в оглавление
назад
Эндокринная система
Выше рассматривалась нервная система — чрезвычайно сложная, но очень эффективно координирующая процессы жизнедеятельности при помощи четких электрохимических сигналов, быстро получаемых ею от конкретных частей тела и посылаемых этим частям ответных сигналов. Во второй интегрирующей системе — эндокринной — информация и команды передаются тканям-мишеням с током крови с помощью химических агентов — гормонов, представленных различными классами органических соединений. Среди них имеются пептиды (окситоцин, вазопрессин, адренокортикотропный гормон, паратгормон, кальцитонин), белки (гормон роста, пролактин, инсулин, глюкагон), гликопротеины (фолликулостимулирующий гормон, лютеинизирующий гормон, тиреотропный гормон), производные аминокислот (трийодтиронин, тироксин, адреналин, норадреналин, мелатонин) и стероиды (глюкокортикоиды, минералокортикоиды, андрогены, эстрогены, прогестерон). Каждый из них обладает своими особыми функциями.
Гормоны вырабатываются железами внутренней секреции (или эндокринными), т. е. железами, лишенными выводных протоков и выделяющими свои продукты в кровь (реже — в лимфу). Такой способ передачи сигналов, несомненно, более медленный, а действие гормонов часто сказывается одновременно на множестве органов и тканей, тогда как в случае нервной системы воздействие может быть «точечным». Несмотря на эти очевидные недостатки, гормоны абсолютно необходимы не только для регулирования работы различных органов и их систем, но и для индивидуального развития организма.
Структуры, вырабатывающие гормоны, в действительности не составляют единую анатомическую систему, а рассеяны по всему телу, буквально от носа до хвоста, будучи производными самых разных органов или развиваясь в качестве самостоятельных элементов. В этом смысле они отчасти сходны с кроветворными органами: одинаково несущественно, в какой части тела вырабатываются и форменные элементы крови, и гормоны, поскольку и те и другие поступают в кровеносную систему и распределяются по всему организму.
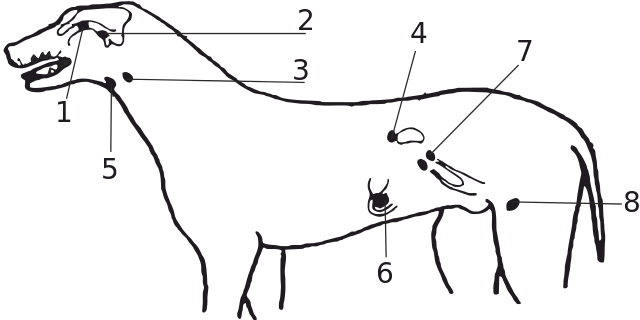
Обобщенная схема эндокринной системы млекопитающего: 1 — эпифиз (шишковидное тело); 2 — гипофиз; 3 — паращитовидные железы; 4 — надпочечники; 5 — щитовидная железа; 6 — поджелудочная железа; 7 — яичники; 8 — семенники.
Нервная и эндокринная системы хотя и обособлены, однако далеко не независимы друг от друга. Гормоны прямо или косвенно могут оказывать на нервную систему мощное воздействие. И наоборот, «верховная железа» эндокринной системы — гипофиз — находится под непосредственным контролем головного мозга. Мозговое вещество надпочечников тоже, хотя и осуществляет внутреннюю секрецию, фактически состоит из видоизмененных нервных клеток. Эндокринными железами попутно являются и многие другие органы: наиболее очевидный пример — гонады, то же самое относится к поджелудочной железе. К эндокринной системе примыкают некоторые органы и ткани, не относящиеся к эндокринным железам, но содержащие эндокринные клетки. К их числу относят сердце, почки, желудочно-кишечный тракт и др. Наконец, секрецией гормонов могут быть заняты отдельные клетки (например, тучные); несколько расширив определение, можно было бы считать их одноклеточными эндокринными органами.
Секреция гормонов происходит только в определенные моменты. Помимо прямого нервного воздействия на эндокринные железы, она может управляться опосредованно, через действие других гормонов. Обычно уровень гормона в крови регулируется по принципу отрицательной обратной связи. Гормоны вырабатываются в очень малых количествах, хотя продолжительность жизни большинства их молекул в крови составляет меньше часа. Для того чтобы присутствие гормонов вызвало необходимый эффект, они должны выделяться непрерывно.
Эндокринная система млекопитающих принципиально сходна с гормональными системами остальных позвоночных животных. Отличия заключаются не столько в составе или числе гормонов, сколько в усилении их активности и специализации; особенно это касается половых гормонов. Кроме того, некоторые гормоны у представителей рассматриваемого класса имеют иное действие, чем у других позвоночных. Тесная взаимосвязь гормональной деятельности желез внутренней секреции друг с другом и с центральной нервной системой обеспечивает согласованное протекание всех жизненных процессов организма, в том числе интенсивности и характера метаболизма и их подстройку к меняющимся условиям внешней среды.
Гипофиз, или нижний мозговой придаток (hypophysis cerebri), — маленький, но очень важный орган, выполняющий функцию гуморального регулятора всех вегетативных процессов в организме. Он находится под промежуточным отделом головного мозга, где помещается в особой ямке дна мозговой коробки — турецком седле (sella turcica). Расположение гипофиза под гипоталамусом важно потому, что эта часть головного мозга сильно влияет на продукцию гипофизарных гормонов, некоторые из которых на самом деле вырабатываются клетками гипоталамуса. Его нейроны синтезируют и секретируют специальные регуляторные пептиды — нейрогормоны, транспортируемые не только в гипофиз, но и в другие структуры мозга и ликвор. Эти нейрогормоны могут действовать на висцеральные органы (окситоцин, вазопрессин), на функции нервной системы (эндорфины, энкефалины, нейротензин, ангиотензин и др.) или же на железистые клетки гипофиза (рилизинг-гормоны: либерины — усиливают секрецию, статины — подавляют).
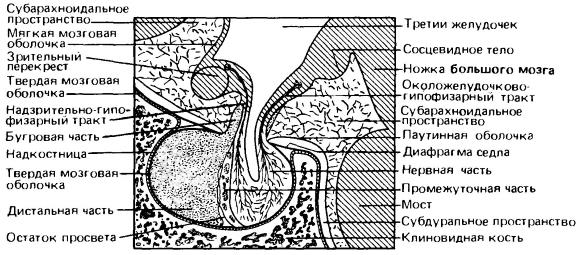
Сагиттальный разрез гипофиза и близлежащих структур в области основания головного мозга у человека.
Гипофиз формируется в эмбриональном развитии из двойного источника — с одной стороны, из нервной ткани в нижней части гипоталамуса, с другой стороны, из участка эктодермы, которая погружается и отшнуровывается в виде пузырька от крыши ротовой полости. Во взрослом состоянии орган разделен на две доли — переднюю железистую, или аденогипофиз (adenohypophysis) и заднюю нервную, или нейрогипофиз (neuro-hypophysis). Аденогипофиз имеет большие размеры и важнее в плане образования гормонов. Он состоит из секреторных клеток и представлен главным образом дистальной частью (pars distalis), но у млекопитающих в нем зачастую выделяется еще бугровая часть (pars tuberalis) — направленный вверх вырост, охватывающий муфтой стебелек воронки, и промежуточная часть (pars intermedia). Последняя в большинстве случаев выражена слабо, особенно в таких группах, как насекомоядные, неполнозубые и хищные. Промежуточная часть имеет тенденцию сливаться с нервной частью (pars nervosa), представляющую собой железистую часть нейрогипофиза и примыкающую к аденогипофизу в турецком седле. К нервной доле следует также относить воронку (infundibulum), т. е. стебелек гипофиза, соединяющий его нервную часть с гипоталамусом, часть которого правильнее было бы тоже относить к нейрогипофизу в качестве его неотъемлемого функционального компонента.
Аденогипофиз представлен скоплениями и тяжами секреторных эпителиальных клеток, разделенными синусоидами и поддерживаемыми каркасом из рыхлой соединительной ткани. В его основной по массе дистальной части имеется два основных типа железистых клеток; при помощи различных гистохимических тестов их можно подразделить еще на несколько групп, каждая из которых предположительно вырабатывает какой-то определенный гормон. Промежуточная часть состоит из многогранных секреторных клеток, а бугровая (если присутствует) — из клеток, по-видимому, практически лишенных секреторной функции.
Нейрогипофиз устроен совершенно по-иному. В нем содержатся большие запасы гормонов, которые могут поступать в ветвящиеся здесь мелкие кровеносные сосуды. Кроме того, здесь присутствуют клетки с ветвистыми отростками; раньше думали, что они и вырабатывают секреты нейрогипофиза, но теперь их считают опорными элементами типа нейроглии других частей центральной нервной системы. В действительности гормоны, обнаруженные в этой «железе», вырабатываются не в ней, а являются нейросекретом ганглионарных клеток гипоталамуса, выделяемым кончиками их аксонов в нервной доле гипофиза.
Из множества мелких ядер гипоталамуса в этом процессе участвуют две пары. Спереди от воронки непосредственно над перекрестом зрительных нервов находится пара ядер, соответственно называемых надзрительными (nuclei supraoptices). Аналогичное положение позади воронки в сером бугре занимают околожелудочковые ядра (nuclei paraventriculares). Из каждой пары вниз, в нервную долю, идут пучки толстых аксонов; по-видимому, небольшое число таких волокон берет начало также в других ядрах гипоталамуса. Тела клеток в названных ядрах обладают секреторной активностью, а их аксоны, хотя, возможно, в некоторой степени и способны передавать обычные нервные импульсы, но специализированы, безусловно, в транспортировке вырабатываемых телами гормонов в нервную долю, где эти последние и скапливаются перед выходом в кровь.
На гипофиз, особенно передний железистый, сильно влияют другие органы внутренней секреции. Это легко осуществимо, так как обе его доли снабжаются веточками внутренней сонной артерии, а к ганглиям гипоталамуса, вырабатывающим гормоны нервной доли, подходят внутричерепные сосуды. На аденогипофиз, кроме того, в значительной мере воздействует головной мозг, причем механизм этого влияния весьма примечателен. В эту долю гипофиза входит несколько нервных волокон, так что можно было бы предположить, что его секреторная активность находится под прямым нервным контролем. Но входя в промежуточную часть, волокна едва ли проникают в дистальную часть, вырабатывающую почти все гормоны (если не считать некоторых, по-видимому, вегетативных, волокон, идущих к кровеносным сосудам).
На самом деле, воздействие осуществляют рилизинг-гормоны, которые вырабатываются гипоталамусом и доставляются в аденогипофиз через любопытно организованную локальную воротную систему. Некоторые артерии, снабжающие аденогипофиз, проходят перед воронкой у самого дна промежуточного мозга или в толще его, где как раз пролегает один из нервных трактов, несущих нейросекрет в нервную долю. В этом месте нижняя поверхность головного мозга млекопитающих часто образует вздутие, называемое срединным возвышением (eminentia mediana). При этом упомянутые артерии сами не достигают железистой доли, а проникают в толщу срединного возвышения, где распадаются на капилляры и затем вновь собираются в пучок мелких воротных вен, которые и входят в дистальную часть. По пути сквозь срединное возвышение кровь подхватывает нейрогормоны, в виде которых и передается в аденогипофиз информация от головного мозга. Может показаться странным, что столь важная функция выполняется таким окольным путем, но, поскольку железистая доля не снабжена нервами, других эффективных способов не остается.
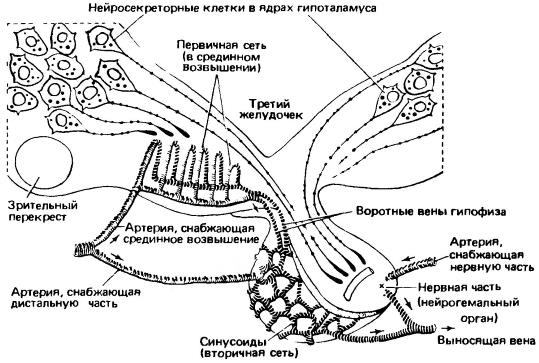
Схема структур, связывающих гипоталамус и гипофиз. Клетки определенных ядер гипоталамуса вырабатывают нейросекреты, которые спускаются по аксонам этих клеток в нервную часть гипофиза и передаются из аксонов в кровь в качестве гормонов. Другие секреторные клетки выводят свои продукты в срединное возвышение, расположенное в основании промежуточного мозга; здесь эти вещества попадают в первичную сеть воротной системы гипофиза, уносятся в синусоиды дистальной части аденогипофиза и стимулируют в ней синтез гормонов.
Известно около десятка гормонов гипофиза; предполагается существование и других. Большинство гормонов вырабатывается пятью группами хромафильных клеток аденогипофиза, расположенными главным образом в его дистальной части. Все это гликопротеины, белки или полипептиды. Ниже приводится их перечень с краткими характеристиками.
Гормон роста, или соматотропин, влияет на самые разнообразные ростовые процессы и обмен веществ в целом. В первую очередь он стимулирует рост скелета и мускулатуры, регулирует обмен жиров и углеводов, синтез белков. Кроме того, этот гормон усиливает влияние других гормонов на деятельность щитовидной железы, коры надпочечников и гонад.
Адренокортикотропный гормон, или кортикотропин, стимулирует эндокринную функцию коры надпочечников.
Тиреотропин регулирует образование и высвобождение гормонов щитовидной железой.
Пролактин стимулирует рост млечных желез и секрецию молока, а у ряда млекопитающих также продлевает время функционирования желтого тела и секреции прогестерона. Примечательно, что у пресноводных рыб пролактин регулирует водно-солевой обмен, у земноводных — тормозит метаморфоз, а у птиц — регулирует метаболизм жиров и репродуктивные функции.
Лютеинизирующий гормон регулирует созревание гонад и секрецию половых гормонов, влияет на формирование желтого тела и секрецию прогестерона в яичниках, а в семенниках стимулирует секрецию интерстициальными клетками андрогенов, влияя тем самым на созревание сперматозоидов.
Фолликулостимулирующий гормон стимулирует рост фолликулов в яичниках и сперматогенез в семенниках, а также, вместе с предыдущим гормоном, секрецию эстрогенов и овуляцию. Оба этих гормона объединены в группу гонадотропинов; вырабатываются они одной и той же группой хромафильных клеток.
Меланоцитстимулирующий гормон, или интермидин, — полипептид, который у рыб, амфибий и рептилий вызывает рассредоточение пигментных гранул в меланофорах и сосредоточение отражательных органелл в иридиофорах. У млекопитающих МСГ не только влияет на распределение пигментов в меланоцитах, но воздействует также на обмен веществ и отвечает за чувство голода. В противоположность гормонам, перечисленным выше, этот гормон вырабатывается в виде нескольких разновидностей промежуточной частью аденогипофиза, если та четко выражена.
В отличие от аденогипофиза, вырабатывающего множество гормонов, нейрогипофиз млекопитающих секретирует лишь небольшое число сходных между собой полипептидов. Это в первую очередь вазопрессин, или антидиуретический гормон, а также окситоцин. Оба гормона представлены цепочкой из восьми аминокислот, шесть из которых у них совпадают. Как и следует ожидать, их функции в некоторой степени пересекаются. Вазопрессин в основном участвует в регуляции процессов отдачи и поглощения воды в почечных канальцах, тем самым влияя на уровень кровяного давления и мочеотделение. Функции окситоцина понятнее всего у самок млекопитающих, у которых он стимулирует сокращение мускулатуры матки, развитие млечных желез и выделение молока после рождения потомства. Таким образом, секреты нервной доли гипофиза влияют главным образом на сократительные элементы (в первую очередь на гладкую мускулатуру) и на полупроницаемые мембраны.
Шишковидное тело, или эпифиз (epiphysis cerebri), является главной составной частью т. н. фотоэндокринной системы. Его секреторные клетки выделяют в кровь гормон мелатонин, синтезируемый из серотонина, который участвует в синхронизации циркадных ритмов (биоритмов «сон - бодрствование») и, возможно, влияет на все гипоталамо-гипофизарные гормоны, а также иммунную систему. Путем восстановления серотонина синтезируется адреногломерулотропин, стимулирующий выработку надпочечниками альдостерона.
К железистым производным глотки принадлежат щитовидная железа, паращитовидные железы, тимус и ультимобранхиальные тельца.
Щитовидная железа (glandula thyroidea) располагается в области шеи вентральнее трахеи. Название соответствует ее внешнему виду у человека. Развивается данная железа из слизистой оболочки брюшной стенки глотки, первоначально формируя углубление, дно которого погружается в мягкие ткани и отшнуровывается в виде небольшого тела. У некоторых млекопитающих оно остается цельной структурой, но в большинстве случаев подразделяется на две отчетливые доли. Железа богата кровеносными сосудами и состоит из множества мелких округлых фолликулов (folliculae glandulae thyroideae), связанных воедино соединительной тканью. Стенка каждого из них образована кубическим секреторным эпителием, клетки которого окружают центральную полость и наполняют ее студенистым коллоидом. В последнем содержится большой запас иодсодержащего белка тиреоглобулина, гидролиз которого дает гормоны тироксин и трийодтиронин. Они (особенно вырабатываемый в большем количестве тироксин) играют крайне важную роль в регуляции общего обмена веществ. Интенсивность метаболизма, особенно связанного с окислением, а соответственно и температура тела млекопитающих поддерживаются на высоком уровне в значительной степени благодаря этим гормонам. Выделяемые железой гормоны также влияют на процессы роста и созревания, половые функции, развитие нервной системы и поведение. Парафолликулярными клетками вырабатывается гормон кальцитонин, снижающий уровень кальция в крови. Эндокринная активность щитовидной железы в значительной степени контролируется гипофизом, но она реагирует и на ряд внешних воздействий. Так, во время зимней спячки активность понижается.
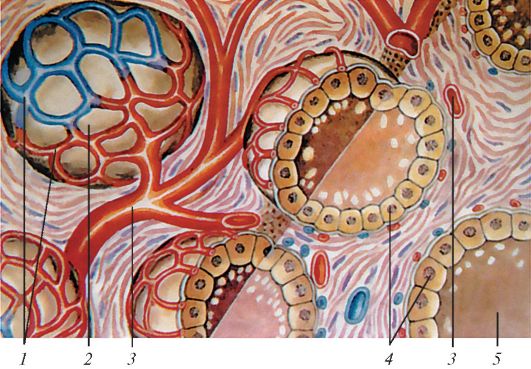
Схема строения фолликулов щитовидной железы человека: 1 — кровеносные капилляры; 2 — фолликул; 3 — артерия; 4 — железистые клетки; 5 — коллоид, содержащий гормоны щитовидной.
Паращитовидные железы (glandulae parathyroideae) обычно представлены двумя парами маленьких округлых телец, расположенными по соседству с щитовидной железой, а иногда (например, у человека) и погруженными в нее. Они происходят от жаберных мешков примитивных позвоночных и представляют собой плотные скопления эпителиальных клеток. В основном это т. н. главные клетки, занятые эндокринной деятельностью; у некоторых млекопитающих также имеется небольшое количество оксифильных клеток. Вырабатываемый железами белковый гормон паратгормон является антагонистом кальцитонина, регулируя в организме концентрацию кальция и (в меньшей степени) выделение фосфора. Для нервно-мышечных взаимодействий в жидкостях тела необходим вполне определенный уровень кальция; если его слишком мало, могут наступить тетания и смерть. Большую часть вещества кости и зубной ткани составляет фосфат кальция, который непрерывно обновляется, и если не поступает достаточного количества «нового» кальция и фосфора, скелет и зубы дегенерируют. Кроме того, уровень концентрации кальция влияет на нервную возбудимость, свертываемость крови, проницаемость клеточных мембран и функционирование различных желез.
Тимус, называемый также вилочковой железой (glandula thymus), представляет собой дольчатый орган, лежащий в передней части грудной полости в области трахеи. Достигая сильного развития у молодых животных, с возрастом он претерпевает обратное развитие. Выше отмечалось, тимус является местом созревания специфических Т-лимфоцитов. Им выделяются гормоны группы тимозинов и тимопоэтин, которые управляют дифференцировкой T-лимфоцитов, а также регулируют процессы синтеза клеточных рецепторов к гормонам и нейромедиаторам, стимулируют разрушение ацетилхолина в нервно-мышечных синапсах и участвуют в регуляции других процессов, важных для всего организма.
Ультимобранхиальные тельца — маленькие эпителиальные образования, которые у взрослых млекопитающих погружены в ткань щитовидной и паращитовидных желез. Как и в них, в тельцах присутствуют С-клетки, вырабатывающие кальцитонин.
Хотя большая часть ткани поджелудочной железы вырабатывает пищеварительные ферменты, поступающие по протокам в кишечник, в ней также имеются участки другой ткани, рассеянные в виде изолированных панкреатических островков, или островков Лангерганса. Они состоят из секреторных клеток нескольких типов, не снабженных выводными протоками, т. е. выделяющих свои секреты в кровь, формируя таким образом эндокринный орган. β-клетки вырабатывают особый белковый гормон инсулин — важный регулятор обмена веществ, особенно углеводов. Он повышает потребление углеводов тканями, сдерживает образование глюкозы в печени, облегчает ее проникновение в клетки некоторых типов (например, мышечные), поддерживая тем самым определенный уровень глюкозы в крови. Секреция инсулина слабо контролируется нервной системой, а зависит главным образом от концентрации глюкозы в крови, проходящей через островки. α-клетки выделяют полипептид глюкагон, который способствует расщеплению запасенного в печени гликогена и в результате вызывает повышение концентрации глюкозы в крови. Кроме того, имеются δ-, ε- и РР-клетки, вырабатывающие соответственно соматостатин, грелин и панкреатический полипептид. При этом соотношение клеточных составляющих островков Лангерганса у разных млекопитающих различно. Так, у грызунов почти вся центральнаю часть островка занята β-клетками, а остальные клетки оттеснены к периферии; у человека же α- и β-клетки равномерно располагаются по всему островку.
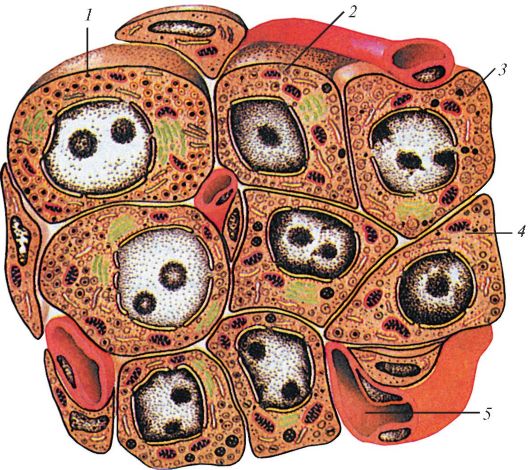
Схема строения панкреатического островка человека: 1 — α-клетка; 2 — β-клетка; 3 — δ-клетка; 4 — РР-клетка; 5 — кровеносный капилляр.
Печень, выделяющая желчь, тоже секретирует ряд гормонов: ангиотензин, тромбопоэтин, гепцидин, бетатрофин, а также инсулиноподобный фактор роста 1.
По соседству с каждой почкой, в виде шапочек на их передних концах присутствуют небольшие органы, называемые надпочечниками (glandulae suprarenales). В составе их компактных телец можно обнаружить две различные ткани, которые образуют четко разграниченные корковый и мозговой слои. Оба эти компонента — железы внутренней секреции, но совершенно различные, причем мозговая ткань в отличие от корковой является производным нервной системы. Гормоны коркового вещества позволяют выдерживать длительные внешние стрессы, в частности, вызывая напряжение окислительных процессов и ряда других функций организма. Гормоны мозгового вещества действуют более массировано, помогая пережить краткие критические ситуации.
Всего из коры надпочечников выделено около 50 стероидных гормонов. Некоторые из них, по-видимому, не обладают гормональной активностью; другие (содержащиеся в небольших количествах) химически очень близки к половым гормонам гонад; третьи — это специфические гормоны коры надпочечников; примерно 6 из них — важнейшие. Специфические стероидные гормоны коркового вещества (адренокортикостероиды) делятся на две основные группы. Глюкокортикоиды включают кортизол, кортизон, кортикостерон и др. Эти гормоны различным образом влияют на метаболизм: в первую очередь стимулируют расщепления белков, накопление гликогена в печени и увеличение концентрации глюкозы в крови; таким образом, они действуют как антагонисты инсулина. Синтез и секреция глюкокортикоидов стимулируются адренокортикотропным гормоном. Другая группа — это минералокортикоиды; сюда относят, в частности, 11-дезоксикортикостерон и альдостерон, усиливающие обратное всасывание натрия и выделение калия в почках. Секреция альдостерона регулируется не адренокортикотропным гормоном, а ренин-ангиотензиновой системой и зависит от концентрации в крови калия. Хотя, как только что говорилось, функции двух групп адренокортикостероидов в обмене веществ весьма различны, они все же в некоторой степени перекрываются. Роль нескольких гормонов коркового вещества в регуляции водно-солевого обмена указывает на возможную связь между этим веществом и почками. Действительно, ткани почки и коры надпочечника происходят из соседних участков зародышевой мезодермы. Заслуживает внимания и то, что в этой же области формируются эндокринные ткани гонад — только они и кора надпочечников вы-рабатывают стероидные гормоны.
В центре надпочечников сконцентрировано мозговая ткань, по происхождению и функции очень сильно отличающаяся от корковой. Выше было показано, что вегетативная двигательная иннервация внутренних органов тела осуществляется особым образом: ее импульсы проходят к гладкой мускулатуре или железам не напрямую, а через постганглионарные нейроны. Клетки мозговой ткани надпочечников представляют собой именно такие нейроны, сильно видоизмененные для осуществления внутренней секреции.
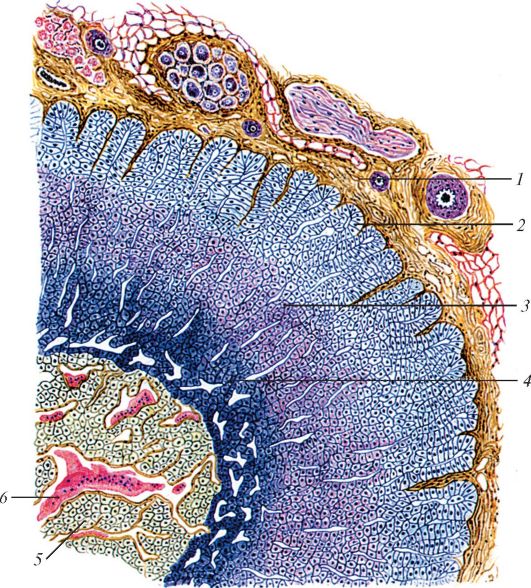
Микроскопическое строение надпочечника человека: 1 — капсула надпочечника; 2 — клубочковая зона; 3 — пучковая зона; 4 — сетчатая зона; 5 — мозговое вещество; 6 — синусоидный капилляр.
Мозговая ткань надпочечников иннервируются преганглионарными вегетативными нервными волокнами. При стимуляции она выделяют в кровь два катехоламина — адреналин и норадреналин, влияющие на кровообращение, углеводный обмен и работу нервной системы. Эти же гормоны выделяются из постганглионарных волокон симпатической системы, однако в первом случае больше адреналина, а во втором — норадреналина. Клетки мозгового вещества не похожи на нервные, так как не имеют отростков. Но поскольку они гомологичны постганглионарным симпатическим нейронам, неудивительно, что ими вырабатываются такие же продукты. Разница в том, что настоящая постганглионарная клетка дает лишь крошечное их количество, воздействующее на ближайшие структуры, а мозговая ткань надпочечников может быстро выделять крупные дозы этих веществ. Кровь разносит их по всему телу, и они производят на различные органы сильное одновременное воздействие, как бы безадресно распространяя эффект симпатической нервной системы. Помимо экстренных обильных выбросов, адреналин и норадреналин постоянно выделяются и в нормальной ситуации, хотя в значительно меньшем количестве.
Размножение и связанные с ним явления подлежат гормональному регулированию в степени значительно большей, чем любые другие функции организма. Синтезируемые гонадами гормоны стимулируют не только развитие вспомогательных структур половой системы, таких как половые протоки и копулятивные органы, но также и развитие вторичных половых признаков, к числу которых относят проявления полового диморфизма в характере волосяного покрова, млечные железы, а во многих случаях даже размеры и пропорции тела. В то же время, гормоны влияют на цикличность половых процессов, беременность, лактацию и особенности поведения животных. При этом действие половых гормонов выступает отдельным звеном в целой «цепочке команд». Гонады вырабатывают свои гормоны не чисто спонтанно, а в значительной степени под действием гормонов гипофиза. В свою очередь гипофиз находится под сильным влиянием примыкающего к нему гипоталамуса, и, кроме того, в порядке обратной связи — под влиянием гормонов гонад и других органов. Наконец, головной мозг в регуляции связанных с размножением гормональных процессов подвержен сильному воздействию различных факторов, как действующих внутри организма, так и внешних.
Все половые гормоны, синтезируемые гонадами, кроме одного, являются стероидами. Гормоны, вырабатываемые главным образом в семенниках, называются андрогенами, а в яичниках — эстрогенами. Эти стероидные гормоны по химической структуре очень похожи на гормоны коры надпочечников. В сущности, они настолько близки, что гонады и кора надпочечников в качестве органов внутренней секреции, так сказать, не могут полностью поделить между собой функции. В составе секрета коры надпочечников можно обнаружить небольшое количество половых гормонов, а в секрете гонад небольшую долю составляют специфические гормоны коры надпочечников. Вдобавок состав секрета мужских и женских гонад отчасти перекрывается: во всех проверенных случаях обнаружено, что семенники секретируют некоторое количество женских половых гормонов, а яичники — мужских.
Белок релаксин — нестероидный гормон женских гонад, который разрыхляет тазовый симфиз, облегчая рождение детенышей. На самом деле он представляет собой исключение и в том, что вырабатывается не только яичником, но также маткой и плацентой.

Андрогенные гормоны формируют в окраске мандрила (Mandrillus sphinx) яркие цвета.
Основными гормонами семенников, т. е. андрогенами, являются тестостерон и андростендион. Из семенника выделены и другие соединения, но, возможно, они являются метаболическими производными этих действенных и, по-видимому, первичных продуктов, которые при экспериментальном введении обычно очень эффективно стимулируют развитие вторичных половых признаков и поведения, свойственных самцу. В семенниках выработкой половых гормонов занимаются особые интерстициальные клетки. Эти клетки, как указывает их название, не входят в состав семенных канальцев, а располагаются (вместе с соединительной тканью) в промежутках между ними (т. е. интерстициально). В стенках канальцев помимо половых клеток, дающих сперматозоиды, находятся поддерживающие клетки Сертоли. Хотя сами клетки Сертоли не вырабатывают гормонов, их секрет связывает андрогены и тем самым поддерживает высокую концентрацию последних внутри семенников.
Некоторое количество интерстициальных клеток может находиться и в яичниках, но большая часть эмбриональной ткани, из которой они образуются, превращается в фолликулярные клетки, окружающие развивающиеся яйцеклетки и снабжающие питательными веществами. Помимо этого, фолликулярные клетки являются основным источником гормонов яичников (эстрогенов), которые секретируются также имеющимися интерстициальными клетками. Из эстрогенов основными являются стероидные гормоны: эстрадиол и эстрон; из них первый значительно эффективнее. В противоположность андрогенам, эстрогены стимулируют развитие и сохранение вторичных половых признаков и полового поведения у самок.
В яичниках кроме первичных половых гормонов может происходить образование дополнительных стероидных гормонов. По выходе яйцеклетки из яичника вмещавший ее фолликул, казалось бы, должен сразу дегенерировать, однако у млекопитающих это происходит с значительной задержкой. При этом фолликулярные клетки увеличиваются в размерах и приобретают желтый цвет. Такие гипертрофированные клетки заполняют опустевший фолликул, образуя желтое тело (corpus luteum). Оно секретирует важный стероидный гормон — прогестерон, который подготавливает эпителий матки к имплантации яйцеклетки и, если оплодотворение и имплантация произошли, стимулирует развитие плаценты.
Сама плацента в течение беременности также служит органом внутренней секреции, вырабатывая ряд гормонов. В первую очередь, это хорионический гонадотропин, тормозящий развитие фолликулов в яичниках матери и поддерживающий синтез прогестерона и эстрогена в желтом теле до тех пор, пока плацента не будет способна сама синтезировать их достаточные количества. Помимо хорионического гонадотропина, эстрогена и прогестерона плацента синтезирует кортикотропин-рилизинг-гормон (кортиколиберин), плацентарный лактоген и проопиомеланокортин.

Турнирная схватка самцов конгони (Alcelaphus buselaphus). Повышенный уровень половых гормонов в период спаривания делает самцов многих видов особенно агрессивными.
Слизистая оболочка желудка и двенадцатиперстной кишки секретирует множество пептидных гормонов, оказывающих преимущественно местное действие. Однако здесь все происходит несколько иначе, чем в уже рассмотренных случаях. Если большая часть гормонов образует взаимозависимую систему, часто подавляя секрецию и уравновешивая действие друг друга, то выделение гастроинтестинальных (желудочно-кишечных) гормонов, по-видимому, не подвержено влиянию других гормонов, и даже гормонов, принадлежащих к этой же группе. Кроме того, их вырабатывают не специальные железы внутренней секреции, а ткани, выполняющие также и другие функции.
В ответ на попадание пищи слизистой пилорической части желудка выделяется гормон гастрин, который уносится кровью к дну желудка, где стимулирует секрецию соляной кислоты, пепсина и активирует моторику желудка. Париетальные клетки желудка вырабатывают грелин, усиливающий аппетит. Холецистокинин, секретируемый преимущественно в двенадцатиперстной кишке, стимулирует образование ферментов поджелудочной железой, т. е. обогащение ее сока, а также опорожнение желчного пузыря. Слизистая переднего отдела тонкого кишечника выделяет в кровь полипептид секретин, стимулирующий подачу в кишечник сока поджелудочной железы, а также в некоторой степени подачу желчи и секрецию кишечных соков. Секреция этого гормона, как и предыдущего, вызывается попаданием пищи в тонкий кишечник и активизирует протекающие здесь пищеварительные процессы. На основании физиологических эффектов утверждается существование и других подобных гормонов.
Кардиомиоциты сердца выделяют предсердный натрийуретический пептид — мощный вазодилататор и антагонист ренин-ангиотензин-альдостероновой системы. В предсердиях в малых количествах образуются также соматостатин и ангиотензин-II.
Некоторые клетки почек вырабатывают гормоны кальцитриол (третий кальцийрегулирующий гормон), ренин (начальное звено ренин-ангиотензин-альдостероновой системы, влияющей на уровень кровяного давления) и эритропоэтин (стимулирует эритропоэз — образование эритроцитов в красном костном мозге).
Клетки жировой ткани (адипоциты) секретируют гормон лептин, подавляющий аппетит и выступающий тем самым антагонистом грелина.
Тучные клетки соединительной ткани вырабатывают гепарин, гистамин и серотонин.
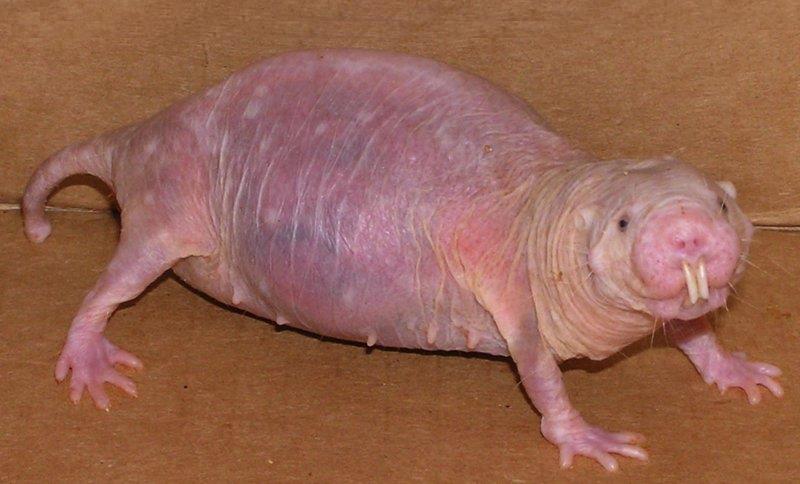
У голых землекопов (Heterocephalus glaber) единственная размножающаяся самка путем выделения феромонов и направленной агрессии подавляет половое развитие у подчиненных членов колонии.