Трогонтерии — Trogontherium
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Териеобразные (Theriiformes)
Клада Голотерии (Holotheria)
Клада Трехнотерии (Trechnotheria)
Клада Кладотерии (Cladotheria)
Клада Затерии (Zatheria)
Клада Трибосфениды (Tribosphenida)
Клада Терии (Theria)
Клада Эвтерии (Eutheria)
Клада Плацентарные (Placentalia)
Магнотряд Бореоэвтерии (Boreoeutheria)
Суперотряд Эвархонтоглиресы (Euarchontoglires)
Грандотряд Глириобразные (Gliriformes)
Клада Глиресы (Glires)
Миротряд Симплицидентаты (Simplicidentata)
Отряд Грызуны (Rodentia)
Подотряд Боброобразные (Castorimorpha)
Надсемейство Кастороиды (Castoroidea)
Семейство Бобровые (Castoridae)
Подсемейство †Кастороидины (Castoroidinae)
Триба †Трогонтериини (Trogontheriini)
Род †Трогонтерии (Trogontherium)
Вид †Trogontherium minutum
Вид †Trogontherium minus
Вид †Trogontherium cuvieri
Трогонтерии (Trogontherium Fischer von Waldheim, 1809) — род подсемейства кастороидин (Castoroidinae Allen, 1877), происходящий из среднего миоцена - среднего плейстоцена Европы и раннего плиоцена - позднего плейстоцена Азии. Установлены 3 вида.
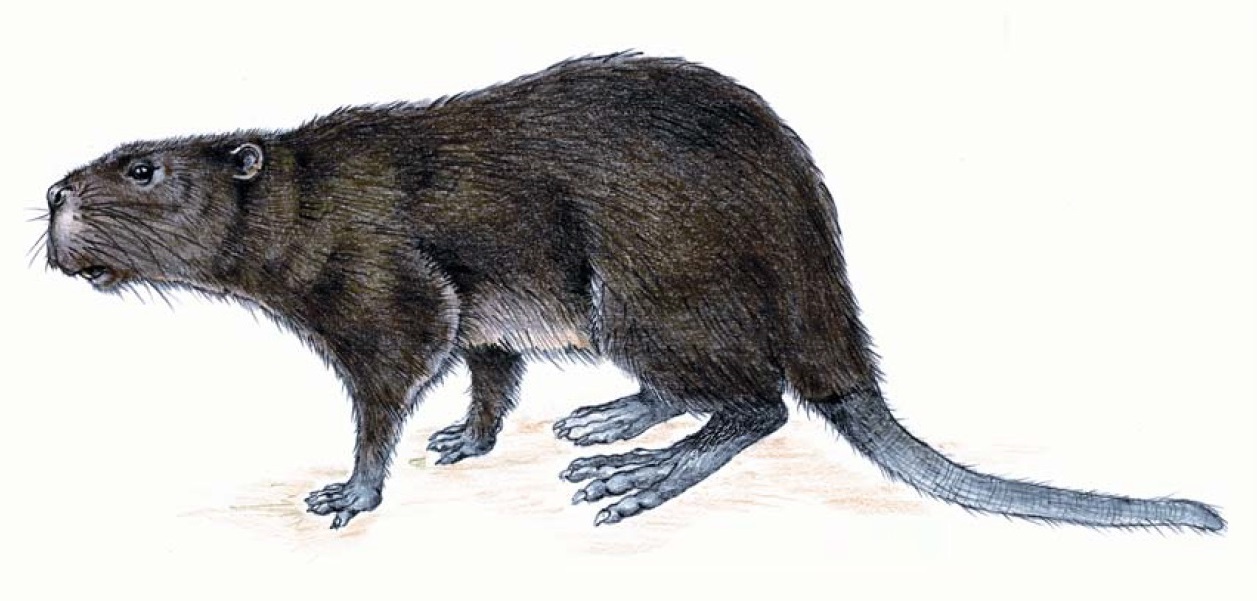
Реконструкция внешнего облика трогонтерия Кювье (Trogontherium cuvieri). Автор — А. Хольда-Михальска.
Синонимы: Castor Linnaeus, 1758 (частью); Diabroticus Pomel, 1848 (nomen nudum); Conodontes Laugel, 1862; Dipoides Schlosser, 1902 (частью); Sinocastor Young, 1934 (частью); Euroxenomys Samson et Radulesco, 1973 (частью).
История изучения
В начале XIX в. князь Александр Сергиевич Строганов передал Григорию Ивановичу Фишеру фон Вальдгейму неполный череп со щечными зубами крупного ископаемого бобра, найденный на побережье Азовского моря близ г. Таганрога. Этот череп происходил из личной коллекции Строганова, поэтому его возраст и слои, в которых был обнаружен, остались неизвестными. Принимая во внимания другие находки в этом районе, можно с достаточной долей уверенности датировать образец средним плейстоценом. Фишер фон Вальдгейм детально изучил череп и, сравнивая его с черепами ископаемых и современных бобров рода Castor, пришел к выводу, что по целому ряду признаков его нельзя отнести к данному роду.

Слепок типового черепа трогонтерия Кювье (Trogontherium cuvieri) из Приазовья, Россия.
Фишер фон Вальдгейм подробно описал находку в 1809 г., но не дал ей официального научного названия. Биноминальное наименование Castor trogontherium было дано и использовалось для обозначения этого животного только в работе Жоржа Кювье за 1812 г. Однако Кювье не упоминал Фишера фон Вальдгейма как автора первоописания до второго издания своей публикации в 1824 г., где уже использовалось название Trogontherium cuvieri. Родовое наименование образовано древнегреческими словами trogon — "грызущий, гложущий" и therion — "зверь".
Для Западной Европы остатки Trogontherium cuvieri впервые описал в 1846 г. Ричард Оуэн из местонахождений Лесного и Норвичского крагов в Англии. По своему возрасту эти отложения относятся к раннему плейстоцену.
В 1862 г. Огюст Ложель по неполному черепу с некоторыми зубами из местонахождения раннеплейстоценового возраста Сен-Пре в окрестностях Шартра на севере Франции описал новый род и вид бобра Conodontes boisvilletti. Это вызвало большие споры среди палеонтологов. По мнению ряда из них, новый вид является младшим синонимом Trogontherium cuvieri, что было доказано путем сравнения с английскими находками. Этой точки зрения большинство исследователей придерживалось и в дальнейшем. Досконально изучив вопрос, А. Шредер в 2008 г. показал, что Trogontherium cuvieri включает 2 хроноподвида — более древний Trogontherium cuvieri boisvilletti, населявший только Западную и Центральную Европу, и более молодой Trogontherium cuvieri cuvieri, широко распространенный от берегов Атлантики до Тихого океана.

Нижняя челюсть и верхнечелюстные фрагменты ранней формы трогонтерия Кювье (Trogontherium cuvieri boisvilletti) из Тегелена, Нидерланды.
С типовым видом в настоящее время также синонимизируют Dipoides lydekkeri (Шльоссер, 1902 г.) и Trogontherium soergeli (Рюгер, 1928 г.) из Европы, а также Sinocastor andersoni (Тейяр де Шарден, 1942 г.) из Китая (местность Чжоукоудянь неподалеку от Пекина). Diabroticus schmerlingi (Помель, 1848 г.), который был описан по остаткам Trogontherium cuvieri, признается таксоном nomen nudum.
Фон Мейер в 1838 г. описал из второй половины миоцена Баварии новый вид Trogontherium minutum. Э. Ньютон описал в 1890 г. из позднеплиоценовых отложений Красного крага, Англия, вид Trogontherium minus, от которого были найдены только верхнечелюстной фрагмент с несколькими зубами. По своему строению эти остатки имели сходство с Trogontherium cuvieri, но были значительно меньше по абсолютным размерам. Младшим синонимом данного вида сейчас также считают Trogontherium dacicum, описанного в 1972 г. Радулеску и Сэмсоном из позднего плиоцена юго-восточной части Румынии. Сегодня виды Trogontherium minutum и Trogontherium minus нередко относят к Euroxenomys, установленного Сэмсоном и Радулеску в 1973 г. и рассматриваемого в качестве подрода Trogontherium, или самостоятельного, более древнего предкового рода.
В 1955 г. Ян из провинции Цзянсу на востоке Китая описал по коренным зубам вид Trogontherium sinensis. Иследователь полагал, что формация Хсиатсаогвань, в отложениях которой он был найден, имеет плейстоценовый возраст. Однако в 1978 г. М. Чоу и Ц.-К. Ли переклассифицировали данного бобра в Youngofiber sinensis и показали гораздо более древний, ранне- и среднемиоценовый возраст слоев, из которых происходят его остатки.
Строение и функции
Величина трогонтериев варьировала от средней до крупной для грызунов, однако они не являлись настолько большими, как это порой утверждается в популярных источниках. Большая часть их ископаемых остатков по абсолютным размерам не превосходят соответствующие элементы скелета обыкновенного бобра (Castor fiber), и лишь некоторые образцы из раннего плейстоцена, принадлежащие наиболее крупному виду трогонтерию Кювье (Trogontherium cuvieri), несколько больше. Максимальная длина его черепа составляет 21 см, а общая длина тела могла достигать 150 см, что приблизительно на 20 % превосходит эти показатели у Castor fiber. Длина нижнего зубного ряда с IV-го предкоренного по III-й коренной включительно достигает 10 см, ширина верхнего резца доходит до 1,1 см.
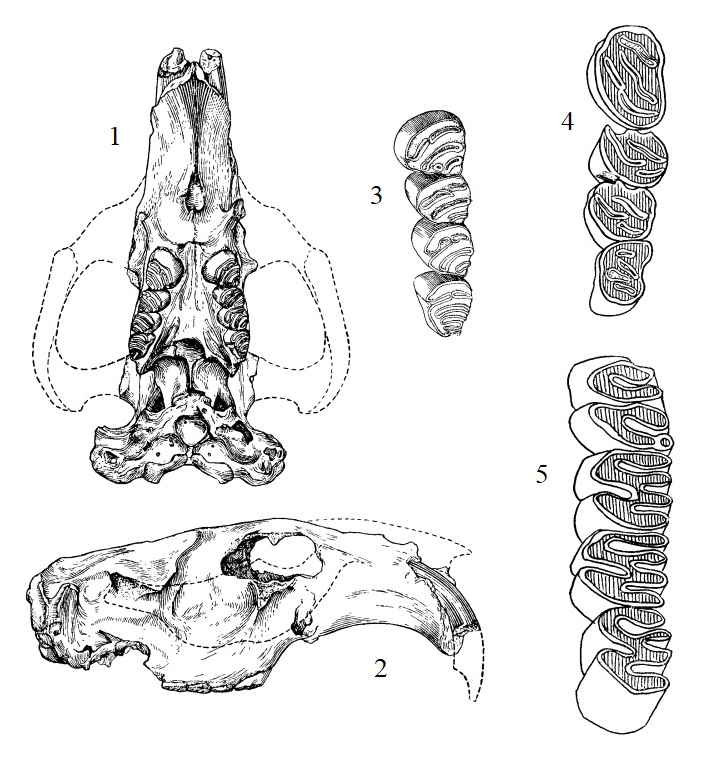
1, 2 — прорисовки типового черепа трогонтерия Кювье (Trogontherium cuvieri) из Приазовья, Россия; 3, 4 — прорисовки жевательной поверхности левых верхних и нижних коренных зубов трогонтерия Кювье (Trogontherium cuvieri), Казахстан; 5 — прорисовки жевательной поверхности правых верхних коренных зубов ранней формы трогонтерия Кювье (Trogontherium cuvieri boisvilletti).
Длина лицевого отдела черепа и резцов трогонтерии относительно больше, чем у дpyгих бобровых, при этом сам череп сравнительно узкий. На нижней поверхности основной затылочной кости находится глубокая впадина. Затылочное отверстие овальное. Твердое небо сходно с таковым у Steneofiber: без желобкообразных углублений позади резцовых отверстий, середина его заднего края расположена напротив середины III-го верхнего коренного. Особая форма передней части челюсти и шероховатость ее поверхности может указывать на наличие сильной и цепкой верхней губы, сходной с таковой ламантинов. Резцовая часть нижней челюсти направлена в сторону (наружу) и составляет с альвеолярной частью угол 70-80 °, тогда как угол расхождения ветвей нижней челюсти у бобров рода Castor не превышает 50 °. Передняя поверхность резцов в боковом направлении сильна выпуклая, в разрезе грушевидной формы, их эмаль у плио-плейстоценовых форм покрыта мелкими продольными бороздками. Коренные зубы с очень высокой коронкой и короткими корнями.
При том, что трогонтерии могли несколько превосходить современных бобров линейно, они не являлись настолько коренастыми. Абсолютная длина локтевой, лучевой, берцовых и метатарсальных костей больше, а пястные и плюсневые кости, а также фаланги пальцев длиннее и менее массивные, чем у Castor. В то же время, бедренная кость относительно меньше, чем у Castor, с высоко расположенным III-м вертлугом. В целом конечности относительно удлинены и, возможно, не имели плавательных перепонок, а хвост слабо сжат в верхне-нижнем направлении, т. е. округлый, а не плоский в сечении. Такое анатомическое строение может свидетельствовать о том, что трогонтерии зависели от водной среды в меньшей степени, чем современные бобры.
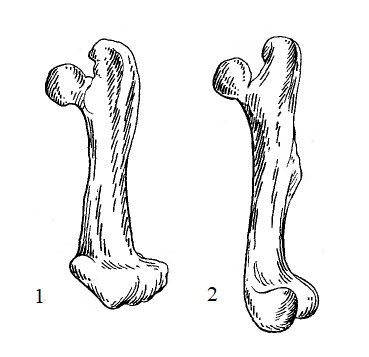
Прорисовки бедренных костей трогонтерия Кювье (Trogontherium cuvieri) и обыкновенного бобра (Castor fiber) из Тегелена, Нидерланды.
За миллионы лет своего существования трогонтерии не претерпели значительных морфо-анатомических изменений. В течение плиоцена, раннего и среднего плейстоцена размеры их тела постепенно увеличивались. Некоторые исследователи полагают, что со временем относительно больше становились резцы, другие указывают, что такое увеличение вполне укладывается в рамки видовой изменчивости. Особенно заметные изменения претерпевали щечные зубы, что, очевидно, было связаны с изменением пищевого рациона. Наряду с увеличением площади их жевательных поверхностей изменялись и особенности строения — складки эмали дольше оставались открытыми, появлялись дополнительные эмалевые складки на нижних предкоренных. Очевидно, изменения в строении зубов были связаны с изменением пищевого рациона животных. Половой диморфизм у трогонтериев выражен незначительно, самцы и самки имели примерно одинаковые размеры.
Возраст и распространение
Согласно имеющимся данным, рассматриваемый род сформировался в среднем миоцене (около 15 млн. лет назад) на территории Европы. На этом континенте он задерживается до второй половины среднего плейстоцена (порядка 200 тыс. лет назад). В Азию трогонтерии проникли в раннем плиоцене (примерно 5 млн. лет назад) и жили здесь почти до конца позднего плейстоцена. Самые молодые остатки, обнаруженные в окрестностях г. Харбин на северо-востоке Китая, имеют возраст около 40 тыс. лет.
Максимальное распространение и увеличение численности трогонтериев приходится на ранний плейстоцен. В это время их ареал простирался от Англии на западе до Северо-Восточной Сибири и Восточного Китая на востоке, а также от Нидерландов и Литвы на севере до Франции, Румынии, Турции, Закавказья, Казахстана, Кыргызстана и Южного Китая на юге. Таким образом, область распространения охватывала значительную часть Европейского континента, за исключением его северного региона и почти всего Средиземноморья. На территории Западной и Центральной Европы остатки этих бобров относительно многочисленны, тогда как на Русской равнине и, особенно, в Сибири, Средней Азии и Китае их известно немного. В Северной Америке остатки трогонтериев не обнаружены.
Состав
Trogontherium minutum von Meyer, 1838 — вид, известный из среднего - позднего миоцена (лангский - русцинийский ярусы, MN 5 - MN 15) Западной, Центральной и Восточной Европы (Франция, Испания, Португалия, Германия, Швейцария, Австрия, Словакия, Венгрия, Румыния, Украина и др.). Синоним: Euroxenomys minutus Samson et Radulesco, 1973. Является самым древним и наименьшим по величине представителем рода. Характерно наличие крупного верхнего III-го предкоренного, как у плио-плейстоценовых видов, но в отличие от них полностью отсутствуют полосы или шероховатости на эмали резцов.
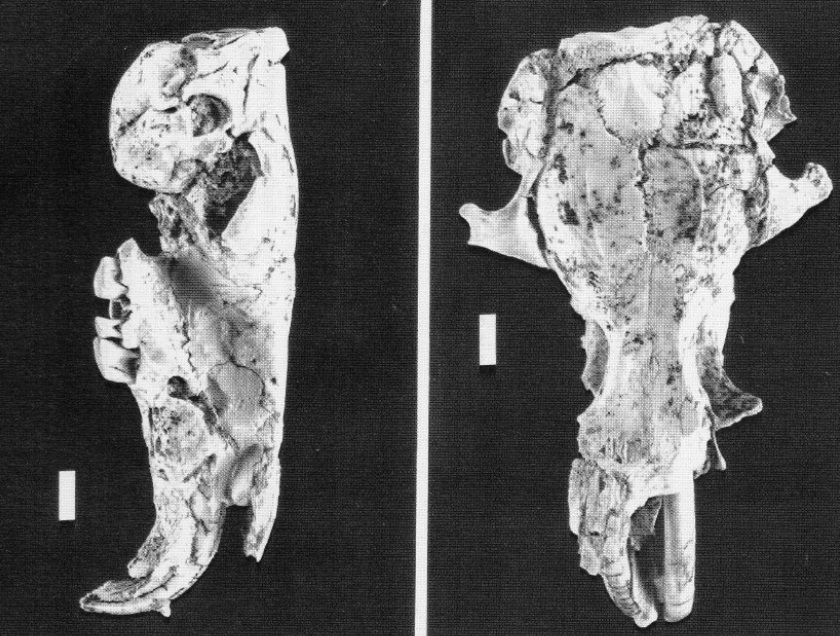
Череп трогонтерия (Trogontherium minutum) из позднего миоцена Петерсбаха, Южная Германия. Длина масштабной линейки — 0,5 см.
Trogontherium minus Newton, 1890 — вид, известный из плиоцена (верхи русцинийского - низы виллафранкского яруса, MN 14 - MN 16) Западной, Центральной и Восточной Европы, а также Средней Азии (Великобритания, Франция, Германия, Словакия, Румыния, юг европейской части России, Северный Казахстан и др.). Синонимы: Trogontherium dacicum Radulesco et Samson, 1972; Euroxenomys minus (Newton, 1890); Trogontherium minor. Размеры средние.

Фрагмент нижней челюсти трогонтерия (Trogontherium minus) из позднего плиоцена Одессы, Украина.
Trogontherium cuvieri (Fischer von Waldheim, 1809) — типовой вид, известный из позднего плиоцена (пьяченцский ярус) Средней Азии и Восточной Европы (Казахстан, юг европейской части России), раннего - среднего плейстоцена Европы и Азии (Великобритания, Франция, Германия, Нидерланды, Австрия, Италия, Чехия, Словакия, Венгрия, Румыния, Молдавия, Польша, Литва, Белоруссия, Украина, Россия, Турция, Казахстан, Китай и др.) и позднего плейстоцена Азии (Китай). Синонимы: Castor trogontherium Cuvier, 1812; Diabroticus schmerlingi Pomel, 1848 (nomen nudum); Conodontes boisvilletti Laugel, 1862; Dipoides lydekkeri Schlosser, 1902; Trogontherium soergeli Rüger, 1928; Sinocastor andersoni Teilhard de Chardin, 1942.
Размеры наиболее крупные в роду: кондилобазальная длина черепа составляет 12-21 см, длина лицевого отдела черепа превосходит 10 см. Носовой проход широкий и высокий. Глазница удлинена и смещена назад. Верхний край глазницы приподнят, отчего лобные кости также выпуклы, а не резко обрываются, как у Castor fiber. Надглазничные отростки лобных костей хорошо развиты.

Череп ранней формы трогонтерия Кювье (Trogontherium cuvieri boisvilletti).
Зубы крупные, радиус кривизны резцов превышает 7 см. Длина верхнего ряда коренных зубов по альвеолам более 4 см. IV-й верхний предкоренной длиннее, чем I-й верхний коренной; IV-й нижний предкоренной по длине приблизительно равен I-му нижнему коренному; III-и верхний и нижний коренные равны или длиннее, чем II-е. Складки эмали на верхних и нижних коренных зубах на ранних стадиях стирания замыкаются в мapки. Наружная складка эмали на нижних коренных заходит внутрь зубов дальше, чем на половину их ширины, а на IV-м нижнем предкоренном соединяется со второй (средней) внутренней складкой, разделяя зуб на две части. Складки на III-х верхнем и нижнем коренных располагаются в 4-5 поперечных ряда. Ранний подвид Trogontherium cuvieri boisvilletti отличается от более позднего Trogontherium cuvieri cuvieri несколько более тонкими резцами и особенностями строения коренных (в частности, более поздним замыканием эмалевых складок в марки).
Данный вид входит в состав 4 фаунистических комплексов, последовательно сменявших друг друга на Русской равнине: хапровского, псекупского, таманского и тираспольского. Он является одной из руководящих форм для раннего и I-й половины среднего плейстоцена и может быть использован для определения возраста континентальных толщ Европы и Азии.
Образ жизни
По мере накопления фактов об трогонтериях, начали появляться предположения об условиях их существования. Сходство морфо-анатомического строения скелета крупных трогонтериев с современными бобрами рода Castor надолго предопределило среди исследователей мнение, что трогонтерии также были водными животными. Многие исследователи, касаясь вопроса о местах их обитания, упоминали берега рек, ручьев, проточных озер и т. д. Среди китайских палеонтологов существовал даже специальный термин "большой речной бобр". Подобные выводы делались на основании общих соображений морфо-анатомического строения скелета. Отдельные морфологические отличия в строении костей трогонтериев и современных бобров обычно не рассматривались с точки зрения морфофункционального анализа.
Впервые иное толкование этому вопросу дал Б.С. Виноградов (1926, 1946, 1952 гг.). Изучая морфологические отличия в строении костей современных близких форм млекопитающих, он отмечал связь морфологических отличий с определенными условиями обитания животных. Виноградовым впервые было высказано мнение о том, что трогонтерии были сухопутными, роющими норы зверями. К этому выводу ученый пришел в результате исследования специфического строения их резцов. Н.И. Бурчак-Абрамович (1964 г.), изучая строение бедренной кости трогонтериев, также отметил, что, по-видимому, эти животные были больше связаны с сушей, чем настоящие бобры.
Б.С. Виноградов (1926 г.) установил на современных грызунах зависимость расположения резцов от образа их жизни. У животных, которые используют резцы только для того, чтобы грызть (например, представители семейства мышиных), верхние резцы, играющие преимущественно роль упора, когда они грызут более твердый субстрат, круто изогнутые, поставленные таким образом, что их свободный отпел располагается более или менее вертикально или даже изогнут назад. Нижние же резцы длинные, круто изогнутые, образующие с верхними тупой угол, близкий к 180°. Кроме того, резцы могут быть в той или иной мере приспособлены и к рытью, что находит свое выражение у некоторых грызунов, ведущих подземный образ жизни (слепушонка, слепыш), а в меньшей степени свойственно и многим норникам. В этом случае увеличивается длина свободного отдела резцов, особенно нижних, а верхние оказываются направленными вперед и образуют с нижними острый угол. При этом, как и в первом случае, общая длина резцов и угол, образуемый верхними и нижними резцами, больше у видов, имеющих дело с более твердым субстратом.
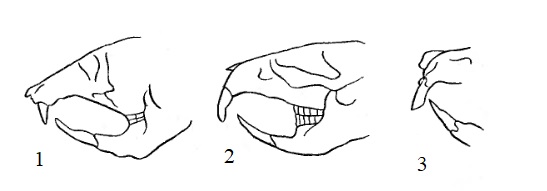
Прорисовки, демонстрирующие взаимное расположение резцов у различных грызунов при сомкнутых коренных: 1 — пеструшка; 2 — слепыш; 3 — желтый суслик.
Если сравнить расположение резцов у трогонтерия Кювье (Trogontherium cuvieri) и у обыкновенного бобра (Castor fiber), то легко заметить, что у последнего верхние резцы круто изогнуты, а свободный их отдел отклонен назад. Это не противоречит данным, изложенным выше, так как обыкновенный бобр является типично грызущим животным. По наблюдениям, сделанным в Березинском заповеднике, обыкновенные бобры подгрызают и валят осины диаметром до 0,5 м. Грызет бобр быстро. Деревце толщиной в палец он перегрызает одним ударом своих мощных резцов. Осину диаметром 5-7 см в течение ночи валит в количестве 4-5 шт. Из этих кратких данных четко видно, что резцы обыкновенного бобра приспособлены к грызению довольно твердого субстрата. Совсем иное у наблюдается у трогонтерия Кювье. Вертикальное расположение его верхнего резца свидетельствует о том, что животное грызло более мягкий субстрат, чем обыкновенный бобр.
У обыкновенного бобра кривизна верхних и нижних резцов значительно больше, чем у трогонтерия Кювье. Это позволяет предполагать, что этот грызун добывал пищу по-другому. Подтверждением этому выводу служит различие в строении режущей части резца трогонтерия и настоящего бобра. У последнего режущая часть нижних и верхних резцов имеет долотообразную поверхность, что вполне объясняется способом добывания пищи. У трогонтерия же нельзя назвать эту поверхность "режущей", так как резцы на концах заострены и имеют округлую форму. Грызть древесный субстрат, подобно обыкновенному бобру, при таком стирании резцов трогонтерий Кювье не мог. Однако, судя по большой абсолютной длине резцов и по мощной защитной эмали, они у трогонтерия интенсивно стирались. Можно думать, что при такой форме стирания резцов животное могло вонзать зубы в довольно твердый субстрат, не уступающий по твердости древесине ивы, осины или березы. Возможно, трогонтерии добывали корм, разрыхляя землю, т. е. питались корнями и клубнями растений.
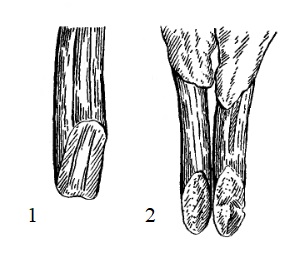
Прорисовки, показывающие характер стирания нижних резцов: 1 — трогонтерий Кювье (Trogontherium cuvieri); 2 — обыкновенный бобр (Castor fiber).
При таком способе добывания пищи у трогонтерия Кювье наблюдается увеличение абсолютной и относительной длины диастемы, которая служила животному преградой при попадании земли на коренные зубы. Установлено, что наибольшее удлинение диастемы происходит у грызунов, которые ведут подземный или водный образ жизни. При этом наибольшая абсолютная длина диастемы наблюдается у слепыша и слепушонки, в связи с необходимостью грызть корни под землей и прокладывать дорогу в почве при помощи резцов. Отсюда и следует ожидать большее увеличение диастемы у трогонтериев, которые, вероятно, добывали себе пишу с помощью резцов в почве, чем у настоящих бобров, у которых удлинение диастемы происходит от питания твердой пищей за счет редукции предкоренных зубов, подобно тому, как это имеет место у веткоядных копытных. Также отмечено, что редукция предкоренных зубов связана у грызунов с ослаблением режущей функции зубного аппарата.
Если сравнить ряды коренных зубов трогонтерия Кювье и обыкновенного бобра, то выясняется, что у последнего предкоренные зубы (особенно нижние) редуцированы более сильно. Поперечные гребни на верхних коренных больше направлены вперед у трогонтерия, чем у современного бобра. Предположительно, данный признак также указывает на большее развитие режущей функции зубов наряду с перетирающей. Это объясняется тем, что пища у современных бобров на коренные зубы попадает уже измельченная, благодаря работе резцов, и режущая функция коренных зубов в связи с этим притупляется. У трогонтериев пища, очевидно, попадала на коренные зубы предварительно не измельченная резцами. Кроме того, на коренных зубах у трогонтерия Кювье впереди каждого зуба развиты мощные корни, так как на них приходится основная нагрузка при жевании, преобладающее значение в котором имеет режущая функция зубов. Для ее усиления на нижних предкоренных зубах в передней части у трогонтерия Кювье появляется иногда даже дополнительная складочка эмали. Как видно, грызущий и жевательный аппараты трогонтериев и настоящих бобров имеют существенные отличия, которые можно объяснить различиями в добывании и переработке пищи этими животными.
Как уже было сказано, глазница у трогонтерия Кювье смещена больше назад, чем у обыкновенного бобра; верхний край глазницы приподнят и над ним развит мощный надглазничный отросток лобной кости. Замечено, что у обитателей более открытых биотопов, где возможен более широкий круговой обзор местности, глаза крупнее и сильнее смешены на боковые поверхности головы, тогда как у обитателей более закрытых биотопов, в условиях которых зрение ограничено в боковых направлениях, глаза мельче и больше направлены вперед. Смещение глазницы назад у трогонтерия, очевидно, связано с расположением глазного яблока у этого животного больше на боковой поверхности головы, чем у обыкновенного бобра, у которого глазное яблоко размещено в передней части глазницы, а сама глазница смещена более вперед.
У одного из черепов трогонтерия Кювье длина глазницы равна 72,5 мм, ширина — 29,8 мм; у соразмерного черепа обыкновенного бобра эти же промеры соответственно равны 52,5 мм и 22,5 мм. Судя по размерам глазниц, можно думать, что глазное яблоко трогонтерия Кювье было крупнее, помещаясь в более крупной орбите и имело сильную защиту в виде прикрывающего его приподнятого верхнего края глазницы и мощных надглазничных отростков. У обыкновенного бобра все эти признаки развиты в очень слабой степени. Он имеет небольшие, мало заметные глаза и не обладает хорошим зрением. Таким образом, глаза у трогонтерия Кювье были, очевидно, приспособлены к обзору более открытых пространств, чем у обыкновенного бобра.
Важно отметить, что у трогонтерия Кювье очень хорошо развиты носовые кости. Они длиннее и почти вдвое шире, чем у обыкновенного бобра. Возможно, такое строение носовых костей у трогонтерия было связано с развитием эндотурбиналий, т. е. наличием хорошо развитого обоняния, и, может быть, специфических особенностей дыхания (в частности, с фильтрующей ролью носовых этмоидных раковин).
Некоторые отличия наблюдаются в строении конечностей трогонтерия Кювье и современного бобра. Так, у трогонтерия, как отмечалось выше, III-й вертлуг бедренной кости сдвинут наверх, что является характерным признаком, отражающим приспособление животного к бегу. У обыкновенного бобра III-й вертлуг развит на середине тела бедренной кости.
Как известно, при выработке приспособлений к бегу удлиняются в большей степени дистальные отделы конечностей, чем плечевые и бедренные кости. В одном из случаев длина сросшихся берцовых костей у трогонтерия равна 153 мм, в то время как у обыкновенного бобра она составляет лишь 142 мм. Сходные же результаты можно наблюдать сравнивая длину метатарсальных костей. Абсолютная длина лучевых, локтевых, пястных, плюсневых костей и фаланг пальцев также больше у трогонтерия, чем у современного бобра.
Все эти данные свидетельствуют о том, что трогонтерий Кювье был приспособлен к бегу значительно лучше, чем обыкновенный бобр, который в связи с медлительностью движений редко удаляется от воды на сколько-нибудь значительное расстояние. Дело в том, что бегущего бобра без труда может догнать человек, не говоря уже о хищниках, например о волке или собаке.
Следует также отметить, что фаланги пальцев задней конечности у трогонтерия Кювье имеют более округлую форму, в то время как у обыкновенного бобра они более плоские, что связано с наличием у последнего хорошо развитых перепонок. Обыкновенный бобр зачастую роет несколько нор длиной 4-10 м. Особенности же строения конечностей трогонтерия показывают, что он мог рыть землю лучше, чем современный бобр.
Основываясь на изложенных выше данных, А.Н. Мотузко (1972 г.) делает вывод что характерные морфологические черты строения скелета трогонтерия Кювье отнюдь не являлись следствием приспособления животного к жизни в воде. Судя по всему, трогонтерии неплохо бегали, могли рыть длинные норы, добывали пищу из земли, очевидно, имели хорошее зрение и обоняние. По всем этим признакам животное вполне могло населять полуоткрытые лесостепные, или даже открытые степные ландшафты — т. е., достаточно засушливые районы.

Трогонтерий Кювье (Trogontherium cuvieri) в естественном окружении. Автор — Р. Евсеев.
Некоторые исследователей соглашаются с тем, что в поисках корма трогонтерии могли отходить от водоемов относительно дальше, чем современные бобры, но все же считают их околоводными животными. Согласно этой точке зрения, трогонтерии предпочитали водоемы со слабо проточной или стоячей водой: заболоченные берега рек, тростниково-рогозовые озера и ольхово-осоковые болота, а также надречные участки с богатой водной и прибрежной растительностью. Населяя перелески умеренного климатического пояса, трогонтерии избегали как сплошных лесов, так и слишком открытой местности. Данные предположения косвенно подтверждаются некоторыми фактами. Так, в местности Тегелен (Нидерланды), где обнаружены многочисленные окаменелости трогонтерия Кювье, также были найдены обильные остатки водных растений — эвриалы (Euryale sp.), рдеста (Potamogeton sp.), наяса (Najas sp.), водяного ореха (Trapa natans), стрелолиста (Sagittaria sp.), частухи (Alisma sp.), телореза (Stratiotes sp.), водяного лютика (Ranunculus aquatilis).
Возможно, трогонтерии питались растительностью, произраставшей в водоемах и в непосредственной близости от них. Основную часть рациона составляли рогоза, тростник, камыш, кувшинки, корневища и клубни, вырываемые из мягкого прибрежного грунта и т. п. Также высказано предположение, что трогонтерии, в отличие от современных бобров, могли кормиться во время плавания. Хотя трогонтерии имели массивные резцы, они использовали их не для валки деревьев, а ограничивались сгрызанием коры и срезанием молодых побегов. Подвижная верхняя губа могла быть полезной при срывании водных растений. Плотин и хаток, подобно современным бобрам, трогонтерии скорее всего не сооружали, а жили в норах, вырытых по берегам водоемов. Таким образом, экологически эти бобры могли больше всего напоминать современную нутрию (Myocastor coypus).
Расцвет и вымирание
В позднем плиоцене климат на территории Восточной Европы становится более континентальным. В фауне этого времени исчезают гиппарионы, бегемоты и тапиры, не встречаются обезьяны (за исключением Закавказья). Начали преобладать животные открытых пространств — лошади, верблюды, эласмотерии, быки, страусы и др. Господствующее значение получают степи с травянистой растительностью. Лесные массивы сохраняются главным образом в долинах рек, где обитали мастодонты, слоны, олени, носороги. В это время появляется и начинает распространяться трогонтерий Кювье. Судя по находкам, в это время он имел уже довольно обширный ареал — Северный Казахстан, Закавказье, Приазовье, но численность этого животного была, очевидно, небольшой.
В раннем плейстоцене ареал трогонтерия Кювье протягивался уже через всю Евразию. Его численность значительно возросла, судя по многочисленным находкам серий костей х в таких местонахождениях, как Норфолк (Великобритания), Тегелен (Нидерланды), Порт-Катон (Россия). Фауна к раннему плейстоцену несколько изменилась — вымерли мастодонты, саблезубые кошки, страусы, уменьшилось количество верблюдов, значительно возрос процент слонов и оленей. Климат становился влажнее и холоднее. Широколиственные леса начали постепенно вытеснять сухие степи. Влаголюбивая растительность позволила трогонтерию сравнительно легче добывать себе пишу. Возможно, этим можно объяснить расширение его ареала и увеличение численности.
В начале среднего плейстоцена в связи с общим похолоданием и иссушением климата меняется фауна млекопитающих. В ней увеличивается процент быков и лошадей, начинает формироваться новая ветвь слонов, ведущая к настоящему мамонту, почти полностью исчезают верблюды, проявляются лоси и некоторые виды оленей. Численность трогонтерия Кювье на территории Восточной Европы резко падает. В Западной Европе и в Китае, где климат был мягче, этот бобр продолжал существовать. Изменение климата, а следовательно, и растительности, оказало свое влияние на трогонтерия. Размеры его стали больше, предкоренные зубы еще более развились, что, очевидно, было связано с более жесткой пищей и усилением режущей функции зубного аппарата. Конечно, трогонтерий Кювье в определенной мере конкурировали с обыкновенным бобром, с которым нередко соседствовал, но в целом их экологические ниши пересекались не слишком значительно.
В результате дальнейшего общего похолодания и развития на Евразиатском материке покровных оледенений трогонтерий Кювье постепенно начал вымирать и к концу среднего плейстоцена полностью исчез в Западной Европе. На территории Китая популяция этого животного еще существовала в угнетенном состоянии и окончательно исчезла ближе к концу позднего плейстоцена. Не исключено, что определенную роль в этом сыграла охотничья деятельность первобытного человека.
Ссылки
Trogontherium (The Paleobiology Database) перейти
Trogontherium (Wikipedia) перейти
Литература
Агаджанян А. К., Мотузко А. Н. Териофауна плейстоцена. — М.: Издательство Московкого университета, 1972
Основы палеонтологии. В 15 т. Т. 13: Млекопитающие / Гл. ред. Орлов Ю. А. — М.: Государственное научно-техническое издательство литературы по геологии и охране недр, 1962