в оглавление
назад
Энергообмен
Чтобы выжить, животные должны уравновешивать поступление энергии и ее расходы. Энергия поступает в организм с питательными веществами и извлекается путем их окисления. Извлеченная таким образом энергия расходуется на все процессы и акты жизнедеятельности. Интенсивность, с которой в организме происходят химические процессы и соответственно должна поступать извне энергия, называется уровнем обмена веществ. Температура тела зависит от количества тепла, образующегося за счет метаболических процессов, протекающих в организме, и от количества тепла, потерянного (или полученного) животным.
Рыбы, амфибии и рептилии в большинстве своем пойкилотермны, или холоднокровны, т. е. температура их тела переменна, изменяясь в зависимости от температуры окружающей среды. Хотя некоторые из них и могут влиять на температуру тела путем увеличения общих размеров и при помощи ряда внутренних механизмов, способности эти весьма ограничены. Непостоянству температуры тела соответствует и невысокий уровень метаболизма низших позвоночных. На выход температуры внешней среды за пределы предпочтительного диапазона (оптимума) пойкилотермные животные реагируют вхождением в состояние анабиоза, и за счет снижения энергозатрат переживают температурный стресс.
Для птиц и подавляющего большинства млекопитающих характерна гомойотермия, или теплокровность, — способность сохранять относительно постоянную температуру тела, несмотря на флуктуации температуры окружающей среды. Это достигается за счет освобождения большого количества энергии, что в свою очередь становится возможным благодаря значительному повышению уровня обмена веществ, в 20-30 раз превосходящего таковой у низших позвоночных. Метаболизм возрастает вследствие прогресса всей телесной организации: дыхательной системы, доставляющей организму много кислорода; сердечно-сосудистой системы, в которой совершилось полное отделение артериального тока крови от венозного, и поэтому ткани снабжаются только кровью, богатой кислородом; мышечной системы, в результате работы которой освобождается много тепла; пищеварительной системы, быстро и эффективно перерабатывающей различные пищевые продукты, которые затем ассимилируются в организме и восполняют расход веществ и энергии, образующейся в результате диссимиляции; выделительной системы, быстро удаляющей из организма продукты диссимиляции, накопление которых тормозило бы обмен веществ; нервной системы, которая координирует работу всех систем и органов и оказывает воздействие на характер и интенсивность обмена веществ.
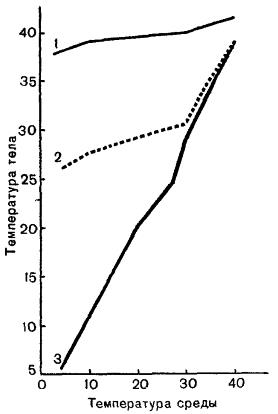
Кривые зависимости температуры тела различных животных от температуры внешней среды: 1 — кролик (Oryctolagus cuniculus); 2 — ехидна (Tachyglossus aculeatus); 3 — ящерица (Lacerta agilis).
Средняя температура у однопроходных составляет 30-33 °С, у сумчатых — 34-37 °С, у плацентарных — 36-40 °С. По-видимому, определенную роль играют климатические условия: средняя температура тела тропических млекопитающих (37,8 °С) несколько ниже, чем арктических (38,3 °С). Температура у большинства видов относительно постоянна, диапазон ее суточных колебаний составляет всего 0,5-2 °C. При этом разница температуры тела от температуры внешней среды может быть очень значительной: например, у зайца-беляка (Lepus timidus) она достигает 68 °С, а у полярного волка (Canis lupus) и песца (Vulpes lagopus) — 74 °С. (Для сравнения — у низших позвоночных разница между температурой тела и окружающей среды редко превышает 1-2 °С.) У птиц средняя температура тела составляет около 42 °С, а его верхняя граница доходит до 45,5 °С. Таким образом, уровень их метаболизма еще выше, чем у млекопитающих.
Теплокровность открыла для освоения перед своими обладателями огромные жизненные пространства умеренных и полярных широт и высокогорий, не доступных большинству видов пойкилотермных животных. Полярные окраины материков, острова Заполярья и даже плавающие льдины служат ареной активной жизни для птиц и млекопитающих. В умеренных зонах в обоих полушариях стоят снежные и холодные зимы, и в этот суровый сезон здесь буквально царят теплокровные, тогда как холоднокровные переживают период низких температур, находясь в неактивном состоянии. Именно поэтому в составе фауны областей с прохладным климатом птицы и млекопитающие составляют относительно более высокий процент по числу видов, чем в тропиках.

Однако гомойотермия не только дает возможность функционировать в неблагоприятной среде, она также обеспечивает биохимическую стабильность внутри самого организма (что особенно важно для процессов, протекающих в головном мозге). Ферментные системы теплокровных животных работают очень эффективно, но сохраняют свою активность лишь в строго определенном диапазоне температур с оптимумом, близким к физиологической температуре тела. Поэтому для большинства теплокровных животных зоны умеренного климата температуры тела свыше 40 °С губительны. Именно с этого уровня температур начинается процесс денатурации белков, в который раньше других вовлекаются белки со свойствами катализаторов, т. е. ферменты. По отношению к понижению температур эти вещества более терпимы. После охлаждения до 4 °С и последующего восстановления температурных условий ферменты восстанавливают свою активность.
Отрицательные температуры губительны для теплокровного организма по другой причине. Как известно, основной составной частью организма млекопитающих (порядка 60-80 % от живой массы) является вода. Поскольку протоплазма клеток представляет собой водную фазу, то при отрицательных температурах вода из жидкого состояния переходит в твердое. Образование кристаллов воды в составе протоплазмы клеток и в межклеточной жидкости оказывает повреждающее воздействие на клеточные и субклеточные мембраны.
В связи с тем, что температура тела у птиц и млекопитающих почти всегда отличается от температуры окружающей среды, даже в неактивном состоянии их физиологические системы должны работать, чтобы обеспечить постоянство внутренней температуры и воспрепятствовать избыточному оттоку или притоку тепла. Например, во время отдыха 80-90 % энергии, «сжигаемой» гомойотермным организмом, используется исключительно для поддержания постоянной температуры тела. Кроме того, при переходе от лета к зиме организму требуется больше энергии, поскольку увеличиваются его теплопотери. Соответственно, теплокровные животные вынуждены вести активный образ жизни, т. е. потреблять достаточно много пищи и быстро ее усваивать, чему в свою очередь способствует высокая температура тела. Таким образом, недостаток гомойотермии в том, что она неизбежно диктует необходимость высоких энергозатрат.
Температура тела и интенсивность обмена веществ некоторых групп позвоночных (по Наумову, 1989)

Постоянство температуры тела является результатом уравновешенной теплопродукции и теплоотдачи. В ходе своей эволюции млекопитающие выработали разнообразные средства нервной и гуморальной регуляции температуры, которые могут затрагивать метаболизм, энергетический обмен и поведение животного. При этом активация тех или иных механизмов зависит от времени суток, сезона, пола и возраста. Главным источником тепла являются многочисленные физиолого-биохимические процессы, интенсивность которых регулируется рефлекторно.
Энергопродукция обеспечивает основной обмен (работоспособность всех физиологических систем в состоянии физиологического покоя) и продуктивный обмен (работу скелетных мышц, рост плода, лактопоэз). Основными тепловыми генераторами организма выступают мышцы (до 50 % всей теплопродукции организма), печень (15-20 % тепла), легкие и почки (7-12 % тепла), а также желудочно-кишечный тракт (10% тепла). У жвачных заметная часть теплопродукции принадлежит симбиотическим микроорганизмам преджелудков и толстого отдела кишечника. Инфузории, бактерии и грибы, населяющие эти отделы пищеварительного тракта, гидролизуют до 80 % клетчатки, 70 % протеина и 60 % липидов рациона.
У пойкилотермных животных внутреннее производство тепла, как правило, превышает собственные потребности. Поэтому в естественной среде обитания значительная часть метаболического тепла выделяется во внешнюю среду. Даже в нормальных температурных условиях пойкилотермные организмы гораздо больше рискуют перегреться, чем переохладиться. При понижении температуры среды обмен веществ хладнокровных животных понижается без отрицательных последствий. При падении температуры воздуха до критической величины животные впадают в спячку.
У теплокровных животных ответ на понижение температуры среды иной. Они повышают обмен веществ и, следовательно, теплопродукцию. Традиционно принято считать, что регулятором этого жизненно важного механизма выступает нервный центр, расположенный в гипоталамической области промежуточного мозга и чувствительный к температурным изменениям.
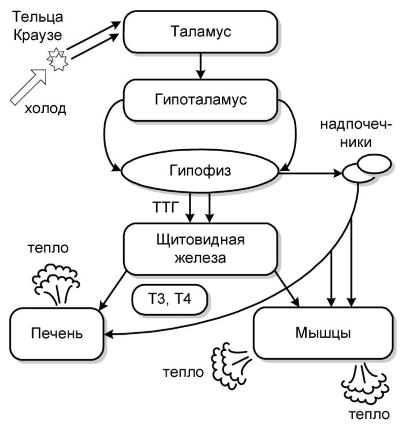
Схема регуляции термического гомеостаза у теплокровных животных при воздействии холода.
Афферентный поток, возникающий в результате возбуждения холодовых рецепторов (телец Краузе), через таламус и гипоталамус активизирует продукцию аденокортикотропного гормона и тиреотропного гормона гипофизом. Под влиянием аденокортикотропного гормона надпочечники выделяют в кровь катехоламины, а щитовидная железа секретирует тиреоидные гормоны. Адреналин и тироксин в печени и мышцах усиливает термогенез за счет окисления аденозинтрифосфорной кислоты. В результате выделяется дополнительное количество тепла, которое согревает тело животного.
Кроме того, под влиянием адреналина активизируется деятельность сердечной мышцы. В результате усиления кровообращения к поверхности тела в единицу времени поступает больше крови и выносится дополнительное тепло, которое повышает температуру кожи и тормозит образование рецепторного потенциала в тельцах Краузе. Афферентный поток с холодовых рецепторов ослабевает, стимулирующее влияние таламуса прекращается. Однако основной обмен обладает некоторой инертностью. Поэтому теплопродукция некоторое время после прекращения действия холодового фактора остается повышенной.
Согласно более современным представлениям, концепция единого управляющего центра отвергается и считается, что температура тела регулируется различными термоэффекторными путями, каждый из которых имеет собственные афферентные и эфферентные ветви. Когда внутренняя температура становится слишком высокой, активируются механизмы охлаждения, а если она падает слишком низко, теплопотери уменьшаются и вступают в действие механизмы согревания. Этот принцип обратной связи такой же, как у термостатически регулируемого электрического обогревателя.
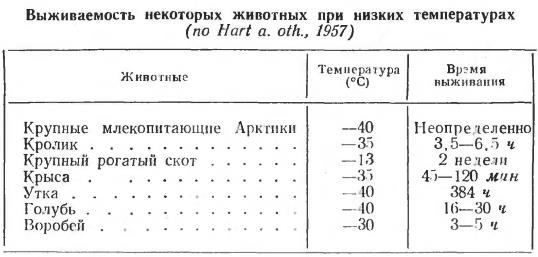
Выживаемость животных при низких температурах среды определяется двумя основными факторами: теплоизолирующими свойствами покровных тканей и способностью животных повышать обмен веществ при охлаждении. Последнее свойство базируется на вегетативных реакциях организма и хорошо развито у полярных животных. Так, у белого медведя (Ursus maritimus) основной обмен повышается при температуре воздуха -50 °С, у песца (Vulpes lagopus) — при -40 °С, у грызунов — при 15 °С. Критически опасной температурой даже для полярных животных считается температура ниже -50 °С, хотя отдельные представители, например эскимосская лайка или песец, поддерживают температуру тела на уровне 38-40 °С даже при температуре воздуха около -80 °С.
Еще более живуча снежная коза (Oreamnos americanus), обитающая в горах Аляски. У нее обнаружен, вероятно, самый совершенный механизм поддержания температуры тела и сохранения жизнеспособности в условиях предельно низких температур среды. Обмен веществ снежной козы остается неизменным в широком диапазоне внешних температур: от +20 °С до -20 °С. Лишь при -30 °С у этого животного удалось зарегистрировать повышение обмена веществ. В 50-градусный мороз снежная коза увеличивает потребление кислорода на 30 %, что достаточно для активного образа жизни. Для сравнения отметим, что у ежа (Erinaceus europaeus) при понижении температуры среды осенью до 5-6 °С обмен веществ возрастает в 3-5 раз по сравнению с летними условиями.
Следует, однако, иметь в виду, что выраженная гомойотермия не является признаком всех без исключения млекопитающих. В полной мере она характерна только для относительно крупных плацентарных. У низших форм, терморегуляционный механизм которых не столь совершенен, а также у мелких плацентарных, имеющих невыгодное для сохранения тепла соотношение между объемом тела и его поверхностью, температура весьма изменчива и во многом зависит от температуры внешней среды. Например, у австралийской ехидны (Tachyglossus aculeatus) она меняется в пределах 22-37 °С, у большой сумчатой крысы (Phascogale tapoatafa) — 29-38 °С, у тенреков (Tenrecidae) — 13-34 °С, у двупалого ленивца (Choloepus didactylus) — 24-33 °C, у одного вида броненосцев — 27-40 °С, у обыкновенной полевки (Microtus arvalis) — 32-37°С. Двугорбый верблюд (Camelus bactrianus) в состоянии покоя имеет довольно низкую ректальную температуру — около 33 °С. Однако в экстремальных условиях (физическая работа на фоне температуры среды свыше 45 °С) температура тела его тела поднимается до 40 °С, т. е. на 7 °С, без заметного влияния на его физиологическое состояние и поведение. Еще значительнее — от 36 °C ночью до 46,5 °C в жаркий день — может подниматься температура тела аравийского орикса (Oryx leucoryx).
В период зимней спячки температура некоторых млекопитающих может опускаться до минусовых значений. Рекордные для гомойотермных животных температурные колебания зафиксированы у гладконосых летучих мышей (Vespertilionidae): в состоянии бодрствования температура их тела достигает 48,5 °C, а во время спячки иногда снижается до -7,5 °C. Обращает на себя внимание то, что относительно широкий диапазон колебаний температуры тела свойствен по большей части животным, обитающим в теплом климате. У северных животных гомойотермия имеет более жесткий характер.
Обитатели пустынь, полупустынь и степей имеют пониженный основной и продуктивный обмен. Это обстоятельство крайне важно для понимания особенностей их поведения. Низкий уровень обмена предполагает пониженную теплопродукцию. Количественные измерения выявили двукратную разницу в интенсивности основного обмена у большой песчанки (Rhombomys opimus) и грызунов, не адаптированных к жизни в аридной зоне — серой крысы (Rattus norvegicus), гребенщиковой песчанки (Meriones tamariscinus). Кроме того, движения животных аридных зон медлительны и рациональны, что также способствует сохранению теплового баланса.
Очень малоинтенсивен для их размеров обмен веществ у роющих подземных грызунов. Основная причина этого — трудности с рассеиванием тепла в душных норах, где пот не может испаряться. Совсем не обладает гомойотермией голый землекоп (Heterocephalus glaber). Замедленность метаболизма, который вдвое ниже, чем у других грызунов, позволяет землекопу выживать в подземных туннелях при очень низком уровне кислорода и высоком — двуокиси углерода. Терморегуляция у многих млекопитающих устанавливается уже в первые после рождения дни. У других (в частности, у мелких землероек и грызунов) — значительно позже, так что их детеныши на какое-то время оказываются по сути пойкилотермными; они согреваются родителями или же укрываются от холода в гнездах или норах. У зверей с хорошо развитой терморегуляцией молодые особи имеют более высокую температуру, чем взрослые.
Потеря тепла у млекопитающих происходит преимущественно (например, у человека — до 70 %) через кожу, которая, таким образом, играет в терморегуляции важнейшую роль. Непосредственно теплообмен с окружающей средой осуществляется развитой системой кожных кровеносных сосудов. Под контролем вегетативной нервной системы содержащиеся в дерме артерии могут расширяться, наполняясь кровью; кожа при этом краснеет, и интенсивно отдает тепло. Если же артериолы сужаются (кожа при этом бледнеет), то теплоотдача резко снижается.
У привычных к холоду животных при резком охлаждении подкожные сосуды сужаются. Южане в этой ситуации демонстрируют прямо противоположную реакцию. У них имеет место расширение сосудов, т. е. усиление кровообращения в периферических сосудах. В связи с этим уместно будет упоминание о «моржах» — людях, которые регулярно купаются зимой в открытых водоемах. Когда в сильный мороз «моржи» вылезают из проруби, их тело имеет красный цвет и от них идет пар. Это говорит о том, что температура поверхности тела человека в этих экстремальных условиях намного превышает температуру окружающей среды.
В наиболее легко реализуемой схеме теплообмена внутреннее тепло просто рассеивается через разветвленную капиллярную сеть в окружающую среду, а охлажденная кровь возвращается по венам. Однако большинство млекопитающих имеет большее сложную систему, в которой теплопотери могут быть минимизированы за счет направления теплой крови через поперечные кровеносные сосуды — анастомозы. Такие артериально-венозные теплообменники описаны в конечностях собак, в плавниках китов, в коже крупного рогатого скота, в хвосте грызунов.
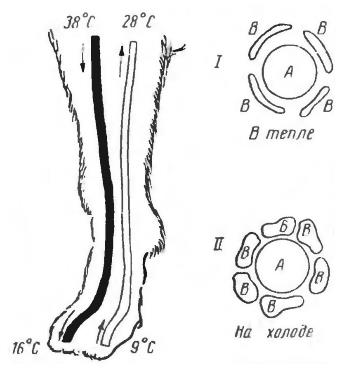
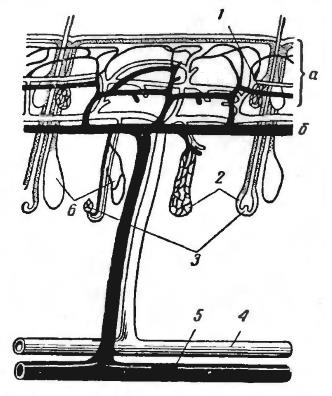
Слева — схема артериально-венозного теплообменника в конечности собаки (Canis lupus familiaris). Поперечный разрез через артерию (А) и оплетающие ее венозные пазухи (В): I — в тепле; II — на холоде.
Справа — кровоснабжение кожи коровы (Bos primigenius taurus): а — поверхностные сосудистые анастомозы; б — глубокие сосудистые анастомозы; 1 — сальная железа; 2 — волосяной фолликул; 3 — волосяная луковица; 4 — вена; 5 — артерия; 6 — потовые железы.
Параллельное и близкое расположение артерий и вен позволяет эффективно выводить из организма избыточное тепло. Артериальная кровь имеет температуру, близкую к физиологической температуре тела. В противотоке часть тепловой энергии от нее забирает венозная кровь. Вены располагаются близко к поверхности, зачастую прямо под кожей, разделяясь и окружая артерии. Следовательно, за счет повышения температуры венозной крови происходит некоторый подогрев поверхности конечности, хвоста или другой части тела. Для животных в условиях низких температур это перераспределение тепла имеет большое значение. За счет противоточного механизма конечности защищаются от обморожения и остаются в рабочем состоянии в экстремальных температурных условиях среды.
У крупного рогатого скота и близких к нему видов противоточный механизм кровообращения имеется в межреберных мышцах. Артерии этих мышц выходят к поверхности тела на спине и боках, где ветвятся и образуют с венами т. н. «чудесную сеть» (rete mirabile). При тепловой одышке (см. ниже) межреберные мышцы отдают тепло прилегающим кровеносным сосудам. В силу высокой теплоемкости крови мышцы эффективно охлаждаются. Температура поверхности тела повышается, следствием чего выступает рассеивание этого тепла во внешнюю среду.
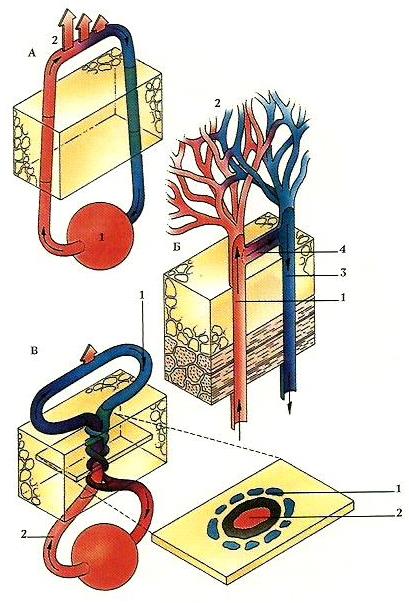
Различные схемы отдачи тепла через кожные кровеносные сосуды:
А — наиболее простая система: рассеивание кровью тепла через артериолы (1) и возвращение охлажденной крови через венулы (2);
Б — более сложная система: поступление теплой крови из артериол (1) в капиллярные сети (2) и ее возвращение через венулы (3), поперечные кровеносные сосуды (4) при необходимости минимизируют отдачу тепла;
С — противоточная система: охлажденная кровь возвращается по венулам (1), которые ветвятся и окружают артериолы (2), подогревающие кровь перед ее возвращением вглубь организма.
Что касается сердца, то его реакции на охлаждение у разных видов неодинаковы. У адаптированных к холоду животных фиксируют снижение частоты пульса. Но у животных, которые не адаптированы к воздействию холода, отмечают прямо противоположный ответ сердца — тахикардию. Например, охлаждение до -4 °С у лабораторных крыс только хвоста вызывает повышение частоты сердечных сокращений на 50-100 %.
От излишних потерь тепла млекопитающих дополнительно предохраняет шерстный покров. Густота и теплоизоляционные свойства шерсти непосредственно связаны с условиями среды. У бореальных млекопитающих она развита особенно хорошо и имеет подшерсток. В преддверие холодного сезона шерстный покров становится еще гуще и длиннее. Кроме того, эти животные влияют на теплоизоляционные свойства шерстного покрова, взъерошивая или укладывая его, чем регулируется количество удерживаемого в нем воздуха. Сама кожа северных зверей сравнительно толстая, что также способствует снижению теплопроводности поверхности тела.
Напротив, у многих тропических млекопитающих шерсть в большей или меньшей степени редуцируется, поскольку не так уж необходима для теплоизоляции. Разница в особенностях шерстного покрова бывает хорошо выражена даже в пределах одного вида. Так, амурскому тигру (Panthera tigris altaica) приходится бродить в глубоком снегу, и все его тело покрыто длинной и пушистой шерстью, которая особенно длинна на загривке и на груди. А суматранский тигр (Panthera tigris sumatrae) имеет короткую и гладкую шерсть, почти совсем без подпушки. Известно, что даже на стоимости мехов (например, лисицы (Vulpes vulpes) и полосатого скунса (Mephitis mephitis)) сказывается то, из какого района они происходят: шкурка тем дороже, чем севернее она добыта. Подобно редкой шерсти, относительно тонкая кожа животных жаркого климата имеет низкие теплоизолирующие свойства.
Удерживать тепло ряду северных видов помогает и развивающийся на зиму подкожный слой жировой ткани; подобная прослойка имеется и у многих водных форм. Помимо выполнения теплоизоляционной функции, жировая прослойка служит источником энергии в период, когда корм недоступен по причине тяжелого ненастья. Установлено, что в результате адаптации к низким температурам у животных меняются свойства жира подкожных отложений и костного мозга. У арктических животных костный жир из пальцевых фаланг имеет низкую точку плавления и не застывает даже в лютые морозы. Однако костный жир из костей, которые не контактируют с холодной поверхностью, например из бедренной кости, имеет обычные физико-химические свойства. Жидкий жир в костях нижней части конечностей обеспечивает теплоизоляцию и подвижность суставов.
Жир накапливают и животные, обитающие в жарком климате. Однако качество, количество и распределение жира по телу у северных и южных животных разное. У арктических млекопитающих жир распределяется в подкожной клетчатке равномерно по всему телу. У животного при этом образуется своеобразная теплоизолирующая капсула. У зверей умеренного пояса жир как теплоизолятор накапливается лишь у видов со слабо развитым шерстным покровом. В большинстве случаев накопленный жир служит им источником энергии в голодный зимний (или летний) период.
В жарком климате подкожные жироотложения несут другую физиологическую нагрузку. Распределение жировых отложений по телу животных характеризуется большой неравномерностью. Жир локализуется в верхней и задней частях тела. Например, у копытных африканских саванн жировая подкожная прослойка локализуется вдоль позвоночника. Она защищает животное от палящего солнца. Брюхо при этом абсолютно свободно от жира. Это также имеет большой смысл. Более холодная по сравнению с воздухом земля, трава или вода обеспечивает эффективный отвод тепла через брюшную стенку в отсутствие жировой прослойки. Небольшие жировые отложения и у животных в жарком климате являются источником энергии на период засухи и связанного с ней голодного существования травоядных.

Лось (Alces alces) неспособен к охлаждению путем потоотделения, вследствие чего ограничен в своем распространении территориями, где летние температуры не превышают 27 °С.
Наличие в носовой полости млекопитающих дыхательных носовых раковин вынуждает воздух при дыхании проходить по узким носовым ходам между этими раковинами, уменьшая потери тепла и влаги. Среди современных позвоночных такие раковины, помимо млекопитающих, имеют только птицы, так что наличие дыхательных раковин считается остеологическим коррелятом теплокровности. Важнейшим доводом в пользу того, что гомойотермными были уже ближайшие предки млекопитающих, стало обнаружение выступов на внутренней поверхности носовых и верхнечелюстных костей тероцефалов и цинодонтов — свидетельство наличия дыхательных раковин и у этих продвинутых зверообразных.
У птиц и млекопитающих имеется такое эффективное средство разогрева организма, как холодовая мышечная дрожь. Понижение температуры среды стимулирует у них теплопродукцию за счет расщепления аденозинтрифосфата в некоторых скелетных мышцах, имеющих для локомоций второстепенное значение. Внешним проявлением этого процесса служит появление тремора — видимой дрожи.
Методом миоэлектрографии показано участие в холодовой дрожи мышц спины, шеи, передних конечностей, жевательных мышц. Мышцы задних конечностей вовлекаются в процесс биохимической терморегуляции в последнюю очередь. Это логично: они больше других ответственны за движения, лежащие в основе поведенческих актов. Однако у водных млекопитающих (нутрия (Myocastor coypus), ондатра (Ondatra zibethicus)) холодовая дрожь ярко проявляется именно у мышц конечностей и мышц спины. У грызунов холодовая дрожь выражена слабее, чем у других животных. При охлаждении у них явно и раньше, чем в других мышцах, развивается тремор жевательных мышц. Мышцы конечностей и туловища включаются в дрожь только при очень сильном охлаждении. Но уровень изученности этого явления выше всего именно у грызунов.

Кашалот (Physeter macrocephalus) способен регулировать свою температуру. Например, при долгой погоне за добычей температура его тела повышается. Избыточное тепло выделяется в окружающую среду. Когда кашалот плавает, то набирает воду через дыхало (1) и пропускает ее через носовые проходы (2). Они соединены с передненосовыми мешками (3). Правый носовой проход проходит через спермацетовый орган (4), оплетенный кровеносными сосудами. Вода при прохождении через носовые проходы и спермацет нагревается. Затем вода выбрасывается через дыхало. Вместе с водой из организма выводится избыточное тепло. Челюстноносовые мышцы (5) контролируют поток воды, образуя подобие насоса. Когда они сокращаются, уплощенные носовые проходы расширяются (6) и наполняются водой (7). Когда мышцы расслабляются, то проходы принимают исходную форму (8) и вода выходит (9).
Многие виды, которые выбирают тактику уклонения от контактов с низкими температурами (мигрирующие, зимнеспящие животные), не обладают столь эффективным приемом термического гомеостатирования, как виды, сохраняющие высокую активность при низких температурах среды. Тропические виды животных вообще не имеют холодовой мышечной дрожи. Поэтому экспериментальное понижение температуры среды обитания у животных тропического пояса приводит к снижению температуры тела и гибели животных от переохлаждения. Животные, обладающие холодовой мышечной дрожью, потенциально более адаптивны к низким температурам. Они способны находить свободные экологические ниши в биотопах высоких широт.
Помимо удержания в организме тепла не менее важно отведение его излишков. Большие размеры в этом случае оказываются менее эффективными, чем мелкие: так, если у бурозубки (Sorex araneus) теплопродукция составляет 151,2 Дж/г в час, то у кошки (Felis silvestris catus) — 12,51, у собаки (Canis lupus familiaris) — 8,34, у слона (Loxodonta africana) — 2,08. У таких крупных обитателей теплых стран, как слоны (Elephantidеa) и носороги (Rhinocerotidae), шерсть почти полностью исчезает, иначе бы они перегревались из-за невыгодного соотношения поверхности тела к его объему. Также теряют шерсть китообразные и сирены, полностью перешедшие к водному образу жизни.
У некоторых китообразных имеется сложная система кровеносных сосудов, переносящих горячую кровь к спинному плавнику, откуда избыточное тепло выводится через кожу. У гренландского кита (Balaena mysticetus) спинной плавник отсутствует, но имеется верхнечелюстное пещеристое тело (corpus cavernosum maxillaris), состоящее из губчатой ткани и способное наливаться кровью, подобно пещеристым телам полового члена. Когда кит раскрывает рот, в него вливается холодная вода, которая забирает тепло из этого органа, охлаждая кровь. Характерно, что чувствительный теплоотводящий орган расположен непосредственно под головным мозгом, касаясь своей верхней частью его основания.
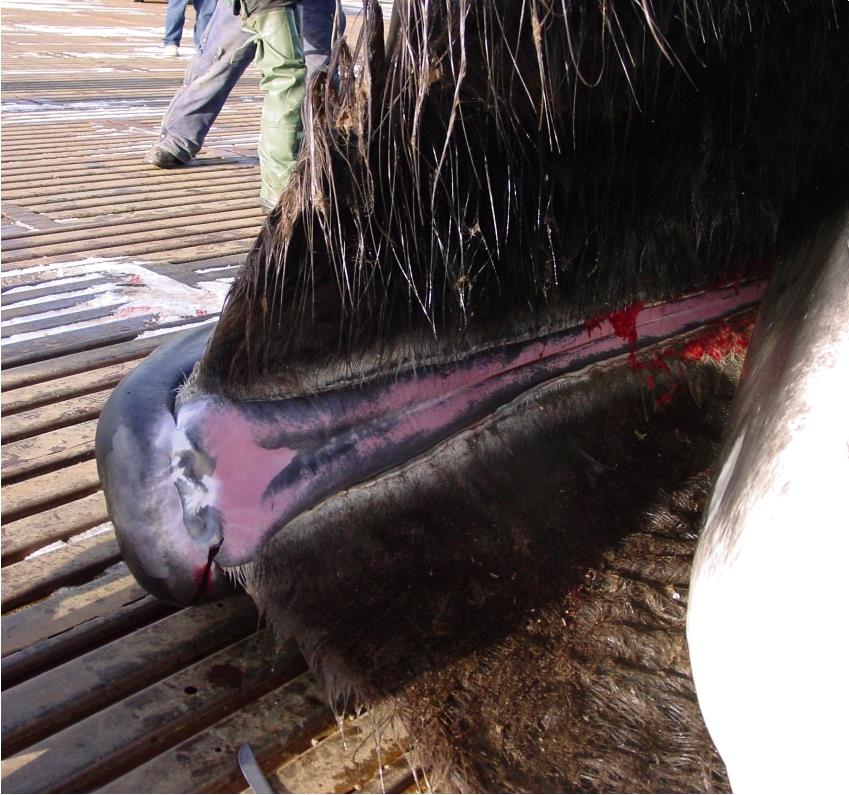
Верхнечелюстное пещеристое тело гренландского кита (Balaena mysticetus).
вперед
в оглавление