Тритилодонтовые — Tritylodontidae
Класс †Синапсиды (Synapsida)
Подкласс †Терапсиды (Therapsida)
Надотряд †Терозавры (Therosauria)
Отряд †Цинодонты (Cynodontia)
Клада †Эпицинодонты (Epicynodontia)
Инфраотряд †Эвцинодонты (Eucynodontia)
Клада †Пробайногнаты (Probainognathia)
Клада †Чиникводонтоиды (Chiniquodontоidea)
Клада †Прозостродонты (Prozostrodontia)
Семейство †Тритилодонтовые (Tritylodontidae)
Тритилодонтовые (Tritylodontidae Соре, 1884) — семейство клады прозостродонтов (Prozostrodontia Liu et Olsen, 2010), происходящее из позднего триаса - средней юры Европы, ранней юры Северной Америки, ранней юры Африки, ранней юры Антарктиды, ранней юры - раннего мела Азии. Установлено порядка 20 родов.

Реконструкция внешнего облика кайентатерия (Kayentatherium wellesi). Автор — Г. Лутерман.
Высокоспециализированная группа, перешедшая к растительноядному питанию и по ряду признаков значительно приблизившаяся к млекопитающим. Самые прогрессивные и последние известные по времени терапсиды.
Размеры варьируют от мелких до довольно крупных, в большинстве случаев небольшие.

Череп с нижней челюстью кайентатерия (Kayentatherium wellesi) из раннеюрских отложений формации Кайента, штат Аризона, США.
Череп внешне сильно напоминает таковой млекопитающих. Его лицевая часть заметно укороченная, низкая и узкая. Септомаксиллы нет. Носовое отверстие непарное. Заглазничная дуга вместе с предлобной, заднелобной и заглазничной костями утрачены. Края глазницы образуют лобные кости. Глазница сливается с увеличенным височным окном. Скуловые дуги широко выступают наружу. Скуловая дуга ориентирована парасагиттально, она очень высокая, образованная, главным образом, скуловой костью, ее нижний край ровный.
Теменные кости длинные и узкие. Они почти всегда образуют очень мощный сагиттальный гребень, разрастающийся впереди на лобные кости почти на всем их протяжении. Лобные и теменные кости с хорошо развитыми нисходящими флангами. Предушная стенка мозговой коробки образована алисфеноидом и орбитосфеноидом, впереди которого лобная кость контактирует с восходящей пластиной небной кости. Пинеальное отверстие отсутствует. Желобок наружного слухового прохода на чешуйчатой кости хорошо выражен и изогнут вперед. Заметно редуцированная квадратная кость рыхло соединена с чешуйчатой. Затылочный мыщелок парный.
Вторичное костное небо развито очень хорошо, образуясь максиллами и небными костями. Задний край неба заходит за уровень переднего края глазницы, располагаясь близ уровня последних максиллярных зубов. Небные кости широко сливаются по средней линии. Небные зубы отсутствуют. Эктоптеригоидов нет. Поперечные отростки птеригоидов короткие, на конце они направлены назад. Базиптеригоидное сочленение неподвижное, образовано птеригоидом и базисфеноидом (возможно, без участия птеригоидов). Квадратные ветви пгеригоида утрачены (по крайней мере, в некоторіх случаях). Межптеригоидных ям и небных отверстий нет. Верхняя крыловидная кость расширена и соединяется швом с передней ушной костью.
Нижняя челюсть почти целиком образована разросшейся массивной зубной костью. Ее очень массивный венечный отросток приподнят вертикально, с вогнутыми верхним и задним краями; самостоятельная венечная кость отсутствует. Массетерная площадка очень крупная, по нижнему краю венечного отростка доходит до его переднего конца. Мыщелковый отросток не достигает чешуйчатой кости. Задние кости нижней челюсти редуцированы и расположены в желобке на внутренней поверхности зубной кости под задним основанием венечного отростка, намного выше нижнего края главной (вентральной) части нижней челюсти. Иногда указывается на наличие дополнительного челюстного сочленения между чешуйчатой и зубной костями, что оспаривается другими авторами.
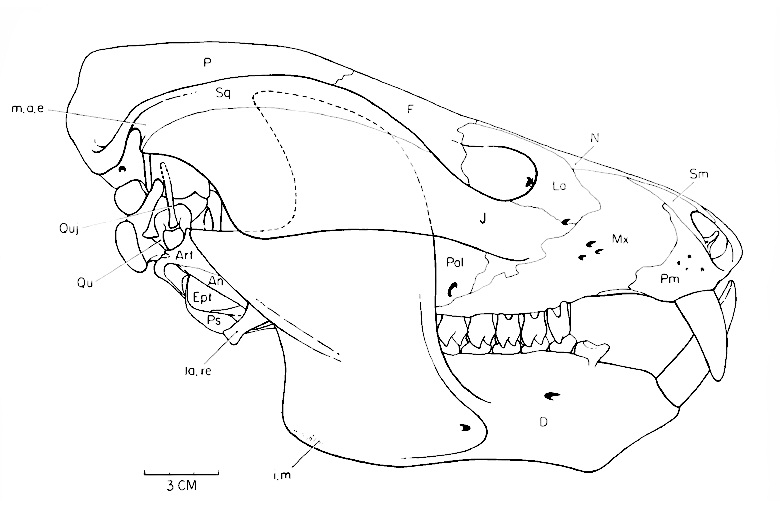
Прорисовка реконструированного черепа с нижней челюстью кайентатерия (Kayentatherium wellesi).
Зубная система довольно сходна с таковой грызунов. Зубная формула: I1-3/i1-3, CО/с0, Рс5-9/рс5-9.
У примитивных форм резцы конические, выпуклые лабиально и вогнутые лингвально. I-я пара резцов, особенно на нижней челюсти, может выступать вперед. Зачастую I-е пары верхних и нижних резцов утрачиваются, а II-е сильно увеличиваются и превращаются в миниатюрные клыкоподобные бивни, зачастую выступающие вперед. При этом нижние резцы проходят впереди верхних, а передний конец премаксилл становится беззубым. Помимо I-й, иногда исчезает и III-я пара резцов.
Клыков нет, и резцы отделены от заклыковых зубов выраженной диастемой, размеры которой связаны с длиной морды: диастема длиннее у длинномордых форм и намного короче у короткомордых.
Заклыковые зубы образуют продольный ряд, смещенный лингвально даже больше, чем у гомфодонтных цинодонтов. Заклыковые зубы моляроподобные — их коронки низкие и расширенные, почти квадратной формы. В отличие от всех прочих терапсид, но как и у млекопитающих, у заклыковых зубов раздвоенные, очень длинные корни, порой доходящие в нижней челюсти до ее нижнего края и значительно укрепляющие посадку зуба.
Коронки заклыковых зубов несут 2 или 3 продольных ряда бугорков в верхней челюсти и 2 в нижней. Ряды бугорков разделены глубокими продольными бороздками — соответственно, 1-2 на верхних и 1 на нижних зубах. Отдельные бугорки серповидные; на верхних зубах они вогнуты спереди, на нижних — сзади. Иногда на верхних зубах бугорки лингвального и буккального рядов превращаются из серповидных в гребневидные.
На верхних заклыковых зубах в центральном продольном ряду, а иногда и в лингвальном развит добавочный бугорок. При этом если в первом случае зубы еще можно считать аберрантными двурядными, то во втором случае их лучше называть аберрантными трехрядными. На нижних зубах также зачастую развито по 4 бугорка в каждом продольном ряду. Бугорки образуют и поперечные ряды, которые, однако, расположены менее правильно, чем продольные.
Нередко утверждается, что движения нижней челюсти у тритилодонтов при питании были пропалинальными — т. е., нижние зубы двигались назад вдоль борозд между верхними зубами. Однако прикус у хорошо изученных представителей был строго ортогональным, что свидетельствует скорее на ортогональность и окклюзию. Ни в одном случае челюстной сустав не был изучен детально, что оставляет свободу для интерпретаций. Для одних форм указывается строго параллельное направление бороздок истирания на бугорках зубов, тогда как у других бороздки истирания направлены строго вертикально. Вопрос, по-видимому, нуждается в дополнительном изучении, но сомнительно, чтобы у разных тритилодонтов был такой различный прикус.
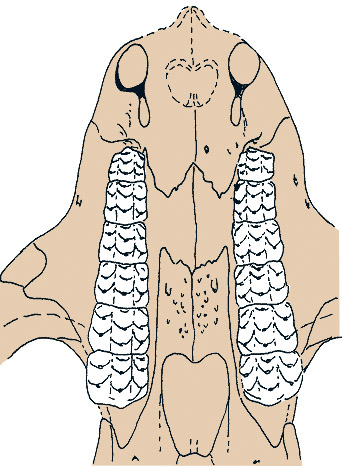
Прорисовка небной поверхности черепа кайентатерия (Kayentatherium wellesi).
Тело эпистрофея с крупным передним зубовидным отростком. Дуга атланта и гипоцентр еще не слиты друг с другом. Концы тел плоские (амфиплатные), как у млекопитающих, с утратой между ними хордальных остатков. Всего предкрестцовых позвонков 26, все они (кроме атланта) несут ребра. Расширенных реберных пластинок нет. Имеются хорошо обособленные шейный отдел из 7 позвонков и поясничный из 3 позвонков. Поясничный отдел хорошо выражен, но в нем сохраняются постепенно укорачивающиеся ребра, так что его передняя граница может быть определена только условно. Крестцовых позвонков, по различным данным, от 2 до 4. Хвостовые позвонки удлинены и образуют длинный, но тонкий хвост.
Пояса конечностей по строению очень близки к таковым однопроходных млекопитающих. В плечевом поясе есть 2 коракоидных элемента, редуцированных сильнее, чем у ранних цинодонтов, и, по-видимому, сращенных с лопаткой. Лопатка немного расширена, с гребнем, отделяющим ее верхнезаднюю часть. Гленоидная впадина очень широко открыта наружу. Хорошо развитый акромиальный отросток и основание переднего края пластины лопатки отклонены латерально от переднего края коракоидной области. Грудина окостеневает.
Таз принимает маммальную ориентацию. Подвздошная кость длинная и направлена резко вперед и вверх, а ее продолжение назад отсутствует. Седалищная кость расположена явно позади подвздошной. Маленькая лобковая и седалищная кости развернуты назад так, что большей частью находятся позади вертлужной впадины. Данная впадина расположена у верхнего края таза вне контакта с лобковой костью, она глубокая, но относительно маленькая. Обтураторное отверстие большое. Известны сумчатые кости.
Конечности короткие. По-видимому, передняя их пара несколько расставлена в стороны, тогда как задняя располагалась парасагиттально. У плечевой кости длинный диафиз и крупная полусферическая головка. Бедренная кость практически прямая с крупной, медиально загнутой полусферической головкой, отклоненной внутрь. Хорошо развиты большой, малый и внутренний грохантеры. Пяточная кость не образует пяточного бугра. Фаланговая формула маммального типа: 2 - 3 - 3 - 3 - 3.
В отличие от млекопитающих, в процессе роста организма тритилодонтов аллометрии не наблюдается, то есть пропорции тела у новорожденных и взрослых особей практически не отличаются. В значительной мере это объясняется маленькими размерами головного мозга, так что череп новорожденных небольшой, а не увеличенный, как у детенышей млекопитающих.
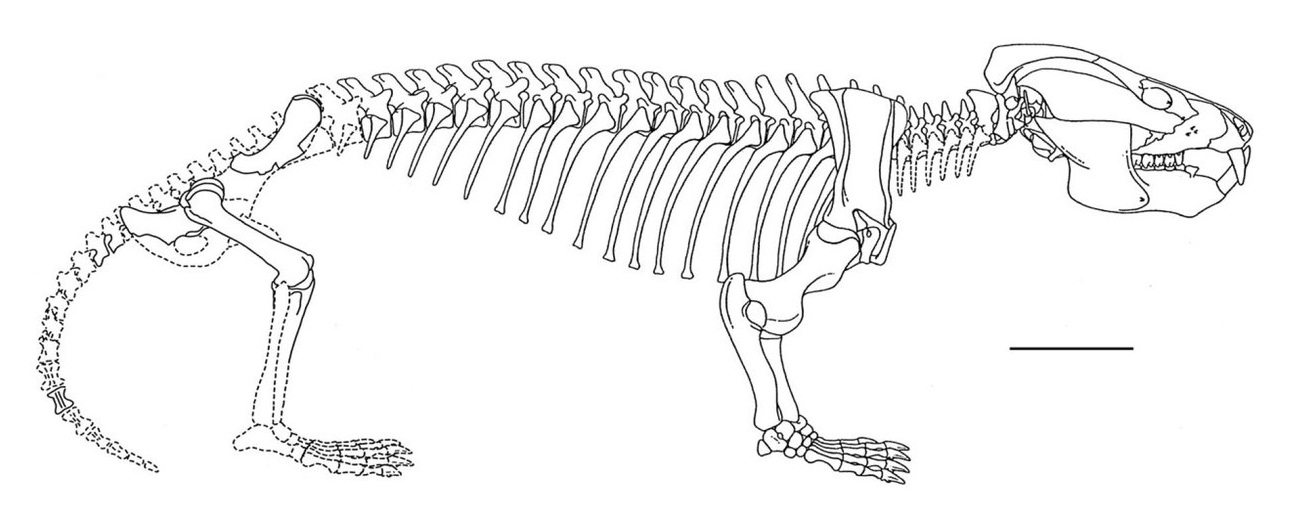
Прорисовка реконструированного скелета кайентатерия (Kayentatherium wellesi). Длина масштабной линейки — 10 см.
Тритилодонты употребляли в пищу мягкую растительностьу, такую как стебли, листья и корни. Их выступающие вперед резцы, заклыковые зубы с рядами бугорков и мощные жевательные мышцы формировали весьма совершенный аппарат для перетирания растительного корма. Долгое время никто другой не мог составить тритилодонтам конкуренцию на этом поприще: самые мелкие растительноядные динозавры были размером с зайца, а среди прочих диапсидных рептилий фитофагов было не так уж много. Так что данная группа спокойно занимала свою особую экологическую нишу, сравнимую с таковой современных грызунов. В то время как большинство насекомоядных и хищных цинодонтов вымерло в раннем юрском периоде, тритилодонты благополучно прожили на Земле еще почти 100 млн. лет. Вероятные причины их исчезновения — климатические изменения, развития новых типов растительности и невозможность конкурировать с растительноядными представителями бурно эволюционирующего отряда многобугорчатых (Multituberculata).
Древнейшие безусловные тритилодонты известны из позднего триаса (рэтский ярус) Западной Европы. Не строго определимые посткраниальные остатки также происходят из позднего триаса (норийский ярус) Аргентины. Самых древних тритилодонтов из Китая, происходящих из нижних горизонтов формации Нижний Люфен, обычно относили к самому позднему триасу, но теперь склонны датировать ранней юрой. Наиболее разнообразная фауна тритилодонтов происходит из юрских отложений китайских провинций Юньнань, Сычуань и Синьцзян-Уйгурского автономного района. Помимо Китая, юрские тритилодонты известны из Африки (ЮАР), Северной Америки (США и Мексика) и Западной Европы (Англия и Шотландия), а одна форма определена из ранней юры Антарктиды. Последние во времени представители группы описаны из раннего мела Кемеровской области, Красноярского края и Якутии в России, а также Японии.
Тритилодонты четко обособлены от всех других цинодонтов, что ставит под вопрос их родственные связи. Они обладают определенными чертами сходства с млекопитающими, и первоначально исследователи причисляли их к последним — поначалу из-за подразделенности корней заклыковых зубов (Оуэн (1848 г.)), потом из-за сходства заклыковых зубов — к многобугорчатым (Брум (1910, 1932 гг.), Симпсон (1928, 1929 гг.), Юнг (1940 г.)). Еще в первом издании руководства А. Ромера (1933 г.) тритилодонты рассматривались как подотряд многобугорчатых. Но к середине прошлого столетия принадлежность тритилодонтов к терапсидам стала общепризнанной (Симпсон (1937 г.), Ватсон (1942 г.), Паррингтон (1947 г.), Юнг (1947 г.), Кюне (1956 г.), Ватсон и Ромер (1956 г.)).
Позднее тритилодонтов обычно пытались выводить из высших гомфодонтных цинодонтов, но с ними их сближают лишь немногие четкие апоморфии: прежде всего гомфодонтность зубов, расположенных в хорошо сформированных альвеолах; также продольная ориентация ряда заклыковых зубов, сдвинутых несколько лингвально на уровень поперечных отростков птеригоидов; и, кроме того, возможно, утрата самостоятельного эктоптеригоида. А. Кромптон и Ф. Элленберджер (1957 г.) — авторы гомфодонтной теории происхождения тритилодонтов, считали их ближайшими родственниками цинодонтов семейства Traversodontidae. По их мнению, у тритилодонтов, как у траверсодонгид, нижние заклыковые зубы при смыкании челюстей смыкались одновременно с двумя верхнечелюстными зубами, хотя следует отметить, что и У. Кюне (1956 г.), и Дж. Хопсон (1964 г.) изображают прикус тритилодонтов в ортогональной проекции без перекрытия нижними заклыковыми зубами двух верхних.
Передний поперечный гребень на зубах тритилодонтов А. Кромптон и Ф. Элленберджер сопоставляют с острым гребнем, развитым у переднего края на зубах примитивного траверсодонтида Scalenodon. В последующей работе А. Кромптон опирается на сравнение тритилодонтов с более типичным траверсодонтидом Massetognathus, у которого на заднем крае верхних заклыковых зубов намечается, помимо лабиальной и лингвальной, также центральная вершина, а дополнительно имеется и лабиальный гребень. 2 передние вершины нижних зубов входили у Massetognathus, как у тритилодонтов, впереди заднего гребня верхних зубов по сторонам центральной вершины, то есть на уровне, на котором у тритилодонтов на верхних зубах появляются продольные бороздки, разделяющие продольные ряды бугорков. Лабиальный гребень на верхних зубах Massetognathus сравнивается с лабиальным продольным рядом бугорков на зубах тритилодонтов, центральный бугорок на заднем крае зубов Massetognathus предположительно дал начало центральному продольному ряду у тритилодонтов, лингвальный бугорок сравнивается с лингвальным рядом зубов тритилодонтов.
Хотя в принципе подобное превращение траверсодонтовых зубов в тритилодонтовые возможно, остается открытым ряд вопросов, касающихся, в частности, путей преобразования простых траверсодонтовых бугорков в серповидные тритилодонтовые, расположенные продольными рядами. Неясность путей этого преобразования подчеркивает и то обстоятельство, что при сравнении со Scalenodon дополнительные тритилодонтовые бугорки на верхних зубах должны были возникать на уплощенном талоне, расположенным кзади от переднего поперечного гребня, а при сравнении с Massetognathus они возникают кпереди от поперечного гребня, расположенного на заднем крае коронки. В любом случае, слишком смелым представляется утверждение, что зубы Scalenodon в передней части почти идентичны тритилодонтовым, или что двухкорневые зубы тритилодонтов свидетельствуют о возможном попарном срастании зубов траверсодонтид. Тем не менее, теория гомфодонтового или траверсодонтового происхождения тритилодонтов получила почти всеобщее распространение (Хопсон и Китчинг (1972 г.), Сюез (1985 г.), Хопсон и Бархьюзен (1986 г.), Баттайл (1991 г.), Хопсон (1991 г.)).
Однако другие исследователи (Татаринов (1974 г.), Кемп (1986 г.), Роу (1999 г.)) склонялись к сближать тритилодонтов с трителедонтами (иктидозаврами) — хотя Т. Кемп в последующем назвал гомфодонтовую гипотезу родства тритилодонтов тоже возможной, но менее вероятной, чем иктидозавровая. Все же на схемах Т. Кемп сначала склонен был помещать и тритилодонтов, и трителедонтов сразу вслед за прогрессивными триасовыми хищными цинодонтами семейства Chiniquodontidae, оставляя, таким образом, гомфодонтных цинодонтов в стороне. В отношении связей тритилодонтов с гомфодонтами Л.П. Татаринов (2009 г.) склонен согласиться с прежним мнением Кемпа, но считаю неясным вопрос о близости тритилодонтов и чиникводонтид. Возможно, предки как тритилодонтов, так и иктидозавров ближе стояли к малоизученным хищным цинодонтам типа Therioherpeton (Бонапарте и Барбарена (1975 г.)). Кемп (1988 г.) оставил тритилодонтов в неопределенном положении, посчитав одинаково возможным их родство и с Therioherpeton, и с траверсодонтидами. Согласно современным воззрениям, из всех терапсид тритилодонты наиболее близки к предкам млекопитающих.
Специализация зубной системы с отсутствием клыков и сложной моляризацией заклыковых зубов, несомненно, исключает всех известных тритилодонтов из предков млекопитающих. Высказано предположение, что у предшественников последних не могло быть зубной специализации тритилодонтов, но имелись производные посткраниальные признаки этой группы. Однако такие цинодонты до сих пор неизвестны. И напротив, можно допустить, что в строении конечностей и их поясов между предками млекопитающих и тритилодонтов наблюдалась значительная конвергенция.
Семейство имеет следующий состав:
? род †Завродесмы (Saurodesmus Seeley, 1891);
род †Олигокифы (Oligokyphus Hennig, 1922);
род †Кайентатерии (Kayentatherium Kermack, 1982);
род †Тритилодоны (Tritylodon Owen, 1884);
род †Биенотерии (Bienotherium Young, 1940);
род †Халепотерии (Chalepotherium Simpson, 1928);
род †Стереогнаты (Stereognathus Charlesworth, 1855);
род †Диннебитодоны (Dinnebitodon Sues, 1986);
род †Бокатерии (Bocatherium Clark et Hopson, 1985);
род †Люфенгии (Lufengia Chow et Hu, 1959);
род †Дянчжунии (Dianzhongia Cui, 1981);
род †Юньнанодоны (Yunnanodon Cui, 1986);
род †Биенотероидесы (Bienotheroides Young, 1982);
род †Юанотерии (Yuanotherium Hu, Meng et Clark, 2009);
род †Полистодоны (Polistodon He et Cai, 1984);
род †Нууртерии (Nuurtherium Velazco, Buczek et Novacek, 2017);
род †Шартегодоны (Shartegodon Velazco, Buzcek et Novacek, 2017);
род †Монтирикты (Montirictus Matsuoka, Kusuhashi et Corfe, 2016);
род †Фоссиоманусы (Fossiomanus Mao, Zhang, Liu et Meng, 2021);
род †Ксенокретозухи (Xenocretosuchus Tatarinov et Matchenko, 1999).
Ссылки
Новиковская А. Сибирские цинодонты. — 2023 (Элементы.Ру) перейти
Тритилодонтовые (Википедия) перейти
Tritylodontidae (Paleobiology Database) перейти
Литература
Богданов Д. В. Звероящеры и другие пермские монстры. — М.: Фитон XXI, 2018
Иванов А. О., Черепанов Г. О. Ископаемые высшие позвоночные. — СПб.: Издательство Санкт-Петербургского университета, 2001
Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Ч. 1 / Отв. ред. Ивахненко М. Ф. и Курочкин Е. Н. — М.: ГеОС, 2008
Кэрролл Р. Палеонтология и эволюция позвоночных. В 3 т. Т. 2 — М.: Мир, 1993
Основы палеонтологии. В 15 т. Т. 12: Земноводные, пресмыкающиеся, птицы / Гл. ред. Орлов Ю. А. — М.: Наука, 1964
Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М.: ГеОС, 2009