, с упором их изнутри в нижнюю челюсть при закрывании рта. Боковые движения нижней челюсти в данном случае совершенно исключены. Судя по всему, все это помогало и удержанию добычи. Приводящие нижнюю челюсть мышцы мало заходят в заглазничную область, что свидетельствует о относительно слабом укусе. Это довольно успешно компенсировалось увеличенными режущими клыками и мощной шейной мускулатурой.

Портретные реконструкции молодого (слева) и взрослого (справа) титанофонеев (Titanophoneus potens). Автор — Р. Евсеев.
Позвоночный столб по сравнению с черепом относительно стройный и легкий. Позвонков около 100, они двояковогнуты и обладают тонким отверстием для хорды. Тела позвонков на переднем и заднем концах — с округлыми внешними краями, что указывает на гибкость позвоночника. В коротком и мощном шейном отделе 7-8 позвонков. Эпистрофей снабжен массивным остистым отростком. Это, вероятно, связано с большим весом головы, что требовало сильной затылочной связки и мускулатуры. В общей сложности, насчитывается не менее 33 предкрестцовых позвонков. 3 крестцовых позвонка неподвижно соединены с тазом крестцовыми ребрами. Хвост длинный, в нем порядка 60 позвонков. Между хвостовыми позвонками сохраняются свободные интерцентры.
Ребра двухголовчатые. Лопатка, ключица и межключица узкие. Лопатка изогнута очень слабо. Поскольку ее кривизна при близком расположении к грудной клетке в известной степени отражает форму самой клетки, вполне возможно приписать титанофонею и сжатое с боков туловище, а не уплощенное сверху вниз, как, например, у крокодилов. Вентральная часть тазового пояса расширенная. В симфизе таза между лобковыми и седалищными костями имеется зона неполного окостенения. В вертлужной впадине между подвздошной, лобковой и седалищной костями окостенение неполное. Подвздошная кость по строению промежуточна между таковой у пеликозавров, где она вытянута назад, и подвздошной более поздних терапсид, у которых верхний отдел кости вытянут вперед, как у млекопитающих.
Прорисовка реконструированного скелета молодой особи титанофонея (Titanophoneus potens), вписанного в телесные очертания. Автор — И. Белоусов.
Конечности довольно длинные и стройные, передние несколько короче задних. Плечо с предплечьем и бедро с голенью конвергентно схожи с крокодильими, что свидетельствует о расположении конечностей по бокам туловища и невозможности постоянно поддерживать туловище над землей. Слабое окостенение эпифизарных районов свидетельствует о подвижности конечностей в суставах. Плечевая кость с обоими эпикондилярными отверстиями. Бедренная кость удлиненная и изогнутая.
Кисть и стопа очень широкие, главным образом за счет пястных и плюсневых костей, увеличивающихся от I-го пальца к V-му. Большая величина лап особенно заметна, если сравнить титанофонея с крокодилом, даже вдвое или втрое превосходящего его по размеру. Пальцы длинные и сильные, их проксимальные фаланги расширены, а на дистальных развивались мощные когти. Фаланговая формула, на первый взгляд, вполне маммального типа — 2 - 3 - 3 - 3 - 3. Однако при более детальном рассмотрении видно, что II-я фаланга IV-го пальца кисти сращена из двух: из них одна, сочлененная с проксимальной фалангой пальца, совсем короткая, а следующая, сочлененная с дистальной, сохранила длину, близкую к нормальной. Очевидно, у близких предков титанофонеев еще относительно недавно этот палец обладал 4 самостоятельными фалангами. Между пальцами могли иметься плавательные перепонки.

Прорисовка скелета правой кисти титанофонея (Titanophoneus potens).