в оглавление
назад
Головной мозг (продолжение)
Наибольшей величины и сложности у млекопитающих достигает передний, или конечный, мозг (telencephalon), состоящий из двух больших полушарий (hemispheri cerebri). Судя по всему, полушария возникли главным образом (а быть может, и исключительно) в связи с обонятельной рецепцией. Запахи не так много значат в жизни высших приматов, в том числе и человека. Однако на более ранних стадиях эволюции, вплоть до предков позвоночных, обоняние являлось главнейшим каналом, через который животные получали сведения об окружающем мире. Поэтому вполне естественно, что обонятельные центры мозга и послужили тем основанием, на котором впоследствии сложились сложные нервные механизмы. Уже на ранних этапах эволюции тетрапод полушария превращаются в крупные и важные центры корреляции сенсорных сигналов. Ко времени, когда появились млекопитающие, сильно разросшаяся поверхность полушарий стала доминирующим ассоциативным центром, местом локализации высшей умственной деятельности. У различных представителей класса отношение массы полушарий переднего мозга к массе всего головного мозга варьирует: у ежа (Erinaceus europaeus) оно составляет 48 %, у белки (Sciurus vulgaris) — 53 %, у волка (Canis lupus) — 70 %, у белобочки (Delphinus delphis) — 75 %, у большинства приматов — 75-80%, у человека — около 85%. У птиц большие полушария по массе примерно соответствуют остальному мозгу или уступают ему, иногда в несколько раз. Наконец, о чрезвычайном значении больших полушарий говорит то, что их разрушение приводит к полной функциональной несостоятельности млекопитающего.
Снизу к передней части полушарий примыкают обонятельные луковицы (bulbi olfactorii). Эти образования наиболее развиты у животных с хорошим обонянием и сильно редуцированны у чисто водных форм. По различию в их развитии отличают несколько типов структуры мозга. У сумчатых, насекомоядных, неполнозубых, хищных, грызунов и некоторых других обонятельные луковицы велики и хорошо выступают вперед при взгляде на мозг сверху. Такой тип мозга с совершенным развитием обонятельных долей называется макросматическим. У ластоногих, сирен и многих приматов луковицы слабо развиты; этим животным свойствен микросматический мозг. Наконец, для китообразных обычен т. н. аносматический мозг с редуцированными обонятельными луковицами. Раньше считалось, что способность различать химические сигналы у китов и дельфинов полностью утеряна, однако оказалось, что это не совсем так.
Поверхностные слои полушарий переднего мозга млекопитающих образуют паллиум, или мозговой свод (pallium). Верхняя прослойка, состоящая из тел нейронов и безмякотных нервных волокон, называется корой (cortex cerebri) и представляет собой серое вещество свода. Тела нейронов расположены в коре послойно, образуя своеобразные экранные структуры. Такая организация мозга позволяет пространственно отображать внешний мир на основе информации, поступающей от органов чувств. Экранные структуры характерны для важнейших мозговых центров млекопитающих, тогда как у других позвоночных они встречаются реже, преимущественно в зрительных центрах. Под корой находится прослойка из миелинизированных отростков нейронов — белое вещество свода. Нервные волокна белого вещества образуют проводящие пучки, спаивающие полушария — мозговые комиссуры. Хотя основная часть серого вещества сосредоточена в коре, оно имеется и в подкорковом слое, где представлено относительно небольшими скоплениями — ядрами. В их число входят полосатые тела (corpora striata), лежащие под полостью бокового желудочка и получившие свое название из-за пересекающих их нервных волокон. Под контролем коры полосатые тела выполняют функцию регуляции стереотипных, автоматизированных реакций — безусловных рефлексов.
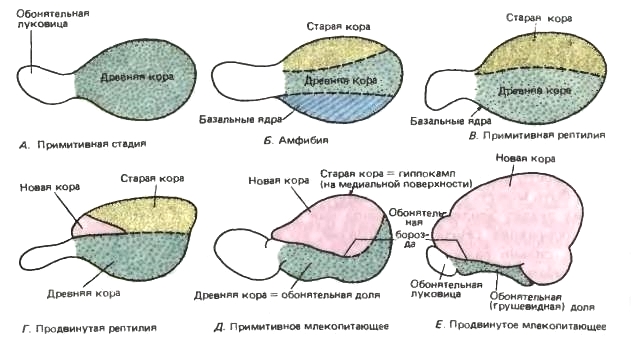
Схемы, отражающие прогрессивное развитие полушарий переднего мозга. Вид сбоку на полушарие с обонятельной луковицей. Различные отделы, отличающиеся друг от друг цитологически, окрашены по-разному.
А — полушарие представляет собой всего-навсего обонятельную долю.
Б — дифференцируются дорсальный отдел — архипаллиум (= гиппокамп) и вентральный отдел — базальное ядро (полосатое тело).
В — базальные ядра переместились во внутреннюю область полушария.
Г — появляется небольшой участок, представляющий собой неопаллиум.
Д — палеопаллиум оттесняется на медиальную поверхность полушария, но неопаллиум все еще имеет скромные размеры, и под обонятельной бороздой сохраняются значительно развитые обонятельные отделы.
Е — примитивный обонятельный отдел сохраняется только в вентральной области, и неопаллиум достигает чрезвычайно сильного развития.
Свод мозга начал формироваться еще у рыб. В связи с прогрессивным развитием нюховой сенсорной системы у них возникает палеопаллиум, или древний свод (paleopallium), полностью покрывающий небольшие полушария. На стадии лопастеперых рыб в дорсальной части полушарий ближе к оси тела появляется архипаллиум, или старый свод (archipallium). У амфибий и примитивных рептилий он получает дальнейшее развитие, вследствие чего палеопаллиум оттесняется и сохраняется только вдоль боковой поверхности полушарий. При этом палеопаллиум продолжает носить преимущественно обонятельный характер и на высших стадиях эволюции формирует обонятельные доли коры полушарий. Архипаллиум в определенной, хотя и малой, степени является коррелятивным центром, принимая восходящие волокна из промежуточного мозга, а также волокна из обонятельной луковицы и обонятельной доли; по всей видимости, он также связан с эмоциональным поведением. Нервный путь из этого участка в гипоталамус составляет основной элемент пучка волокон, называемого у млекопитающих сводом (fornix).
У амфибий впервые появляется зачаток неопаллиума, или нового свода (neopallium). У рептилий неопаллиум уже образует небольшую область между древним и старым сводом. С самого начала своей эволюции этот участок представляет собой ассоциативный центр, подобно базальным ядрам принимающий волокна, которые переключают на него сенсорные сигналы из ствола мозга и, наоборот, передают команды непосредственно в двигательные столбы.
У однопроходных неопаллиум все еще вклинивается между палеопаллиумом снаружи и архипаллиумом изнутри. У сумчатых он разрастается за пределы крыши и боковых стенок полушарий. Архипаллиум при этом вытесняется на медиальную поверхность, а палеопаллиум ограничивается вентролатеральной частью полушария, расположенной ниже носовой борозды (fissura rhinalis) — канавки, которая представляет собой границу между обонятельной и необонятельной областями коры. У плацентарных в связи с дальнейшим усложнением и экспансией неопаллиума полушария достигают такой величины, что превышают по объему остальные отделы мозга вместе взятые. Полушария разрастаются назад и в стороны, последовательно прикрывая промежуточный мозг, средний мозг и часть мозжечка. Парные желудочки и древние структуры, отвечающие главным образом за обоняние (обонятельные луковицы, старые слои мозгового свода и связанные с ними нервные пучки и ядра), в мозге плацентарных оттеснены и деформированы. Так, палеопаллиум сохраняется на вентральной поверхности полушария в виде маленького обонятельного участка, именуемого грушевидной долей (lobus piriformis), а архипаллиум свернут в глубине складки височной доли в рулон под названием гиппокамп (hippocampus). Превосходство больших полушарий над прочими отделами мозга заметно у всех млекопитающих, но особенно ярко оно выражено у продвинутых форм, например у человека. Доминируют полушария и в функциональном отношении. В отличие от млекопитающих, усложнение переднего мозга у птиц выражается главным образом в разрастании базальных ядер (nuclei basales), а не остающегося тонким свода.
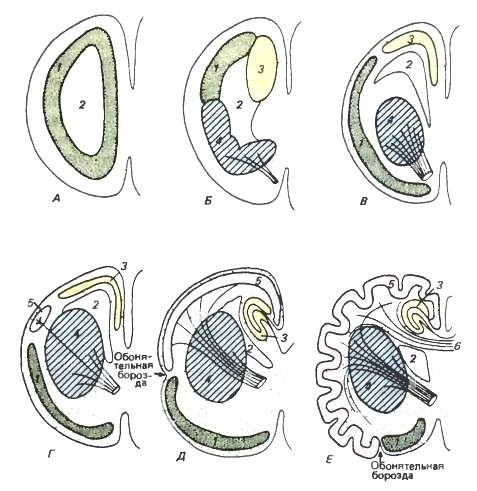
Схематизированные срезы через левое полушарие переднего мозга. Цветовые обозначения такие же, как на предыдущем рисунке.
1 — палеопаллиум; 2 — боковой желудочек; 3 — архипаллиум; 4 — базальные ядра; 5 — неопаллиум; 6 — мозолистое тело.
А — примитивная стадия. Полушарие, по сути дела, представляет собой обонятельную долю. Слабо дифференцированное серое вещество располагается внутри мозга.
Б — стадия, наблюдаемая у современных амфибий. Серое вещество помещается все еще далеко от наружной поверхности, но уже подразделено на палеопаллиум (= обонятельная доля), архипаллиум (= гиппокамп) и базальные ядра (= полосатое тело). Последнее приобретает значение ассоциативного центра, имеющего афферентные и эфферентные связи с таламусом (изображены линиями, символизирующими перерезанные пучки волокон).
В — более прогрессивная стадия, на которой базальные ядра погружаются внутрь полушария, тогда как отделы коры несколько продвинулись кнаружи.
Г — стадия, на которой находятся продвинутые рептилии. Появляется неопаллиум.
Д — стадия примитивного млекопитающего. Неопаллиум увеличился. Он имеет обширные связи со стволом мозга. Архипаллиум на медиальной поверхности полушария заворачивается в качестве гиппокампа. Палеопаллиум пока еще сильно развит.
Е — стадия высокоорганизованного млекопитающего. Неопаллиум чрезвычайно разрастается и собирается в складки. Палеопаллиум занимает ограниченную вентральную область, представляя собой грушевидную долю. Развивается мозолистое тело — мощная перемычка, соединяющая области неопаллиума двух полушарий.
Кора неопаллиума именуется новой корой, или неокортексом (neocortex). У млекопитающих она служит центром высшей (условнорефлекторной) нервной деятельности, координирующим работу других отделов мозга. Отсюда посылаются импульсы к различным органам и тканям тела, здесь же осуществляется регуляция физиологических процессов соответственно условиям среды. Именно новой корой накапливаются следы единичных возбуждений и их сочетаний, вследствие чего обогащается оперативная память, обеспечивающая возможность выбора оптимальных решений в новых ситуациях. Чаше эти решения представляют собой новые комбинации уже известных ранее поведенческих элементов, но происходит также выработка и закрепление новых вариантов действий. По мере своего развития новая кора не только принимает на себя функции коррелятивного и ассоциативного центра вновь возникающих типов высшей нервной деятельности, но и начинает выполнять многие функции, принадлежавшие ранее центрам ствола мозга и базальных ядер. При этом древние центры, управляющие инстинктивными актами, не ликвидируются, а лишь подчиняются высшему контролю.
В связи с развитием новой коры утрачивает свое былое значение крыша среднего мозга, оставаясь лишь рефлекторным и передаточным центром. Слуховые и прочие соматические сенсорные импульсы передаются вперед к таламусу, здесь же прерывается большая часть зрительных волокон, и все эти сигналы из таламуса передаются в полушария по мощным нервным пучкам. Сходные таламические связи с базальными ядрами возникли еще у низкоорганизованных групп позвоночных и получили наибольшее развитие у птиц. В отличие от птиц, у млекопитающих основная масса волокон проходит через полосатые тела насквозь и расходится к поверхности новой коры. Таким образом, к ней стекается полный набор сенсорных данных, на основе которых в коре принимаются соответствующие двигательные «решения».
Как уже упоминалось, часть сигналов передается из коры в мозжечок через мост и обеспечивает необходимые регуляторные эффекты. У новой коры также имеются связи с полосатым телом и даже с гипоталамусом — и тем самым с вегетативной нервной системой. Однако основная масса двигательных команд направляется по пирамидному пути (tractus corticospinalis) — особому нервному пучку, который непосредственно, без переключений, идет от коры больших полушарий через средний мозг к соматическим двигательным областям мозгового ствола и спинного мозга. При этом волокна латеральной части этого пути перекрещиваются и иннервируют противоположную сторону тела (т. е. левое волокно иннервирует правую часть тела, и наоборот), а вентральные волокна сохраняют связь со своей стороной тела. Пирамидный путь имеется лишь у млекопитающих, что ярко демонстрирует доминирующее положение у них новой коры. Наибольшего развития эта структура достигает у обезьян и, особенно, у человека, играя важную роль в прямохождении. У сумчатых пирамидные аксоны доходят только до грудного отдела, а у однопроходных пирамидный путь полностью отсутствует.
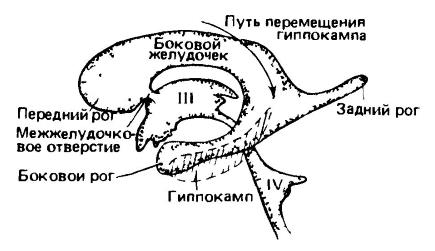
Желудочки мозга человека; вид сбоку с левой стороны. Желудочки представлены в виде отливки, а ткани мозга не изображены. С разрастанием полушария переднего мозга боковой желудочек распространился назад с образованием заднего рога в затылочной доле, а в своей боковой части — вниз и вперед с образованием бокового рога в височной доле. Подобные разрастания, направленные назад и вниз, привели к изменениям в расположении различных частей мозга. Гиппокамп, развившийся в дорсальном положении на медиальной поверхности полушария, у высокоразвитых млекопитающих переместился назад и вниз в вентральное положение.
Поскольку новая кора представляет собой тонкий листок слоистого клеточного материала, под которым лежит белая волокнистая масса мозга, простое увеличение объема полушарий не может осуществить пропорциональное разрастание коры. Вместе с тем, у продвинутых форм площадь коры может значительно увеличиться вследствие ее складчатости. Образовавшиеся подобным образом складки называются извилинами (gyri), а глубокие щели между ними — бороздами (sulci). И те, и другие содержат общие морфологические компоненты. В простейшем случае имеется одна глубокая сильвиева борозда, отделяющая лобную долю (lobus frontalis) от височной (lobus temporalis). Затем выше и кпереди от сильвиевой борозды появляется поперечно идущая роландова борозда, отделяющая сверху лобную долю от теменной (lobus parietalis). У приматов поперечная борозда отделяет небольшую заднюю затылочную долю (lobus occipitalis). Кроме основных борозд, образуется много добавочных; их число особенно велико у приматов и зубатых китов. Раньше считали, что борозды в некоторых случаях обозначают морфологические границы, соответствующие определенным областям коры. Однако дальнейшие исследования показали отсутствие фиксированной связи между распределением складчатости и структурным подразделением коры (если не считать носовой борозды и в какой-то мере центральной борозды у приматов, о чем еще будет сказано). Примечательно, что складчатость коры развилась в нескольких эволюционных стволах млекопитающих совершенно независимо. У относительно примитивных млекопитающих, таких как однопроходные, сумчатые, а также некоторые плацентарные (насекомоядные, рукокрылые, грызуны, зайцеобразные), кора развита более скромно и имеет гладкую поверхность.
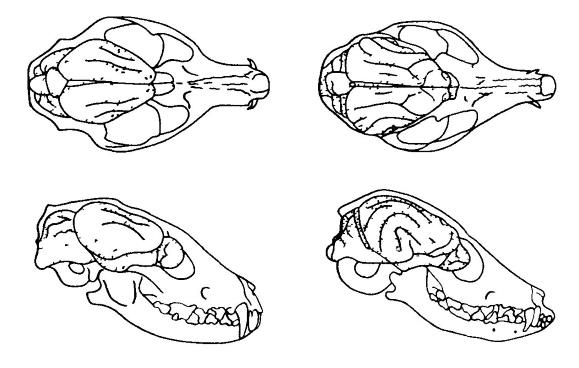
Расположение мозга в черепе у ископаемого и ныне живущего представителей псовых. Заметно увеличение размеров и усложнение мозга, в особенности полушарий переднего мозга. Геспероцион (Hesperocyon gregarius) (слева) — олигоценовая форма, жившая приблизительно 30 млн. лет назад. Фенек (Vulpes zerda) (справа) — современная форма сходных размеров.
Серое вещество новой коры характеризуется сложным гистологическим строением. У плацентарных млекопитающих выделяются 6 лежащих друг над другом слоев клеток и внедряющихся между ними волокон; это сильно отличает новой коры от сохранившихся участков коры палеопаллиума и архипаллиума, где можно различить лишь от 2 до 4 слоев клеток. Согласно существующим оценкам, у млекопитающих, имеющих особенно крупный мозг, число клеток новой коры может достигать миллиардов.
Белое вещество, расположенное под серым, помимо веера связей, идущих от коры к нижележащим отделам мозга и обратно, включает огромное количество переплетающихся поперечных волокон, соединяющих между собой различные области самой коры. Образовавшаяся таким образом комиссура растянута по направлению назад (согласно курсу разрастания полушарий) и разделена на две пластинки, слитые по заднему краю. Нижняя, более тонкая и отклоненная передним краем вниз — это свод (fornix), комиссура коры архипаллиума (т. е. гиппокампа). Верхняя, более толстая горизонтально расположенная комиссура принадлежит новой коре и именуется мозолистым телом (corpus callosum). Это образование позволяет объединить память обоих полушарий и значительно повышает способность мозга к обучению. Мозолистое тело имеется только у плацентарных в связи со значительным развитием новой коры, однопроходные и сумчатые его лишены. Кроме того, у всех млекопитающих имеется передняя комиссура (commissura anterior), соединяющая обонятельные участки коры.

Послойное расположение нервных клеток в коре конечного мозга млекопитающего.
Сложная система «проводников», связывающая все части коры, наводит на мысль о том, что серое вещество представляет собой в принципе единое образование, все части которого имеют одинаковые возможности для осуществления любых функций больших полушарий. В определенной степени так оно и есть: опыты показывают, что у лабораторных животных можно разрушить значительную часть новой коры, не вызвав стойких нарушений их нормальной активности. Данные об увечьях и болезненных изменениях подтверждают, что это справедливо и в отношении человеческого мозга. Вместе с тем ясно, что определенные участки коры в норме связаны с выполнением вполне конкретных функций. Выше были упомянуты участки палеопаллиума и архипаллиума, предназначенные в основном для анализа обонятельной информации и сохранившиеся соответственно в виде грушевидной доли и гиппокампа. Дифференцировка отдельных областей имеет место и в коре неопаллиума. Передняя часть полушарий содержит двигательную область. Находящаяся здесь лобная доля помимо прочего осуществляет управление общением животных, в том числе акустическим; у человека она связана с речью, т. е. второй сигнальной системой. Задняя часть полушарий связана с восприятием ощущений. В затылочной и височной долях находятся участки, управляющие соответственно зрением и слухом. Дальше вперед вблизи двигательной области располагаются участки, воспринимающие осязательные и проприоцептивные сигналы. У приматов центральная борозда (sulcus centralis), пересекающая вершину полушария от медиальной до боковой поверхности, отграничивает (хотя и не совсем точно) двигательную область от сенсорной. Вдоль переднего края центральной борозды в линейном порядке расположены специфические двигательные участки, обслуживающие каждую часть тела и конечностей. Вдоль заднего края центральной борозды в том же порядке размещаются участки сенсорного восприятия соответствующих частей тела.
Таким образом, у многих млекопитающих почти вся поверхность новой коры занята участками, более или менее тесно связанными с определенными чувствующими или двигательными функциями. Хотя центральная борозда может отсутствовать, у плацентарных в большинстве случаев соблюдается аналогичное линейное расположение сенсорных и двигательных участков друг против друга. У сумчатых (а среди плацентарных — у ксенартр) «разметка» участков тела примерно такая же, но сенсорные участки не отделены от двигательных, а перемежаются с ними. Но, например, у человека эти специфические функциональные участки занимают на поверхности новой коры относительно немного места. Между ними возникли обширные зоны серого вещества (один особенно крупный такой участок занимает большую часть лобной доли), которые не связаны с конкретными сенсорными или двигательными функциями. Поэтому такие области часто называют «белыми пятнами», хотя, как показывает повреждение этих участков, именно в них локализованы наши высшие умственные способности, включая возможности обучения, инициативу, умение предвидеть и способность к суждениям. Вместе с тем имеются и участки, которые могут быть удалены без серьезных последствий для интеллектуальной деятельности.
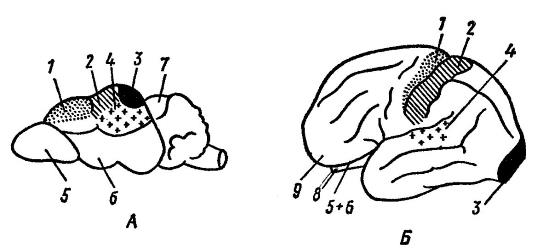
Функциональные центры коры головного мозга землеройки (Sorex sp.) (А) и человека (Homo sapiens) (Б): 1 — двигательный центр; 2 — центр кожно-мышечной чувствительности; 3 — зрительный центр; 4 — слуховой центр; 5 — обонятельная луковица; 6 — обонятельные доли; 7 — крыша среднего мозга; 8 — мозжечок; 9 — лобная доля.
На эволюцию головного мозга большое влияние оказывает внешняя среда и двигательная (пищедобывающая, оборонительная) деятельность. При этом развитие различных частей головного мозга определяется в основном способами отыскивания пищи: у собаки (Canis lupus), пользующейся в этом процессе обонянием, больше развита обонятельная область; у кошки (Felis silvestris), отыскивающей пищу с помощью зрения — зрительная; у макаки (Macaca mulatta), пользующейся зрением и слухом — зрительная и слуховая.
Обычно предполагается, что размеры больших полушарий обусловливают различия в умственных способностях разных млекопитающих. В определенном смысле это верно, но с существенными оговорками. Более крупный мозг состоит из большего числа нервных клеток. Если площадь имеющейся поверхности коры каким-либо образом связана с интеллектом, то очевидно, что из двух вариантов мозга одинаковых размеров более развитым будет тот, у которого поверхность бороздчатая, а менее развитым — мозг с гладкой поверхностью. Размеры самого животного тоже влияют на объем мозга. Это происходит уже хотя бы потому, что мозг должен иметь более обширные области для обслуживания более обширных сенсорных и двигательных связей. Однако увеличение размеров мозга происходит не полностью пропорционально массе тела, так что крупные животные демонстрируют тенденцию иметь относительно более мелкий мозг без всякого видимого ущерба для умственных способностей. Таким образом, абсолютные размеры головного мозга не являются безусловным критерием интеллекта. На это определенно указывает тот факт, что мозг кита может быть в пять раз больше по объему, чем мозг человека.
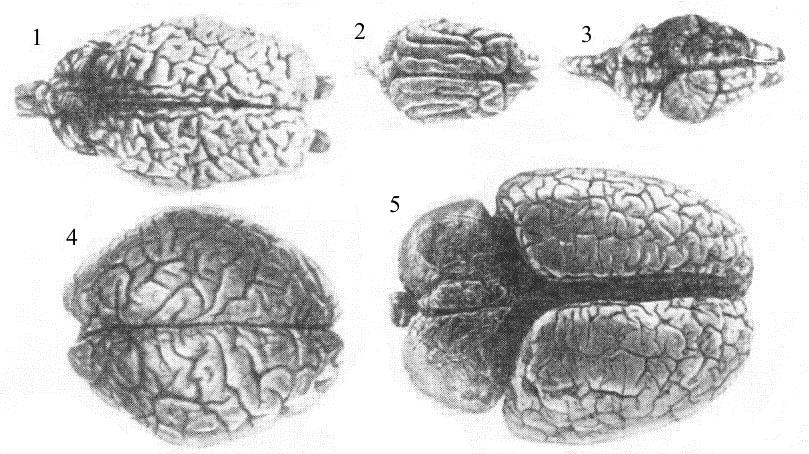
Сравнение головного мозга некоторых млекопитающих: 1 — лошадь; 2 — собака; 3 — кенгуру; 4 — человек; 5 — слон.
Процентная доля головного мозга в массе всего тела именуется индексом цефализации. У крупных насекомоядных он составляет около 0,6 %, у мелких — до 1,2 %, у крупных китообразных — порядка 0,3 %, а у мелких — до 1,7 %. У большинства приматов индекс цефализации составляет 1-2 % . У человека он достигает 2-3 %, а некоторые мелкие широконосые обезьяны имеют мозг, масса которого составляет до 7 % от массы тела. В то же время, у современных рептилий и птиц индекс цефализации колеблется в пределах от 0,05 до 0,5 %.
Ниже приводится масса головного мозга некоторых млекопитающих (в скобках указана масса животного):
виргинский опоссум (Didelphis virginiana) — 7,6 г (5 кг);
коала (Phascolarctos cinereus) — 19,2 г (8 кг);
саванный слон (Loxodonta africana) — 6000 г (5000 кг);
обыкновенный еж (Erinaceus europaeus) — 3,3 г (1 кг);
домовая мышь (Mus musculus) — 0,3 г (0,02 кг);
серая крыса (Rattus norvegicus) — 2 г (0,3 кг);
обыкновенная белка (Sciurus vulgaris) — 7 г (0,4 кг);
европейский кролик (Oryctolagus cuniculus) — 11 г (3 кг);
домашняя лошадь (Equus ferus) — 530 г (500 кг);
черный носорог (Diceros bicornis) — 500 г (1200 кг);
белохвостый олень (Odocoileus virginianus) — 500 г (200 кг);
жираф (Giraffa camelopardalis) — 680 г (800 кг);
домашняя овца (Ovis orientalis) — 140 г (55 кг);
домашний бык (Bos primigenius) — 490 г (700 кг);
двугорбый верблюд (Camelus bactrianus) — 762 г (700 кг);
гиппопотам (Hippopotamus amphibius) — 580 г (3500 кг);
дельфин-белобочка (Delphinus delphis) — 815 г (60 кг);
нарвал (Monodon monoceros) — 2997 г (1578 кг);
кашалот (Physeter macrocephalus) — 8028 г (35833 кг);
синий кит (Balaenoptera musculus) — 3636 г (50900 кг);
домашняя кошка (Felis silvestris) — 25 г (3 кг);
лев (Panthera leo) — 270 г (250 кг);
рыжая лисица (Vulpes vulpes) — 53 г (4,5 кг);
домашняя собака (Canis lupus) — 64 г (10 кг);
белый медведь (Ursus maritimus) — 500 г (700 кг);
морж (Odobenus rosmarus) — 1130 г (700 кг);
мармозетка Гельди (Callimico goeldii) — 7 г (0,2 кг);
белолобый капуцин (Cebus albifrons) — 57 г (1 кг);
макак-резус (Macaca mulatta) — 88 г (6,5 кг);
бабуин (Papio cynocephalus) — 200 г (25 кг);
серебристый гиббон (Hylobates moloch) — 112 г (6,5 кг);
калимантанский орангутан (Pongo pygmaeus) — 413 г (50 кг);
западная горилла (Gorilla gorilla) — 506 г (126 кг);
обыкновенный шимпанзе (Pan troglodytes) — 430 г (55 кг);
человек разумный (Homo sapiens) — 1400 г (72 кг).
Из приведенных примеров видно, что у более мелких млекопитающих мозг почти всегда относительно крупнее, а с возрастанием размеров тела животного относительная величина мозга уменьшается. Особенно отчетливо это проявляется среди близких видов млекопитающих — например, у кошки (Felis silvestris) и льва (Panthera leo). Весьма убедительны в этом смысле и собаки различных пород. Если массы тела самой мелкой и самой крупной пород находятся приблизительно в соотношении 1 : 33, то массы мозга у тех же пород соотносятся как 1 : 3.
У одомашненных животных, лишенных потребности добывать пищу и обороняться от врагов, размеры головного мозга значительно уменьшаются. Например, объем мозга волка (Canis lupus) на 30 % больше, чем у собаки одного с ним размера. Интересно, что эти изменения касаются не только традиционно домашних животных, но и представителей свободно живущих видов, какое-то время содержащихся в неволе. Так, лисицы (Vulpes vulpes), родившиеся в природе, но с первых же дней живущие в неволе, обладают меньшим мозгом, чем их сородичи, обитающие в естественных условиях. При этом различия достигают 20 %, что приблизительно соответствует разнице в объеме мозга диких и настоящих домашних животных. Уменьшение мозга, хотя и не столь выраженное (приблизительно на уровне 5 %), было обнаружено у содержащихся в неволе волков (Canis), хорьков (Mustela), крыс (Rattus). При этом уменьшение захватывает не все отделы мозга, а лишь те его области, которые связаны с работой органов чувств. Самое же примечательное, что у выпущенных на волю домашних животных вес мозга увеличивается. Например, у одичавших кошек мозг примерно на 10 % больше мозга их сородичей, живущих в домашних условиях. Значительное увеличение мозга было обнаружено также у одичавших кроликов (Oryctolagus cuniculus) на островах Кергелен. Одичавшие ослы (Equus asinus) в Южной Америке обладали на 15 % большим мозгом, чем домашние. Интересно также, что мозги неандертальца (Homo neanderthalensis) и палеолитического человека разумного (Homo sapiens) по своим размерам несколько превышали мозг современного человека.
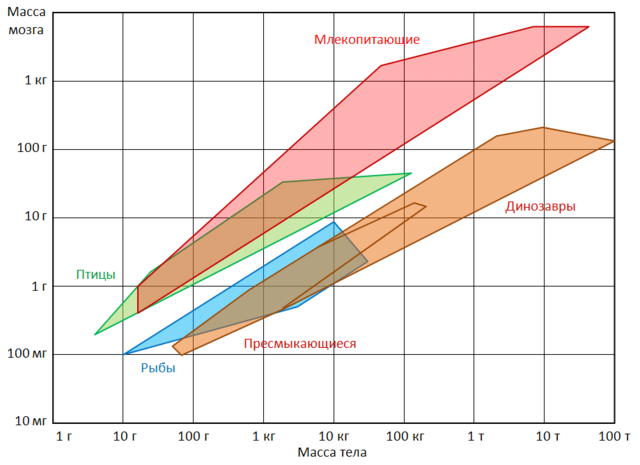
Области значений массы мозга и тела для некоторых групп позвоночных.
Установлено, что у многих млекопитающих наблюдается двигательная асимметрия, т. е. преимущественное использование правой или левой половины тела. Например, при изучении необъезженных лошадей (Equus ferus) фиксировали, с какой ноги животные начинают ходьбу, с какой стороны они предпочитают огибать препятствия и на каком боку предпочитают лежать в стойле на сене. В результате большинство кобылиц оказались правшами, а большинство жеребцов — левшами. Примерно 10 % лошадей не отдавали предпочтения ни правой, ни левой конечности. По наблюдениям, около 90 % моржей (Odobenus rosmarus) выкапывают моллюсков из морского ила правым ластом. Баюкая малышей, порядка 80 % самок шимпанзе (Pan) и горилл (Gorilla) прижимают их головками к левой стороне своей груди (примерно такое же процентное соотношение отмечается и среди женщин). Крысы (Rattus), ведущие поиск корма с помощью вибрисс, расположенных на правой стороне морды, являются более добычливыми, чем их сородичи-левши.
вперед
в оглавление