Тринаксодоны — Thrinaxodon
Класс †Синапсиды (Synapsida)
Подкласс †Терапсиды (Therapsida)
Надотряд †Терозавры (Therosauria)
Отряд †Цинодонты (Cynodontia)
Клада †Эпицинодонты (Epicynodontia)
Семейство †Тринаксодоновые (Thrinaxodontidae)
Род †Тринаксодоны (Thrinaxodon)
Вид †Thrinaxodon liorhinus
Тринаксодоны (Thrinaxodon Seeley, 1894) — монотипический род семейства тринаксодоновых (Thrinaxodontidae Seeley, 1894), происходящий из раннего триаса Африки и Антарктики.
Реконструкция внешнего облика тринаксодона (Thrinaxodon liorhinus). Автор — Г. Угуэто.
Синонимы: Ictidopsis Broom, 1912; Notictosaurus Broom, 1936 (частью); Micrictodon Broom, 1937.
Наименование рода образовано древнегреческими словами θρῖναξ — "трезубые вилы" и ὀδούς — "зуб".
Частичный скелет тринаксодона (Thrinaxodon liorhinus) из раннего триаса провинции Фри-Стейт, ЮАР.
Единственный установленный вид — Thrinaxodon liorhinus Seeley, 1894 (тип). Его остатки обнаружены в ЮАР (провинции Восточная Капская, Фри-Стейт, Квазулу-Натал) и Антарктиде (Трансантарктические горы). Они датируются ранним триасом (зона Lystrosaurus, индский - оленекский ярусы). Синонимы: Ictidopsis elegans Broom, 1912; Ictidopsis formosa van Hoepen, 1916; Thrinaxodon putterilli Broom, 1932; Notictosaurus luckhoffi Broom, 1936 (частью); Micrictodon marionae Broom, 1937; Notictosaurus gracilis Broom et Robinson, 1948.
Многочисленные находки включают практически полные скелеты, что делает тринаксодона одним из наиболее изученных цинодонтов и синапсид в целом.
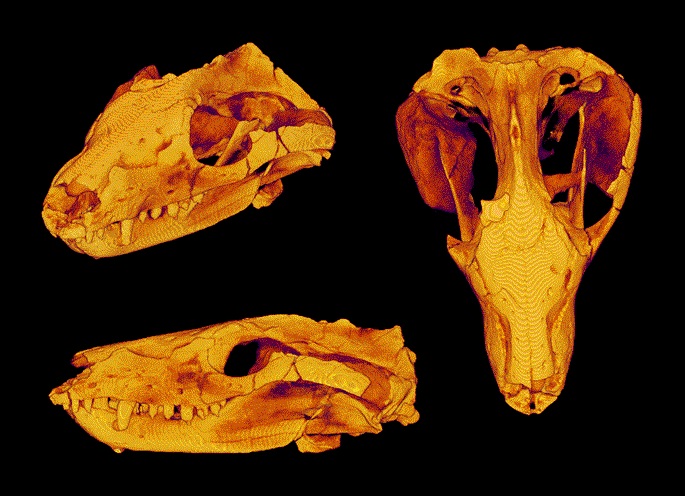
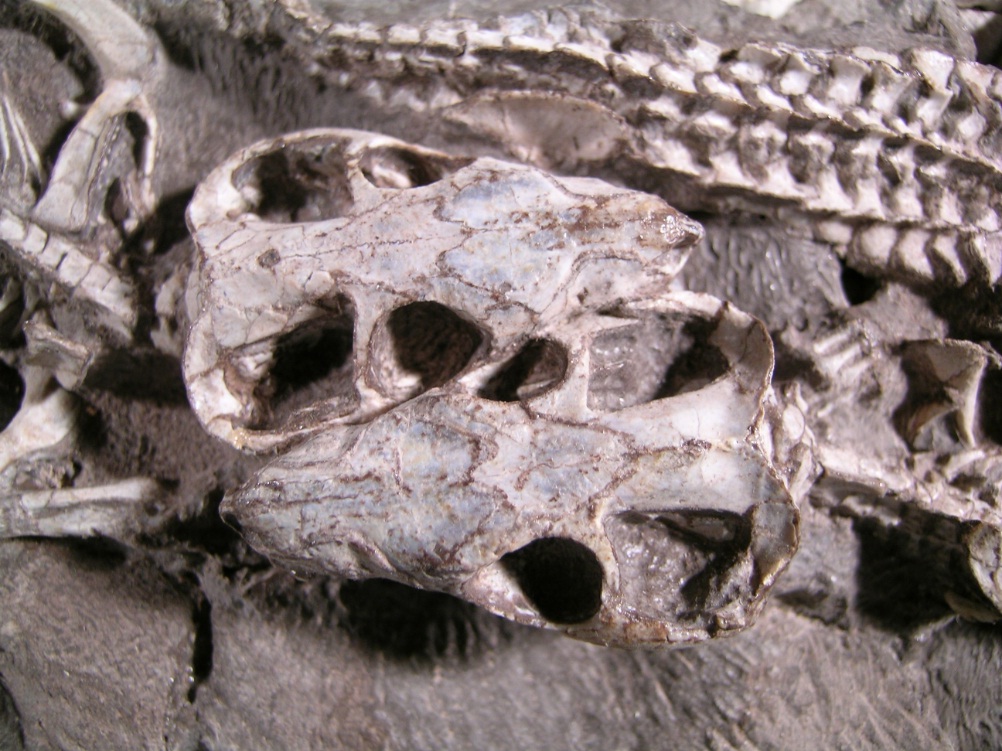
Череп с нижней челюстью тринаксодона (Thrinaxodon liorhinus) из ЮАР.
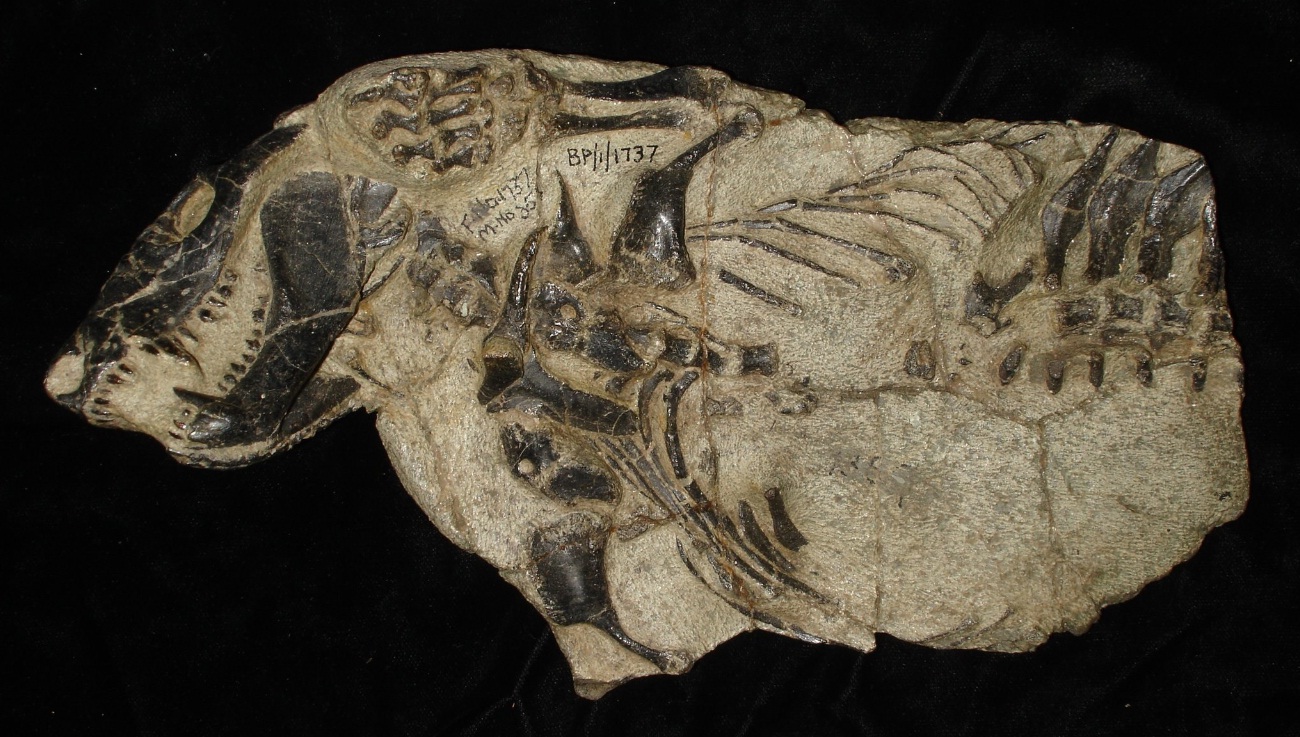
Череп с нижней челюстью и передняя часть скелета тринаксодона (Thrinaxodon liorhinus).
Череп с нижней челюстью и передняя часть скелета тринаксодона (Thrinaxodon liorhinus).
Скелет тринаксодона (Thrinaxodon liorhinus), погибшего свернувшимся в норе.
Часть позвоночного столба с ребрами тринаксодона (Thrinaxodon liorhinus).
Скелет тринаксодона (Thrinaxodon liorhinus) из отложений формации Фремоу, Антарктида.
Размеры небольшие: череп длиной около 10 см, общая длина составляет порядка 50 см.
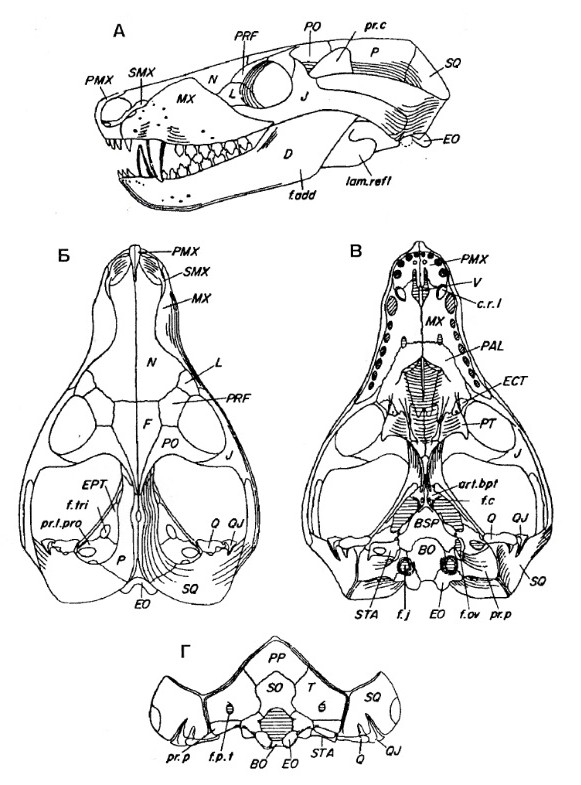
Череп крупный относительно тела. Глазницы большие. Заглазничная дуга тонкая. Височные окна очень велики и скуловые дуги широко выступают наружу. Лобные кости небольшие и входят в края глазниц. На месте заглазничной и заднелобной костей примитивных терапсид находится единственное окостенение, встречающееся с предлобной костью над глазницей. Очень узкая черепная коробка несет острый сагиттальный гребень, еще сохраняющий пинеальное отверстие. Почти половину длины черепа занимает увеличенная аддукторная камера. Чешуйчатая кость широко охватывает сзади аддукторную камеру и образует латеральную поверхность затылка. На затылке у заднего основания скуловой дуги имеется зачаточная вырезка. Яремные отверстия расположены низко на черепе. Затылочный мыщелок отчетливо парный.
Верхнечелюстные кости и передние части небных костей образуют шовное соединение по средней линии под носовым проходом, формируя сплошное вторичное небо. Кзади небные кости расходятся в стороны. У взрослых особей крыловидные кости встречаются на средней линии и закрывают межкрыловидную полость. В носовой полости имеются турбиналии.
Томограмма черепа с нижней челюстью тринаксодона (Thrinaxodon liorhinus).
Нижняя челюсть состоит в основном из зубной кости с хорошо развитым венечным отростком, заходящим дорсально выше скуловой дуги. Яма жевательной мышцы достигает его основания. Яма жевательной мышцы m. masseter достигает основания венечного отростка, где прикрепляется под m. temporalis. В отличие от высших цинодонтов, постдентальные кости редуцированы не сильно, но их задний край чуть приподнят по отношению к нижнему краю зубной кости. Эти кости утратили шовное соединение с зубной костью и могут двигаться отдельно от нее. Отогнутая пластина угловой кости продолжает оставаться крупным образованием, латерально выступающим над остальной костной поверхностью. Сочленовная кость полностью образует сочленение с черепом. Как и у Dvinia, на верхнезаднем крае сочленовной кости имеется утолщение, которое при сомкнутых челюстях соприкасалось с утолщением на наружном крае чешуйчатой или квадратноскуловой костей, благодаря чему достигалось упрочнение сустава. Маленькая квадратная и тесно связанная с ней квадратноскуловая кости сильно редуцированы и рыхло закреплены в вырезках у основания чешуйчатой кости.
Сверху и снизу имеется соответственно 4 и 3 резца, а также по довольно крупному клыку. Заклыковых зубов — от 7 до 9. Их коронки сжаты латерально и трехзубчаты по наружному краю, с главной центральной вершиной и передней и задней дополнительными. На заклыковых зубах взрослых особей имеется язычный поясок, не развитый у молодых. Передние заклыковые зубы построены проще. Наблюдается некоторое истирание зубов, но окклюзия отсутствует. По-видимому, все зубы (включая заклыковые) на протяжении жизни многократно сменялись.
Прорисовки реконструированного черепа с нижней челюстью тринаксодона (Thrinaxodon liorhinus): А — сбоку, Б — сверху, В — с неба, Г — сзади.
Телосложение легкое, напоминающее пропорциями куньих. Туловище удлиненное и способное сильно изгибаться в горизонтальной плоскости. Сохраняется проатлант. Имеются 7 шейных позвонков, отличающихся сохранением гипоцентров, утрачиваемых в туловищном отделе, относительно небольшим углом зигапофизов и простотой ребер. Удлиненное туловище включает 13 грудных, 6-7 поясничных и 5 крестцовых позвонков. Передние туловищные позвонки имеют дополнительные супразигапофизальные сочленения. На протяжении всего туловищного отдела позвоночника зигапофизы круто приподняты для уменьшения боковых изгибов.
Уже заметно четкое различие между грудным и поясничным отделами в длине ребер. Проксимальные части ребер разрастаются в расширенные реберные пластины, перекрывающие друг друга. Ребра поясничного отдела состоят из одних таких пластин без дистальных стержней. Реберные пластины логически связаны с усилением жесткости позвоночного ствола, позволяющей постоянно держать туловищный отдел над землей. Поясничный отдел позвоночника и реберные пластинки характерны и для высших цинодонтов. Относительно короткий хвост состоит из 10-15 позвонков.
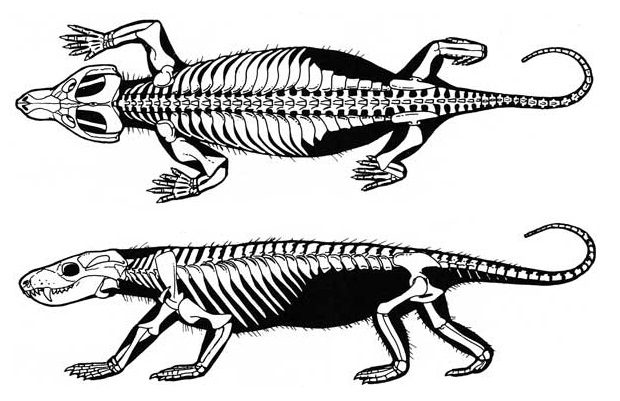
Прорисовки реконструированного скелета тринаксодона (
Thrinaxodon liorhinus), вписанного в телесные очертания. Автор — Д. Петерс.
Коракоид и лопатка принимают примерно одинаковое участие в формировании гленоидной впадины, а небольшой прокоракоид образует узкий участок ее переднего края. Лопатка длинная и тонкая; она вогнута медиально в соответствии с конфигурацией туловища. Передний край лопатки редуцирован, но вперед отходит короткий отросток, акромион, сочленяющийся со стержнем ключицы. Лопаточная часть гленоидной впадины ориентирована вентрально и латерально, вероятно принимая на себя большую часть веса тела, действующего на переднюю конечность. Вентральная часть таза не пластинчатая: лонная кость укорочена, а седалищная вытянута назад.
Конечности укороченные, передние широко расставлены в стороны, но задние уже несколько сближены. Плечевая кость остается тяжелой и сложной. Ее головка отклонена дорсально для более эффективной передачи силы от конечности к вертикальной лопатке. Вертлужная впадина относительно открыта спереди и снизу. Головка бедренной кости отклонена от диафиза вверх и вперед. Обе берцовые кости ориентированы вертикально. Сочленовные поверхности берцовых костей видоизменены в соответствии с их более вертикальной ориентацией. Между голенью и стопой развилось простое сочленение с рычажной системой. Таранная кость прочно присоединена к большой берцовой, и основной сустав находится между таранной и пяточной костями, причем последняя интегрирована в стопу и развивает пяточный бугор. Животные было стопоходящим. Судя по всему, фаланговая формула составляет 2 - 3 - 4 - 4 - 3 (сохраняется по одной предпоследней фаланге дисковидной формы в III-м и IV-м пальцах).
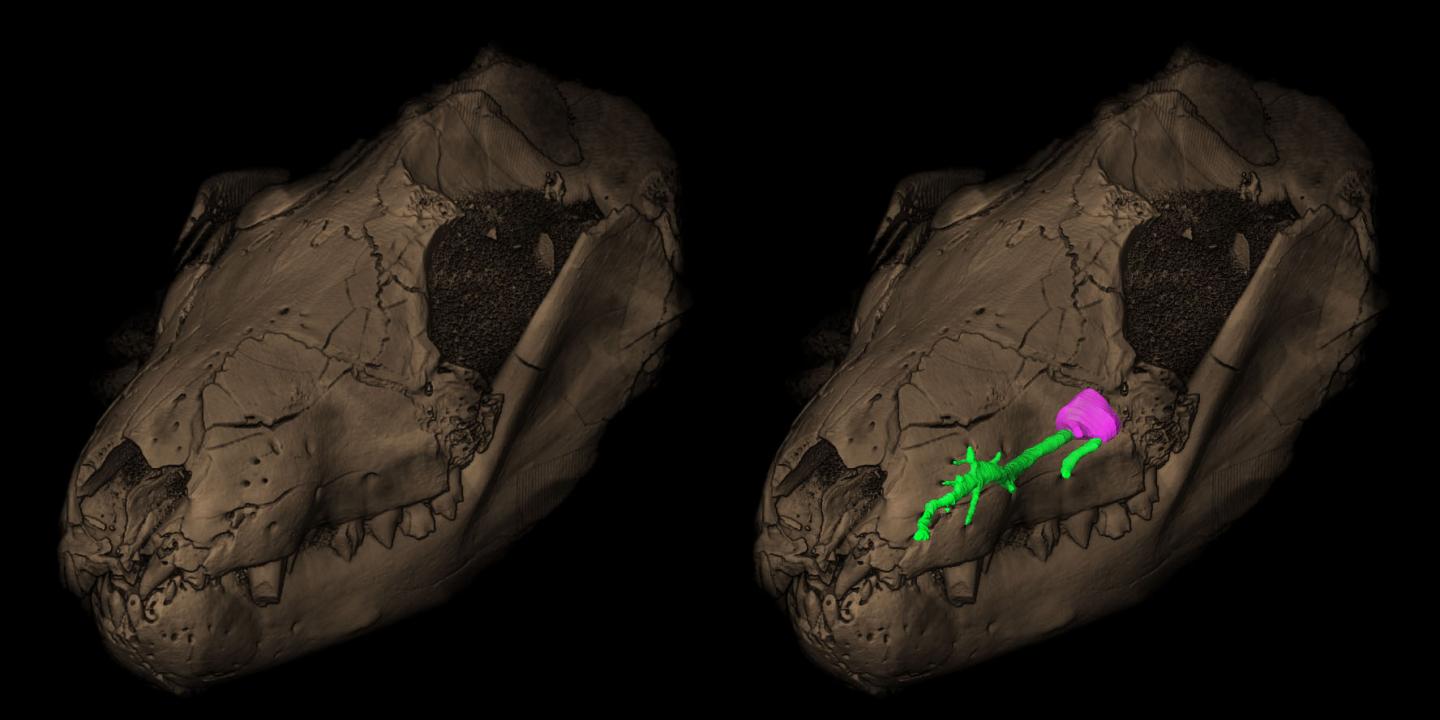
Томограмма черепа с нижней челюстью тринаксодона (Thrinaxodon liorhinus). Справа зеленым цветом выделен верхнечелюстной канал тройничного нерва, слева видны соответствующие ему ямки на морде.
Маленькие ямки по бокам морды позволяют предположить наличие вибрисс и как следствие то, что тело тринаксодона было покрыто шерстью. Между четко разделенными грудной и брюшной областью, вероятно, имелась мышечная диафрагма. Это может свидетельствует о прогрессивном типе дыхания и теплокровности, хотя со всей определенностью этого утверждать нельзя.
Блок породы с хорошо сохранившиеся скелеты 2 молодых особей тринаксодона (Thrinaxodon liorhinus), погибших в одной норе. Образец был найден Р. Смитом в 2000 г. на перевале Вападсберг в районе Грааф-Рейнет, ЮАР. Длина черепов составляет около 4 см.
Обширный ареал тринаксодона объясняется существованием в то время суперконтинента Пангеи. Судя по всему, он являлся активным хищником. Рацион составляли насекомые, другие безпозвоночные и мелкие позвоночные животные. Судя по большим глазницам, тринаксодон мог охотиться преимущественно по ночам. Также высказано предположение, что он был способен погружаться в глубокую спячку, помогавшую пережить сухие и голодные летние месяцы. Однако гистологический анализ костной ткани не выявил т. н. линий задержки роста, свидетельствующих о длительном бездействии и голодании. Это позволяет сделать вывод, что образ жизни тринаксодона позволял ему избегать неблагоприятных климатических факторов и обусловливал непрерывную скорость роста его организма. Таким образом, если тринаксодон и впадал в оцепенение, то эти периоды были относительно короткими по продолжительности.
В речных берегах и склонах холмов тринаксодон вырывал неглубокие норы, помогающие ему избежать хищников и защититься от неблагоприятных погодных условий. Норному образу жизни весьма способствовали гибкость позвоночника и устройство конечностей этого цинодонта. В слепках нор из отложений древних речных долин обнаружено множество следов роющей деятельности тринаксодонов, таких как царапины их когтей на стенках, а также найдены остатки самих животных. Обычно они погибали спящими, свернувшись калачиком. Известны находки погребенных в одном укрытии скелетов нескольких особей, включая пары (самец и самка), что может указывать на некоторую форму социального поведения. Найденные вместе скелеты крупных и мелких особей по-видимому представляют собой остатки матерей с детенышами, выводимых ими в подземных убежищах.

Самка тринаксодона (
Thrinaxodon liorhinus) присматривает за играющими детенышами. Автор — Д. Гендерсон.
В 1975 г. на перевале Оливирсшук в провинции Квазулу-Натал, ЮАР, был найден продолговатый слепок конечной камеры норы (образец BP/1/5558) из алевритового мелкозернистого песчаника с залегающими в нем обильными костными остатками, которые предварительно идентифицировали как принадлежащие тринаксодону. В 2013 г. данную окаменелость подвергли лазерному сканированию, в результате чего была получена ее подробная рентгенограмма. Она показала, что в слепке норы рядом со скелетом тринаксодона находится скелет молодой темносподильной амфибии брумистеги (Broomistega putterilli).
Остатки обоих животных прекрасно сохранились, включая узорчатую кожу амфибии. Наблюдаются лишь незначительные посмертные повреждения и смещения костных элементов, и только некоторые из них (часть фаланг и позвонков) утрачены. Тринаксодон лежит на брюхе, как бы прижавшись левым боком туловища к стенке норы. Его голова максимально вывернута влево, передние конечности согнуты по бокам от головы, задние вытянуты вперед, а хвост резко загнбается вправо. Расположенная брюхом вверх брумистега упирается в правый бок цинодонта. Эта ассоциация из двух очень разных животных поднимает вопрос о их возможном взаимодействии.
Естественный слепок неполной конечной камеры норы со скелетами тринаксодона (Thrinaxodon liorhinus) и брумистеги (Broomistega putterilli) (образец BP/1/5558). Находка составляет 28,1 см в длину и 15 см в самом широком месте.
Изучение остатков показало, что брумистега серьезно травмирована: у нее сломано 7 идущих друг за другом правых ребер. Можно было бы предположить, что тринаксодон напал на амфибию и затащил ее в свое логово. Однако сломанные ребра имеют признаки заживления и свидетельствуют о том, что ранение было получено несколько раньше. Вполне вероятно, что травма явилась результатом единственного сокрушительного удара — возможно, на брумистегу наступили, — произошедшего за несколько недель до смерти животного. Безусловно, это повлияло бы на его способность передвигаться и вызывало бы сильную боль, особенно при дыхании. Раненая амфибия, с трудом ковыляющая под палящим солнцем, была практически беззащитной перед хищниками.
Судя по всему, в поиске укрытия брумистега забралась в нору тринаксодона. Основываясь на строении конечностей брумистеги можно утверждать, что сама она не могла рыть почву. И наоборот, конечности тринаксодона вполне приспособлены для копания, что в сочетании с обнаружением других его экземпляров внутри их убежищ позволяет предположить, что нору создал именно он. Поскольку наружная поверхность слепка в некоторой степени эродирована, установить наличие на стенках камеры следов его когтей не представляется возможным. Поскольку какие-либо признаки ссоры отсутствуют, тринаксодон либо спал, либо терпел присутствие незваного гостя. Такие отношения являются примером комменсализма, возникающего при взаимодействии особей разных видов, когда одна из них (в данном случае амфибия) получает выгоду, не причиняя вреда или пользы другой.
Компьютерные модели скелетов тринаксодона (Thrinaxodon liorhinus) и брумистеги (Broomistega putterilli), найденных в одной норе: a — вид снизу; b — вид сверху. Длина масштабной линейки — 5 см.
С другой стороны, может оказаться, что тринаксодон находился в своей норе уже мертвым, а брумистега забралась туда и позднее также умерла, хотя исключительная сохранность и непосредственная связь их остатков позволяют предположить, что они погибли одновременно. Таким образом, следуя инстинкту выживания, брумистега искала защиту и забралась в нору, где, возможно, также погрузилась в сон. Подобное поведение наблюдается и у ныне живущих амфибий, особенно у молодых особей, которые порой укрываются в норах других животных.
Итак, скорее всего, тринаксодон спал в норе, когда к нему забралась раненая брумистега. Каким бы ни был точный сценарий, приведший к этой встрече, обоих животных постигло общее несчастье — внезапное наводнение быстро заполнило нору осадками. Судя по всему, течение прижало к стенке камеры тринаксодона, а также перевернуло брюхом вверх и вытолкнуло на его спину брумистегу. Данный случай дает непосредственное, редкостное представление о взаимодействии между совершенно не связанными друг с другом животными, которое просто не было бы рассмотрено, если бы не эта случайная находка.
Раненая брумистега (Broomistega putterilli) пытается скрыться от палящего солнца и находит нору со спящим тринаксодоном (Thrinaxodon liorhinus). Автор — Б. Николлс.
Ссылки
Thrinaxodon (Paleobiology Database)
перейти
Литература
Богданов Д. В. Звероящеры и другие пермские монстры. — М.: Фитон XXI, 2018
Вуд П., Вачек Л., Хэмблин Д. Дж., Леонард Дж. Н. Жизнь до человека. — М.: Мир, 1977
Кэрролл Р. Палеонтология и эволюция позвоночных. В 3 т. Т. 2 — М.: Мир, 1993
Основы палеонтологии. В 15 т. Т. 12: Земноводные, пресмыкающиеся, птицы / Гл. ред. Орлов Ю. А. — М.: Наука, 1964
Протеро Д. Отпечатки жизни. 25 шагов эволюции и вся история планеты. — СПб.: Питер, 2016
Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М.: ГеОС, 2009
Fernandez V., Abdala F., Carlson K. J., Cook D. C., Rubidge B. S., Yates A., Tafforeau P. Synchrotron Reveals Early Triassic Odd Couple: Injured Amphibian and Aestivating Therapsid Share Burrow. PLoS One 8 (6), 2013
Lomax D. R. Locked in time. Animal behavior unearthed in 50 extraordinary fossils. — New York: Columbia University Press, 2021