Улемозавры — Ulemosaurus
Класс †Синапсиды (Synapsida)
Подкласс †Терапсиды (Therapsida)
Надотряд †Динообразные (Dinomorpha)
Отряд †Диноцефалы (Dinocephalia)
Подотряд †Тапиноцефалиды (Tapinocephalida)
Надсемейство †Тапиноцефалоиды (Tapinocephaloidea)
Семейство †Улемозавровые (Ulemosauridae)
Род †Улемозавры (Ulemosaurus)
Вид †Ulemosaurus svijagensis
Вид †Ulemosaurus gigas
Улемозавры (Ulemosaurus Riabinin, 1938) — род монотипического семейства улемозавровых (Ulemosauridae Ivachnenko, 1994), происходящий из средней перми Европы. Установлены 2 вида.
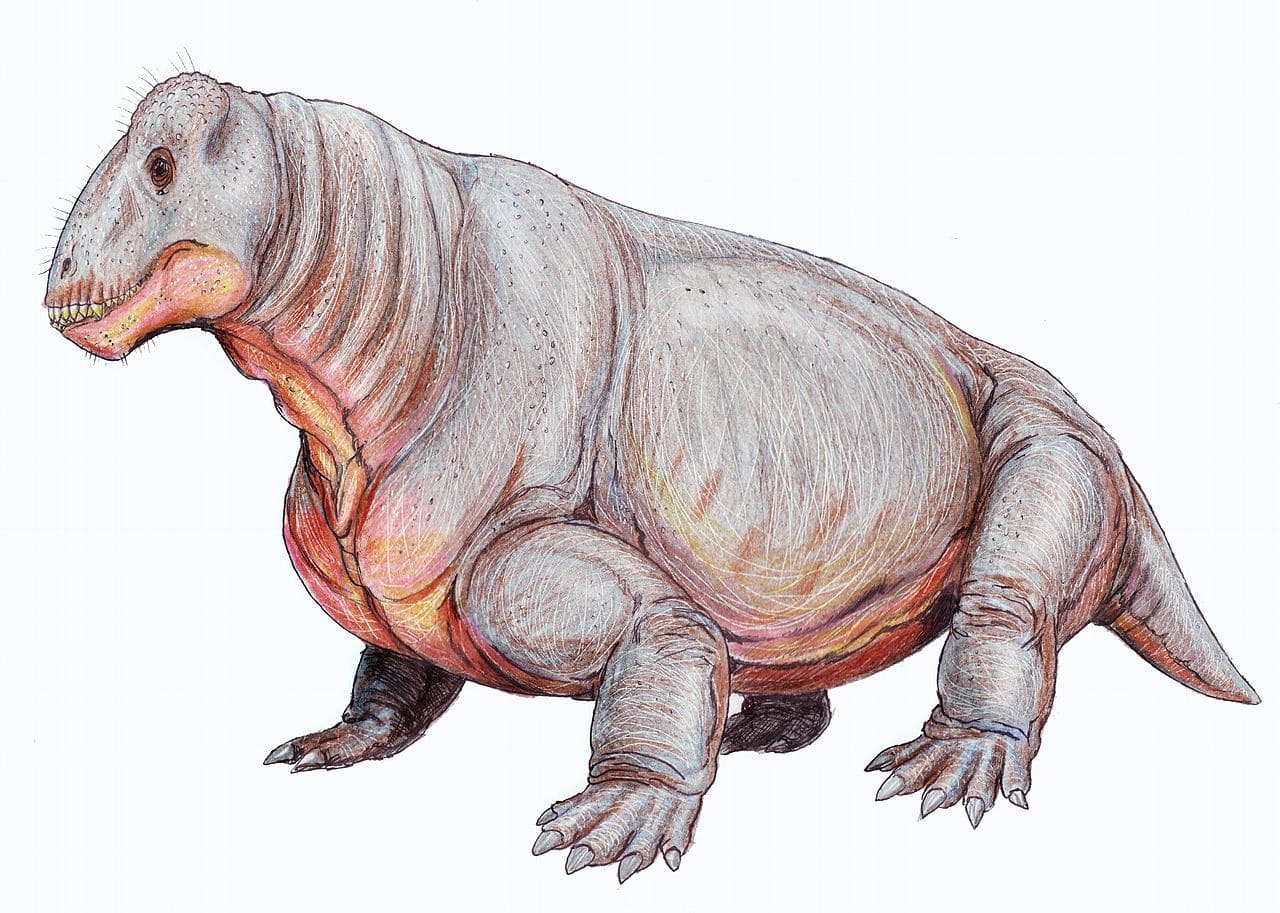
Реконструкция внешнего облика улемозавра (Ulemosaurus svijagensis). Автор — Д. Богданов.
Синонимы: Eurosaurus von Waldheim, 1842 (частью); Deuterosaurus Eichwald, 1846 (частью); Moschops Broom, 1911 (частью); Taurocephalus Broom, 1928 (частью).
Систематика
Изначально Ulemosaurus был описан в качестве самостоятельного рода А.Н. Рябининым в 1938 г. по остаткам, найденным около села Ишеево в Татарстане. Свое название род и типовой вид получил по месту первого обнаружения — бассейну реки Улемы, правого притока Свияги, протекающего в Поволжье. В дальнейшем неоднократно высказывались предположения о близости этого животного к южноафриканским тапиноцефалидам Moschops, вплоть до признания улемозавров видами последних (Л.П. Татаринов (1965 г.), П.К. Чудинов (1983 г.)). Однако, несмотря на то, что эти животные существовали приблизительно в одно время и на первый взгляд весьма сходны между собой, улемозавры демонстрируют немало архаичных черт, не свойственных настоящим Tapinocephalidae. По мнению ряда М.Ф. Ивахненко (1994, 2008 гг.), примитивность морфологии не только подтверждает валидность данного рода, но и дает основания для его выделения в отдельное, архаичное семейство Ulemosauridae.

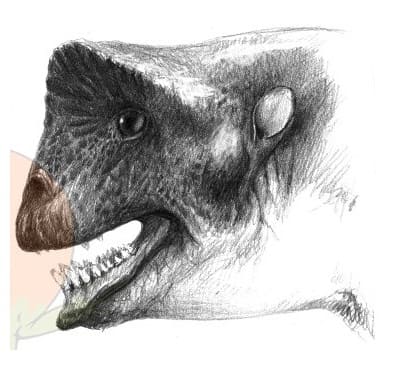
Слева: череп улемозавра (Ulemosaurus svijagensis) из местонахождения Ишеево, Республика Татарстан, Россия.
Справа: портретная реконструкция улемозавра (Ulemosaurus svijagensis). Автор — Ф. Шпиндлер.
Возраст и распространение
Улемозавры являются типичными представителями ишеевского фаунистического комплекса наземных позвоночных. Их остатки вполне обычные в ряде местонахождений Приуралья (Татарстан, Башкирия, Оренбуржье). Они датируются средней пермью (уржумский горизонт, уржумский ярус, биармийский отдел), что приблизительно соответствует времени образования самых нижних горизонтов южноафриканской системы Карру. Можно предположить, что предки улемозавров мигрировали в Приуралье из южных регионов Гондваны.

Череп с нижней челюстью улемозавра (Ulemosaurus svijagensis) из местонахождения Ишеево, Республика Татарстан, Россия, раздавленный весом вышележащих слоев.
Строение и функции
Размеры крупные: длина черепа составляет порядка 40-45 см, общая длина оценивается в 3-4 м.
Массивный череп короткий, высокий и широкий. Его лицевая часть относительно узкая, вытянутая, наклоненная вперед и вниз. Ноздри низко опущены. Глазницы небольшие. Снизу височное окно узкое, но его верхняя часть существенно шире, чем у Moschops. Наблюдается сильный пахиостоз костей в межглазничной и теменной областях. Лобные и все окологлазничные кости бугристы на внешней поверхности и так массивны, что при рассматривании черепа сверху глазные впадины совсем не видны: они прикрыты "нахмуренным" лбом и задними краями глазниц. Верхний край глазницы настолько утолщен, что сбоку кажется лобным рогом. Теменные кости в плане образуют фигуру косого креста. При длине черепа немногим более 40 см их толщина достигает не менее 7 см; очевидно, еще большей была длина нерва пинеального органа.

Череп с нижней челюстью улемозавра (Ulemosaurus svijagensis) из местонахождения Ишеево, Республика Татарстан, Россия.
Затылок очень высокий, наклоненный назад, вогнутый сильнее, чем у Moschops. Околозатылочные отростки очень массивные. Отогнутый назад и вниз затылочный мыщелок, нависший над ним затылок и форма скошенных на переднем и заднем концах шейных позвонков указывают на наклонное положение головы. Затылочная поверхность слабовогнутая, с мощными гребнями; она пропорционально выше, чем у хищных диноцефалов. Межптеригоидная щель снизу сужена вследствие сближения друг с другом крыловидных костей, с дорсальной же стороны она очень длинная, широкая, прикрытая с боков их вертикальными септами.
Верхняя крыловидная кость широким, но слабым основанием лежит на крыловидной. От основания вверх она резко сужается в очень тонкий, узкий и слабый пластинчатый стержень, который непропорционально мал и узок по сравнению с его развитием у хищных диноцефалов (например, у Notosyodon). Имеются не ограниченные крыловидными костями примитивные видиевы каналы. Поперечные отростки крыловидных костей развиты слабее, чем у хищных диноцефалов, не доходят до ветвей нижней челюсти и не ограничивают ее движений. В небной щели в передней части сошника отмечен выход каналов якобсонова органа. Область квадратоартикулярного сочленения слабо продвинута вперед, не далее уровня середины длины височного окна.
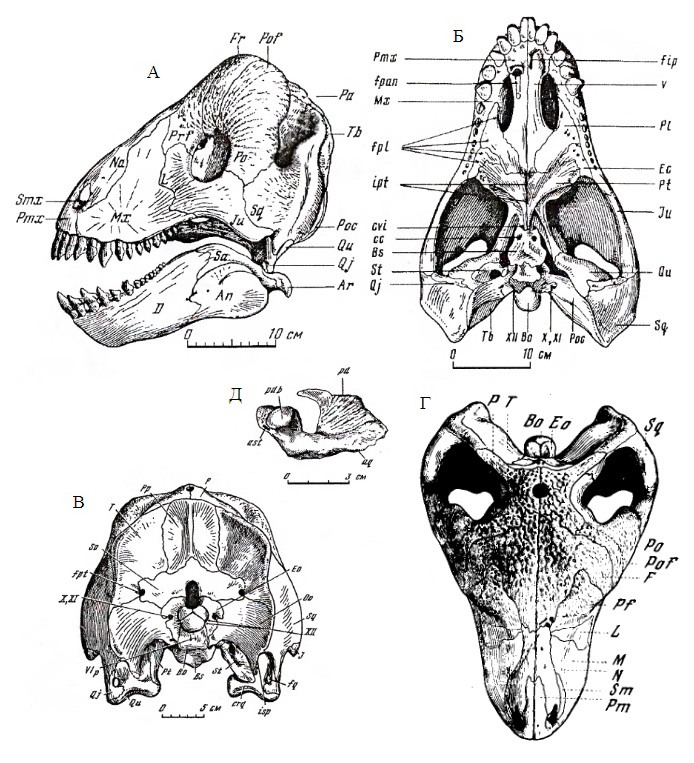
Прорисовки реконструированного черепа с нижней челюстью улемозавра (Ulemosaurus svijagensis) из местонахождения Ишеево, Республика Татарстан, Россия; сбоку (А), снизу (Б), сзади (В), сверху (Г), слуховая косточка (Д).
Челюстной сустав опущен значительно ниже уровня затылочного и допускает некоторую свободу боковых движений нижней челюсти, что обычно характерно для растительноядных позвоночных. Нижняя челюсть относительно укороченная, массивная и высокая, с отогнутой вниз сочленовной частью и очень массивным симфизом. В целом скуловая дуга, морда и нижняя челюсть короче, выше и массивнее, чем у хищных диноцефалов. Небо сходно с их небом, однако лишено зубных бугров на небных костях и не несет зубов на крыловидных. В то же время, следы небных зубов все еще различимы на эктоптеригоидах и прилежащей части птеригоидных флангов.
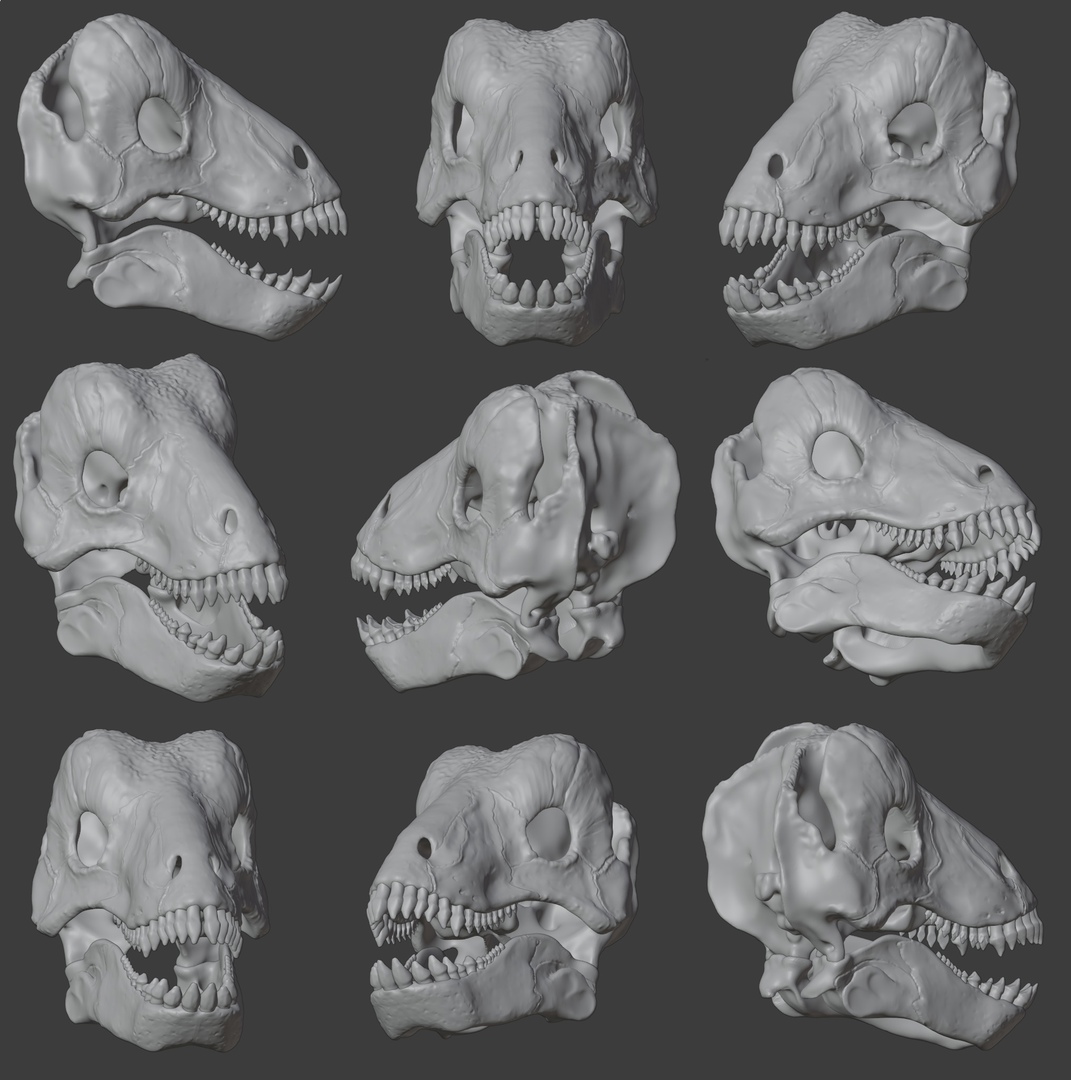
Компьютерные модели черепа с нижней челюстью улемозавра (Ulemosaurus svijagensis). Автор — И. Белоусов.
Строение уха также сходно с таковым у хищных диноцефалов — слуховая косточка, или стремечко, имеет вид распорки в основании черепа между квадратной костью, сочлененной с нижней челюстью, и мозговой коробкой. Своим расширенным концом оно закрывает овальное окно, ведущее в лабиринт внутреннего уха. Стремечко очень массивно, и на первый взгляд, с трудом может быть заподозрено в способности проводить звук, механизм восприятия колебаний которого остается до конца невыясненным.
В верхней челюсти с каждой стороны имеется 14-15, в нижней — 16 зубов, обладающих характерным для растительноядных тапиноцефолоидов строением. Особенно развиты передние резцеобразные зубы, в связи с чем предчелюстные и нижнечелюстные кости спереди очень массивны. Передний край каждого резца высокий и заостренный, а в основании его коронки на его язычной стороне имеется широкий пяточный уступ; эмаль по краям коронок острая, зубчатая. Резцы постепенно уменьшаются в заднем направлении.
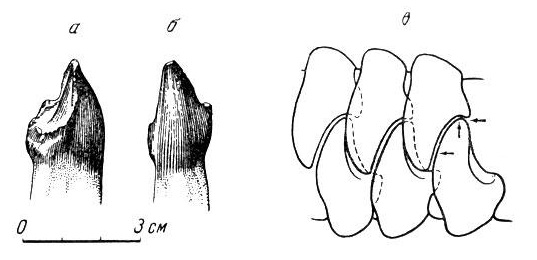
Прорисовки правого нижнечелюстного III-го резца улемозавра (Ulemosaurus svijagensis) из местонахождения Ишеево, Республика Татарстан, Россия; сбоку (а) и спереди (б), а также схема раздавливающей работы резцов (в).
За расположенными на премаксилле резцами, шестым по счету идет I-й зуб верхнечелюстной кости. По диаметру и длине он примерно в 1,5 раза превосходит предыдущий и последующий зубы, обладает режущими краями, но при этом уже почти полностью утратил свою клыкообразность и стал коротко-цилиндрическим, массивным, с выраженным пяткообразным выступом. Морфологически этот клык очень похож на последний предклыковой зуб Deuterosaurus. На нижней челюсти подобного увеличения зубов не наблюдается. Следующие дальше щечные зубы постепенно уменьшаются в размерах, их мелкие лепестковидные коронки также имеют острые передние и задние режущие грани.
При закрытии пасти зубные ряды плотно смыкались, при этом вершины верхних резцов вклинивались в углубления на пятках их нижних антагонистов (и наоборот), одновременно перерезая, раздавливая и перетирая какой-то умеренно плотный корм, очевидно, растительного происхождения. В отличие от большинства растительноядных четвероногих, небольшие щечные зубы не измельчали, а всего лишь удерживали находившиеся во рту пищевые массы. Подобному устройству зубного аппарата соответствовал и характер работы челюстной мускулатуры. Об интенсивности питания свидетельствует значительный износ давящих поверхностей передних зубов.
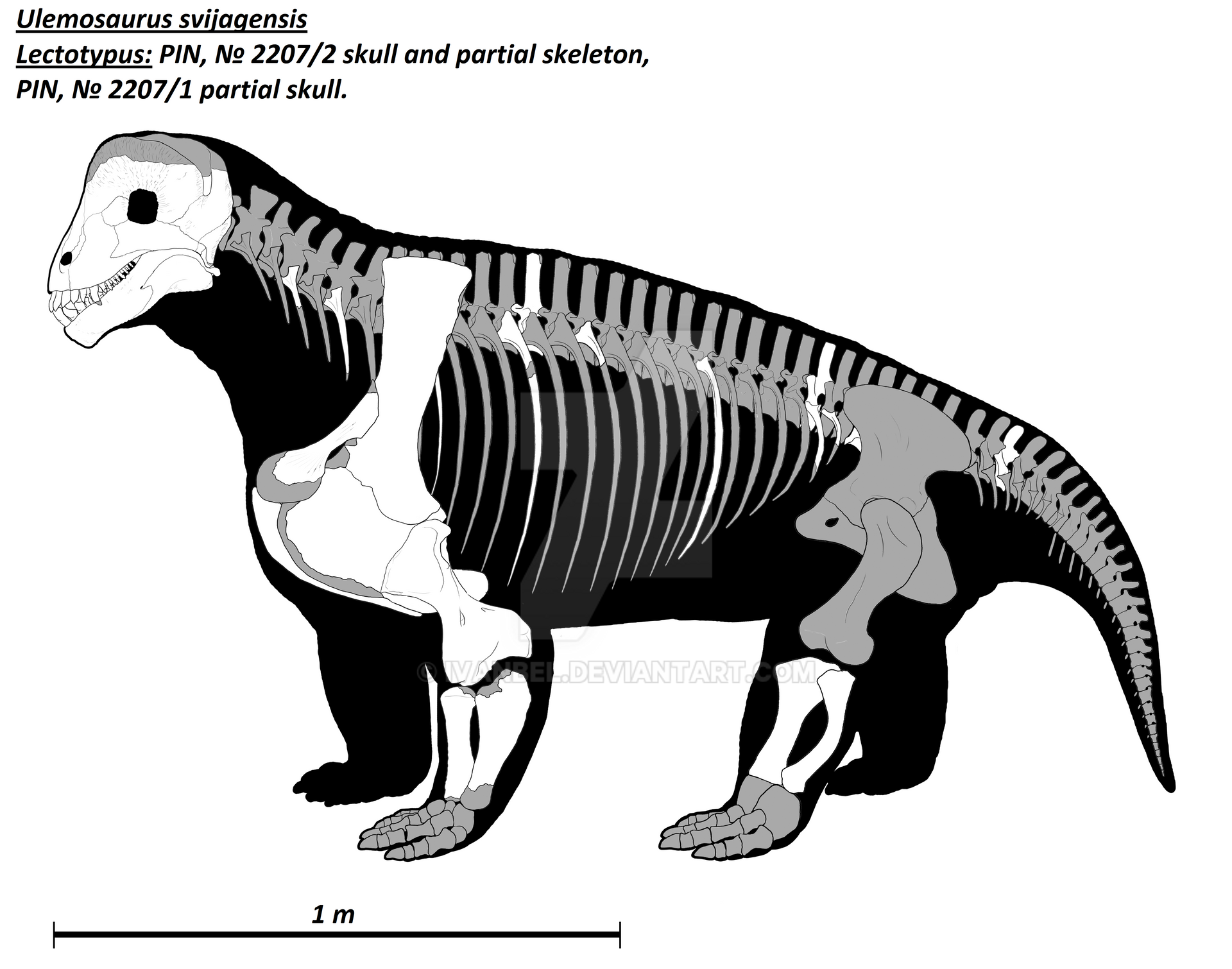
Прорисовка реконструированного скелета улемозавра (Ulemosaurus svijagensis), вписанного в телесные очертания. Автор — И. Белоусов.
Подобно тапиноцефалидам, наклоненная вниз голова находится довольно высоко над землей, а спина полого опускается в заднем направлении. Бочкообразное туловище переходит в короткий хвост. Плечевой пояс массивный. Более длинные и массивные передние конечности расставлены в стороны, в то время как задние в большей мере подведены под туловище.
Сравнение
По сравнению с Moschops у Ulemosaurus относительно длиннее морда; задний отросток септомаксиллярной кости развит меньше, а септомаксиллярный канал имеет иное положение; размеры височных окон несколько больше, а их верхние и дорсальные участки расширены гораздо значительнее; теменная область черепа более сужена; затылок относительно более вогнут; челюстное сочленение менее смещено вперед; альвеолярный край верхнечелюстной кости более выпуклый; межптеригоидная щель имеет большую протяженность; задние участки квадратных ветвей птеригоидов расставлены уже. Примечательно, что в последней особенности Ulemosaurus сходен с хищным диноцефалом Notosyodon, к тому же обладающим сильным и равномерным пахиостозом теменной области.
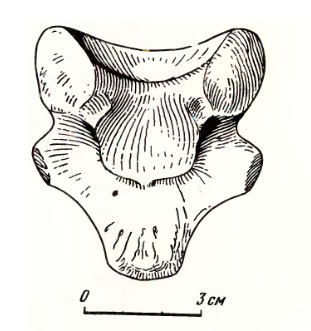
Прорисовка интерцентра атланта улемозавра (Ulemosaurus svijagensis); сверху.
Ulemosaurus превосходил Moschops по величине, при этом его тело не обладало настолько округленными очертаниями. По типу строения посткраниальный скелет животных довольно схож, однако у Ulemosaurus наблюдается больше примитивных черт. Плечевой пояс заметно массивнее, чем у Moschops. Клейтрум, по-видимому, отсутствует. Основные структурные отличия от южноафриканского рода заключаются в большей ширине и мощности заднего отростка межключицы, большей длине и стройности плечевой кости при меньшей ее массивности, а также в сильно выраженном изгибе тела малой берцовой кости. Как и у тапиноцефалид, тела позвонков укороченные.

Череп с нижней челюстью улемозавра (Ulemosaurus svijagensis) из местонахождения Ишеево, Республика Татарстан, Россия.
Состав
Ulemosaurus svijagensis Riabinin, 1938 (тип) — типовой вид, известный из средней перми (уржумский горизонт, уржумский ярус, биармийский отдел) окрестностей села Ишеево (Апастовский район, Республика Татарстан, Россия). Синонимы: Taurocephalus sр., err. determ. Efremov, 1940; Moschops sviyagensis Tatarinov, 1965. Таксон был описан русским палеонтологом А.Н. Рябининым в 1938 г. Он входит в ишеевский подкомплекс ишеевского фаунистического комплекса наземных позвоночных.

Смонтированный неполный скелет улемозавра (Ulemosaurus svijagensis) из местонахождения Ишеево, Республика Татарстан, Россия.
Обнаружены черепа и некоторые части посткраниального скелета (необходимо отметить, что найденные черепа подверглись деформации, вследствие чего их очертания несколько отличаются). В качестве лектотипа принят хорошо сохранившийся череп с нижней челюстью. III-IV-й резцы со слабо и неравномерно зазубренным цингулюмом, умеренно удаленным от вершины и ограничивающим углубленную пятку, круто поднимающуюся к вершине. Поверхность пятки гладкая, с 1-2 продольными варикозными гребешками.



Реконструированный скелет улемозавра (Ulemosaurus svijagensis) из местонахождения Ишеево, Республика Татарстан, Россия.
Ulemosaurus gigas (Efremov, 1954) Ivachnenko et al., 1997 — вид, известный из средней перми (зона Ulemosaurus, уржумский горизонт, уржумский ярус, биармийский отдел) местонахождения Ключевский рудник-2 (Стерлибашевский район, Республика Башкортостан, Россия). Синонимы: Eurosaurus uralensis Eichwald, 1860 (частью); Deuterosaurus gigas Efremov, 1954; Moschops gigas Tchudinov, 1983. Таксон был описан И.А. Ефремовым в 1954 г. Он представляет малокинельский подкомплекс ишеевского фаунистического комплекса наземных позвоночных.
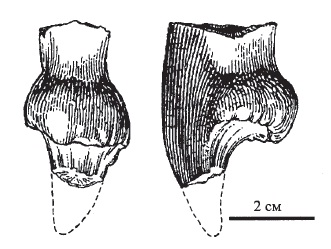
Прорисовки левого I-го верхнечелюстного резца улемозавра (Ulemosaurus gigas) из местонахождения Ключевский рудник-2, Республика Башкортостан, Россия: сзади и сбоку.
Обнаружены разрозненные резцы. В качестве лектотипа принят левый III-й верхнечелюстной резец. Зубы типично тапиноцефаловые по облику и отличаются от резцов Ulemosaurus svijagensis несколько большими размерами и меньшей изношенностью коронок. Коронки сильно расширены поперечно и удлинены в лингвальном направлении. Цингулюм обладает сильно зазубренным кантом, пятка полого поднимается от него к вершине зуба и покрыта многочисленными варикозными гребешками.

Частичный череп с нижней челюстью улемозавра (Ulemosaurus cf. gigas) из местонахождения Озерки, Оренбургская область, Россия.
Остатки, обозначенные Ulemosaurus cf. gigas Efremov, 1954, происходят из средней перми (большекинельская свита, уржумский горизонт, уржумский ярус, биармийский отдел) местонахождения Озерки (Новосергеевский район, Оренбургская область, Россия). Найден неполный череп: предчелюстные кости, части максилл, часть неба, затылка и зубных костей. Череп крупнее и с явно более длинной мордой, чем у Ulemosaurus svijagensis. Сохранился клык, по строению идентичный типовому экземпляру Ulemosaurus gigas.

Прорисовка реконструированного скелета улемозавра (Ulemosaurus gigas). Автор — И. Белоусов
Образ жизни
Предполагается, что улемозавры, подобно более прогрессивным тапиноцефалидам, были преимущественно растительноядами-сапрофагами. Основу их питания могли составлять отмершие растительные массы (например, гниющие стволы членистостебельных каламитов), накапливавшиеся по берегам водоемов. Вполне вероятно, что эти животные немало времени проводили в воде, однако из-за громоздкого телосложения вряд ли хорошо плавали. Вероятно, улемозавры не являлись редкими животными — так, Ulemosaurus svijagensis принадлежит до 20 % остатков, обнаруженных в Ишеевском местонахождении.

Улемозавры (Ulemosaurus svijagensis) в естественном окружении. Автор — Г. Соколов.
Ссылки
Улемозавры (Википедия) перейти
Ulemosaurus (Paleobiology Database) перейти
Литература
Богданов Д. В. Звероящеры и другие пермские монстры. — М.: Фитон XXI, 2018
Ивахненко М. Ф. Тетраподы Восточно-Европейского плаката — позднепалеозойского территориально-природного комплекса. — Пермь, 2001
Ивахненко М. Ф., Голубев В. К., Губин Ю. М., Каландадзе Н. Н., Новиков И. В., Сенников А. Г., Раутиан А. С. Пермские и триасовые тетраподы Восточной Европы. — М.: ГЕОС, 1997
Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Ч. 1 / Отв. ред. Ивахненко М. Ф. и Курочкин Е. Н. — М.: ГЕОС, 2008
Орлов Ю. А. В мире древних животных. Очерки по палеонтологии позвоночных. — М.: Наука, 1989
Основы палеонтологии. В 15 т. Т. 12: Земноводные, пресмыкающиеся, птицы / Гл. ред. Орлов Ю. А. — М.: Наука, 1964
Ромер А. Ш. Палеонтология позвоночных. — М. — Л.: Государственное научно-техническое издательство нефтяной и горно-топливной литературы, 1939
Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М.: ГЕОС, 2009
Чудинов П. К. Ранние терапсиды. — М.: Наука, 1983