в оглавление
назад
Конечности
Скелет свободных конечностей построен в целом типично для тетрапод, но отражает характерную для млекопитающих позу. Их конечности не расставлены широко, как у амфибий и большинства рептилий, а подведены под туловище, располагаясь в вертикальной продольной (парасагиттальной) плоскости. Этой же плоскостью ограничены их изгибы и перемещения при шагающих движениях, что позволяет делать более длинный шаг и развивать большую скорость. Кроме того, увеличивается экономичность передвижения, поскольку все четыре конечности поддерживают тело без постоянного напряжения приводящей мускулатуры, неизбежного при прежнем, расставленном положении ног. Такой способ локомоции начал развиваться еще у прогрессивных зверообразных и остается относительно мало измененным у большинства наземных млекопитающих. Исключение составляют однопроходные и представители некоторых древних групп, сохраняющие «рептильную» постановку конечностей. Из плацентарных эту черту вторично приобрели кроты (Talpinae), что связано с их своеобразным способом продвижения под землей. Кроме того, ряд млекопитающих освоил двуногое передвижение, плавание и полет, что отразилось на особенностях устройства их конечностей.
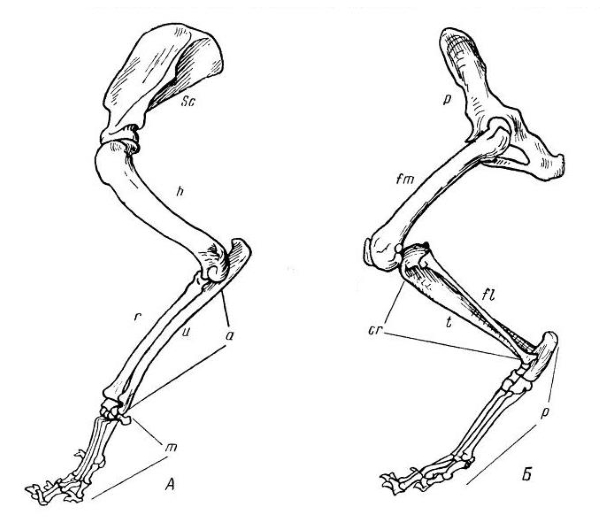
Передняя (А) и задняя (Б) конечности волка (Canis lupus): sс — лопатка (scapula); h — плечевая кость (humerus); а — предплечье (antebrachium); u — локтевая кость (ulna); r — лучевая кость (radius); m — кисть (manus); р — таз (pelvis); fm — бедренная кость (femur); cr — голень (crus); t — большая берцовая кость (tibia); fi — малая берцовая кость (fibula); р — стопа (pes).
Подобно амфибиям, в кистевом сгибе млекопитающих доминирует предплечно-запястное сочленение, а в пяточном — голеностопное сочленение. В то же время, для рептилий и птиц характерны межзапястное и межзаплюсневое сочленения. Кисть и стопа млекопитающих изначально относительно примитивны и несут все пять пальцев. Однако у большого числа видов в дальнейшем происходит их редукция или видоизменение. Чаще всего это касается первого пальца — во многих случаях он либо сильно уменьшается, либо совсем исчезает (например, у большинства копытных). Число пальцев даже может сократиться до одного (у продвинутых лошадей и литоптернов). Пальцы состоят из отдельных фаланг (phalanges digitorum). Дистальные, когтевые фаланги (phalanges unguales) могут быть сильно видоизменены и нести когти (исходный тип), ногти или копыта. В исходном варианте пятипалой конечности млекопитающих в I-м пальце имеется 2 фаланги, а в остальных — по 3. Таким образом, их фаланговая формула 2 - 3 - 3 - 3 - 3. У зверообразных она первоначально составляла 2 - 3 - 4 - 5 - 3, однако затем произошла редукция фаланг до состояния, характерного для млекопитающих. Несмотря на то, что часть пальцев у млекопитающих утрачивается, сокращение числа фаланг в сохранившихся пальцах происходит редко. Это наблюдается, например, у быстробегающих копытных. У китообразных фиксируется значительное увеличение числа фаланг в средних пальцах и уменьшение их числа в крайних пальцах передних конечностей. В первом случае диафизы и эпифизы фаланг окостеневают отдельно и образуют как бы самостоятельные фаланги. У представителей некоторых групп (сумчатых, насекомоядных, грызунов, хищных) в дистальных отделах конечностей нередки сесамоидные образования (преполлекс (prepollex) в кисти, прегаллюкс (prehallux) в стопе), которые иногда считаются рудиментами предпервого пальца.
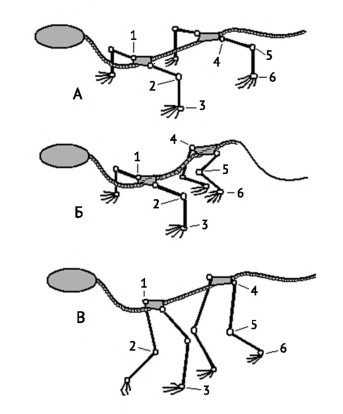
Схемы расположения конечностей относительно тела: А — типичная рептилия; Б — зверообразное; В — млекопитающее; 1 — плечевой сустав; 2 — локтевой сустав; 3 — лучезапястный сустав; 4 — тазобедренный сустав; 5 — коленный сустав; 6 — голеностопный сустав.
Передняя конечность состоит из трех отделов: плеча (humerus), предплечья (antebrachium) и кисти (manus).
В плечевом отделе имеется одна хорошо развитая плечевая кость (humerus), состоящая из центрального стержня и расширений на его концах. В проксимальной части и на стержне развиты пекторальный и дельтовидный гребни для прикрепления грудной, дельтовидной и подлопаточной мышц; последняя оканчивается на малом бугорке (tuberculum minus). Расположенный поблизости большой бугорок (tuberculum majus) предоставляет место для окончания других идущих от лопатки мышц — предостной и заостной. Кроме того, проксимальная часть плечевой кости обладает округлой головкой, входящей в суставную ямку лопатки. В дистальной части плечо несет на своей передневентральной поверхности округлый мыщелок для лучевой кости; постаксиальнее этого мыщелка на конце кости имеется выемка с блоковидной сочленовной поверхностью (trochlea) для локтевой кости. Дистальная часть плечевой кости расширена в обе стороны для крепления мышц предплечья. На переднем (наружном) крае находится наружный мыщелок (ectepicondylus), на котором начинаются разгибательные мышцы предплечья. Более крупный внутренний надмыщелок (entepilcondylus) располагается на заднем (внутреннем) крае и служит местом для начала сгибательных мышц. У многих примитивных млекопитающих имеется большое отверстие внутреннего надмыщелка (foramen entepicondylare) для нерва и кровеносных сосудов. У архаичных зверообразных также имелось отверстие наружного надмыщелка (foramen ectepilcondylare); у млекопитающих оно отсутствует. Короткое и массивное, со слабо выраженным стержнем у ранних зверообразных, плечо становится длиннее и тоньше у более прогрессивных представителей этой группы и у млекопитающих. В пределах последнего класса оно значительно отличается разнообразием своей длины и толщины. Так, плечевая кость вытянута и тонка у рукокрылых, но коротка и расширена у различных землероев.
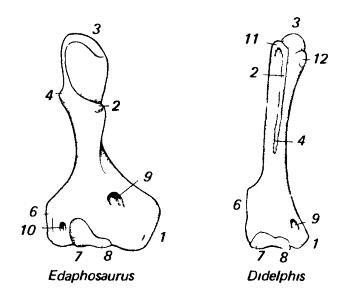
Плечевые кости зверообразного (пеликозавр эдафозавр (Edaphosaurus)) и млекопитающего (сумчатое опоссум (Didelphis)): 1 — внутренний надмыщелок; 2 — пекторальный гребень; 3 — головка; 4 — дельтовидный гребень; 6 — наружный надмыщелок; 7 — мыщелок для лучевой кости; 8 — мыщелок, или блок, для локтевой кости; 9 — отверстие внутреннего надмыщелка; 10 — отверстие наружного надмыщелка; 11 — большой бугорок; 12 — малый бугорок.
Предплечье состоит из расположенных параллельно лучевой (radius) и локтевой (ulna) костей. Толстая, колоннообразная лучевая кость поддерживает тело на кисти и сочленяется на своих верхнем и нижнем концах соответственно с плечом и запястьем. За исключением случаев крайней специализации к водному образу жизни, ее строение варьирует слабо. Верхняя фасетка лучевой кости сочленяется с плечом. Локтевая кость располагается латеральнее лучевой. Она редко принимает на себя сколько-нибудь значительную часть нагрузки, но важна для крепления мышц. Внизу она сочленяется с запястьем; наверху, в локте, посредством изогнутой выемки, примыкающей к фасетке лучевой кости, сочленяется с дистальным краем плеча. Над суставной выемкой локтевая кость продолжается в качестве массивного локтевого отростка (processus olecranon). За этот отросток тянется оканчивающаяся на нем мощная трехглавая мышца плеча, ответственная за разгибание (т. е. выпрямление) локтя. Отросток особенно велик у утконосов (Ornithorhynchidae), броненосцев (Dasypodidae), муравьедов (Myrmecophagidae), кротов (Talpidae) и некоторых других животных, прилагающих значительные усилия при рытье. В случае же быстробегающих млекопитающих (непарнопалых, парнопалых) ситуация с локтевым отростком прямо противоположна — конечность разгибается в значительной мере за счет силы тяжести.
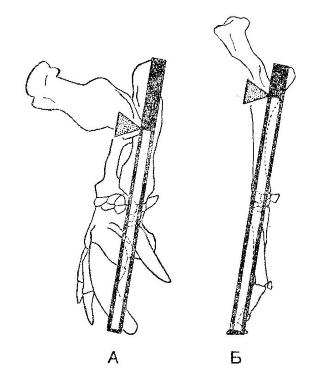
Относительная величина локтевого отростка у гигантского броненосца (Priodontes maximus) (А) и лошади (Equus ferus) (Б).
У многих млекопитающих лучевая и локтевая кости имеют приблизительно одинаковую длину и толщину. Так, они очень укорочены у китообразных и сильно вытянуты у ленивцев. Нередко дистальная часть и стержень локтевой кости срастаются с лучевой или полностью утрачиваются, но локтевой отросток имеется всегда. К примеру, у рукокрылых лучевая кость значительно больше и массивнее локтевой, при этом лучевая кость длиннее плечевой. Очень слаба локтевая кость у лошадей (Equidae) и жвачных (Ruminantia), хорошо развита у тапиров (Tapiridae), носорогов (Rhinocerotidae), бегемотов (Hippopotamidae) и свиней (Suidae). Иногда (например, у человека) развивается способность лучевой кости вместе с кистью вращаться вокруг локтевой, так что рука из положения ладонью вверх и вперед (при параллельном положении костей предплечья) — супинация — может переходить в расположение ладонью вниз и назад (с перекрестом локтевой кости лучевой) — пронация.
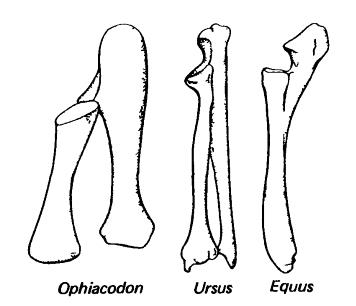
Левые лучевая и локтевая кости при виде с разгибательной (дорсальной, переднелатеральной) стороны у зверообразныого (пеликозавр офиакодон (Ophiacodon)) и млекопитающих (плацентарные медведь (Ursus) и лошадь (Equus)).
Кисть разделяется на проксимальный отдел — запястье (carpus), промежуточный — пясть (metacarpus) и дистальный — пальцы (digiti).
Запястье представляет собой гибкий посредник между предплечьем и пальцами. У млекопитающих оно состоит из 3 рядов элементов:
— 3 косточки, находящиеся непосредственно дистальнее костей предплечья: ладьевидная кисти (scaphoideum), полулунная (lunare) и клиновидная (cuneiforme);
— 1 центральная косточка (centralе), вклинивающаяся между предыдущим и последующим рядом (у ранних тетрапод их было 4, но затем количество сократилось);
— 5 карпальных костей (carpalia), каждая из которых лежит напротив одного из пястных элементов.
Также имеется добавочная маленькая гороховидная кость (pisiforme), прикрепленная к наружному краю запястья и служащая местом прикрепления сухожилия мышцы, идущей вдоль этой стороны конечности.
Помимо индивидуальных названий элементов запястья, используемых только в отношении млекопитающих, имеются их наименования, общие для всех тетрапод. Кроме того, в медицинской школе некоторым из этих элементов присвоены еще и другие названия, непригодные в иных случаях. Ниже приводятся общие названия костей запястья и их синонимы:
radiale, scaphoideum, naviculare manus — лучевая запястья, или ладьевидная кисти;
intermedium, lunare, semilunare, lunatum — промежуточная, или полулунная;
ulnare, carpi ulnare, cuneiforme, triquetrum, pyramidale — локтевая запястья, или клиновидная, либо трехгранная;
centrale, mediale — центральная кисти;
pisiforme, pisiforme — гороховидная;
carpale distale 1, trapezium, multangulum majus, cubiforme — большая многоугольная, или кубическая;
carpale distale 2, trapezoideum, multangulum minus — малая многоугольная, или трапециевидная;
carpale distale 3, magnum, capitatum — большая, или головчатая;
carpale distale 4(4+5), unciforme, hamatum, uncinatum — крючковидная.
Для многих млекопитающих характерна редукция и срастание между собой отдельных запястных элементов. К примеру, наличие центральной кости является архаичным признаком. Она имеется у грызунов и китообразных, многих ксенартр, но отсутствует или слаборазвита у большинства копытных (за исключением даманов (Procaviidae), тапиров (Tapiridae), свиней (Suidae)); также присутствует у молодых слонов, у которых позднее сливается с ладьевидной костью. Также широко распространена тенденция к утрате V-й карпальной кости (зачастую она срастается с IV-й); это происходит даже тогда, когда V-й палец сохраняется, а редукция пальцев обычно влечет дальнейшие утраты.
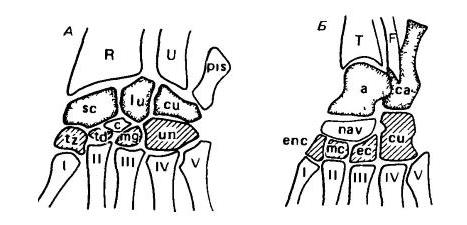
Схема запястья (А) и предплюсны (Б) млекопитающих. Проксимальный ряд элементов выделен точками, центральный ряд (и гороховидная кость) оставлен белым, дистальный ряд заштрихован. Пястные и плюсневые элементы обозначены римскими цифрами. R — лучевая кость (radius); U — локтевая кость (ulna); sc — ладьевидная кость (scaphoideum); lu — полулунная кость (lunare); сu — клиновидная кость (cuneiforme); c — центральная кость (centrale); pis — гороховидная кость (pisiforme); tz — многоугольная кость (trapezium); td — трапециевидная кость (trapezoideum); mg — большая кость (magnum); un — крючковидная кость (unciforme); Т — большая берцовая кость (tibia); F — малая берцовая кость (fibula); а — таранная кость (astragalus); са — пяточная кость (calcaneum); еnс — I-я предплюсневая кость (entocuneiforme); nav — центральная кость стопы (naviculare); mс — II-я предплюсневая кость (mesocuneiforme); ес — III-я предплюсневая кость (ectocuneiforme); сu — IV-я предплюсневая кость (cuboideum).
В типичной пятипалой конечности количество пястных костей (metacarpalia) соответствует числу пальцев, однако у некоторых млекопитающих оно может отличаться вследствие редукции некоторых пястных элементов или их слияния между собой. Например, у парнопалых кости пясти часто срастаются в одну кость, называемую cannon. С другой стороны, даже в случае полной редукции пальцев могут сохраняться рудиментарные пястные элементы. У млекопитающих, специализированных в быстром беге, особенно у копытных, может происходить значительное удлинение пястных элементов сохраняющихся пальцев; тем самым в конечности прибавляется дополнительный функциональный сегмент. За пястью находятся свободные пальцы кисти. Начиная с I-го (внутреннего), они носят следующие наименования: большой (digitus pollex), указательный (digitus index), средний (digitus medius), безымянный (digitus annularis) и мизинец (digitus minimus).
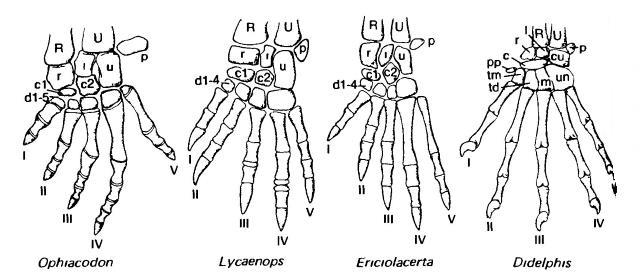
Кисть зверообразных (пеликозавр офиакодон (Ophiacodon), горгонопс лиценопс (Lycaenops), тероцефал эрициолацерта (Ericiolacerta)) и млекопитающего (сумчатое опоссум (Didelphis)). В запястье происходит утрата V-го дистального элемента и уменьшение числа центральных с 2 до 1; в дистальной части утрачиваются "лишние" фаланги III-го и IV-го пальцев. R — лучевая кость (radius); U — локтевая кость (ulna); сu — клиновидная кость (cuneiforme); l — полулунная кость (lunare); m — большая кость (magnum); p — гороховидная кость (pisiforme); r — ладьевидная кость (radiale); td — трапециевидная кость (trapezoideum); tm — многоугольная кость (trapezium); c, c1, c2 — центральные кости (centralia); Рр — преполлекс (prepollex); d1-5 — дистальные кости запястья (carpalia distalia).
Задняя конечность также имеет 3 отдела: бедро (femur), голень (crus) и ступню (pes). У большинства млекопитающих бедро короче голени.
Бедренный отдел содержит одну мощную бедренную кость (femur) цилиндрической формы с расширенными концами. Проксимальная часть бедра снабжена более или менее сферической головкой (capitulum), которая обращена внутрь и входит в вертлужную впадину таза. Также в проксимальной части находятся 2-3 вертлуга (trochanteres) — специальные выступы для прикрепления мышц. На верхнем конце бедра развивается большой вертлуг (trochanter major), на котором оканчиваются глубокие ягодичные мышцы, идущие от подвздошной кости. На вентральной поверхности дистальнее головки находится свойственный только для млекопитающих малый вертлуг (trochanter minor), на котором оканчивается подвздошнопоясничная мышца. С противоположной стороны проксимальной части бедра расположен III-й вертлуг, на котором оканчивается большая ягодичная мышца. Характерный для примитивных тетрапод IV-й вертлуг (trосhаnter quartus) отсутствует, поскольку прикреплявшиеся к нему сильные мышцы, идущие от хвоста к бедру, у млекопитающих уменьшились в размерах или исчезли. Вдоль стержня бедра вентрально проходит аддукторный гребень для прикрепления мощных приводящих мышц; иногда гребень редуцируется, принимая вид шероховатых линий (linea aspera). Дистально бедро расширяется и имеет на обоих боках шероховатости, от которых начинаются мышцы голени; вдоль середины дорсальной поверхности в дистальной части проходит глубокий желоб для сухожилия разгибателей колена, которое тянется к большой берцовой кости. Дистальный конец несет широкую двойную сочленовную поверхность для головки большой берцовой кости и рядом — на заднем (латеральном, постаксиальном) крае бедра — поверхность для причленения малой берцовой кости.
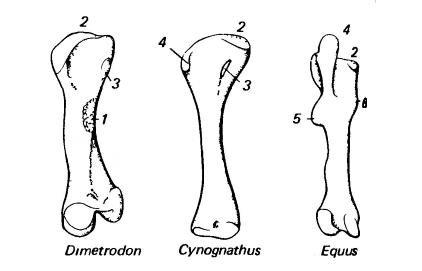
Левые бедренные кости (вид с вентральной стороны) зверообразных (пеликозавр диметродон (Dimetrodon), цинодонт циногнат (Cynognathus)) и млекопитающего (плацентарное лошадь (Equus)). Проксимальный конец наверху; дистально расположены сочленовные поверхности для большой берцовой кости. 1 — IV-й вертлуг; 2 — головка; 3 — внутренний вертлуг; 4 — большой вертлуг; 5 — III-й вертлуг; 6 — малый вертлуг.
Голень состоит из большой и малой берцовых костей.
Большая берцовая кость (tibia), соответствующая лучевой в передней конечности, является главным опорным элементом голени; она всегда лучше развита, сочленяясь дистально на большом протяжении с внутренней частью предплюсны, а проксимально посредством сильно расширенной треугольной в сечении головки почти со всей вентральной поверхностью дистального конца бедра. Передний край головки несет мощный кнемиальный гребень (crista cnemialis), к которому прикрепляется конечное сухожилие крупных мышц-разгибателей колена. Первично сухожилие прикреплялось к нему непосредственно. Однако по пути сухожилию приходится круто перегибаться через конец бедра, поэтому у млекопитающих в этом месте обычно развивается крупная сесамовидная кость — коленная чашка (patella), которая ездит поверх коленного сустава и по функциям сравнима с локтевым отростком в передней конечности. Сухожилие разгибателей оканчивается на этом окостенении, которое в свою очередь связано с кнемиальным гребнем.
Малая берцовая кость (fibula) в некоторых отношениях сравнима с локтевой в предплечье; она несет небольшую нагрузку, в связи с чем подвержена редукции. Поскольку в задней конечности разгибательное сухожилие прикрепляется к большой берцовой кости, малая не образует ничего подобного локтевому отростку. Малая берцовая кость сочленяется проксимально с задней (наружной) стороной конца бедра, а дистально — с наружной частью предплюсны. У многих млекопитающих малая берцовая кость утратила свое сочленение с бедром; сочленение с предплюсной тоже может сильно редуцироваться или даже утрачиваться. Обычно эта кость сохраняется в качестве тонкого элемента, но иногда происходит срастание ее концов с большой берцовой костью или утрата ее стержня.
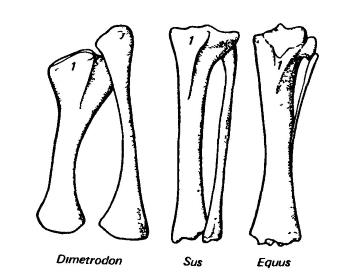
Левые большая и малая берцовые кости (вид с разгибательной, или дорсальной, стороны) зверообразного (пеликозавр диметродон (Dimetrodon)) и млекопитающих (плацентарные свинья (Sus) и лошадь (Equus)). 1 — кнемиальный гребень.
Стопа образована предплюсной (tarsus), плюсной (metatarsus) и пальцами (digiti).
Проксимальный ряд предплюсны состоит из 2 костей. С внутренней стороны стопы расположена таранная кость (astragalus), которая является продуктом слияния редуцированной большеберцовой кости предплюсны (tibiale), промежуточной (intermedium) и центральной стопы (centrale). У таранной кости развивается головка в виде поверхности вращения с килями, которые входят в соответствующие желобки в дистальном конце большой берцовой кости. Сустав является шарниром, практически полностью исключающим вращение, но допускающим большой размах разгибания и сгибания. Для парнопалых характерна таранная кость, у которой нижний конец тоже имеет поверхность вращения с килями, так что при беге возможен чрезвычайно широкий размах переднезадних движений стопы. Расположенная снаружи стопы пяточная кость (calcaneum) в лучшем случае имеет небольшое сочленение с более проксимальными элементами конечности, но образует направленный назад мощный пяточный бугор, к которому прикрепляются посредством ахиллова сухожилия икроножные мышцы. Дистальная часть предплюсны содержит 5 предплюсневых элементов (tarsalia).
Для обозначения костей предплюсны у млекопитающих, как и для обозначения костей запястья, существуют сложные системы названий, которые приводятся ниже:
tibiale + intermedium + centrale proximale, astragalus, talus — малоберцовая предплюсны, или таранная;
fibulare, calcaneum, calcaneus, calcis — пяточная;
centrale distale, naviculare, naviculare pedis — центральная стопы, или ладьевидная стопы;
tarsale distale 1, entocuneiforme, cuneiforme mediale (primum) — I-я предплюсневая, или I-я клиновидная;
tarsale distale 2, mesocuneiforme, cuneiforme intermedium (secundum) — II-я предплюсневая, или II-я клиновидная;
tarsale distale 3, ectocuneiforme, cuneiforme laterale (tertium) — III-я предплюсневая, или III-я клиновидная;
tarsale distale 4(4+5), cuboideum — IV-я предплюсневая, или кубовидная.
Плюсна образована 5-6 плюсневыми костями (metatarsalia). I-й палец ступни называют большим (digitus hallux), остальные 4 именуются по порядковым номерам.
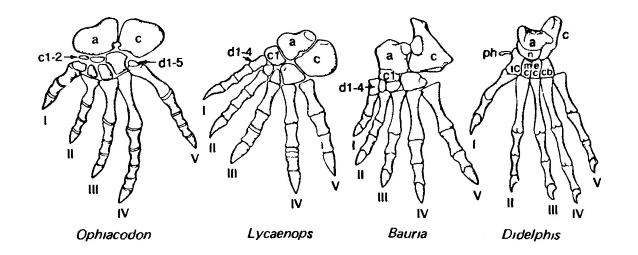
Стопа зверообразных (пеликозавр офиакодон (Ophiacodon), горгонопс лиценопс (Lycaenops), тероцефал баурия (Bauria)) и млекопитающего (сумчатое опоссум (Didelphis)). Принципиальные изменения включают развитие блоковидной поверхности на таранной кости, развитие бугра на пяточной кости, утрату двух элементов предплюсны и редукцию числа фаланг. а — таранная кость (аstragalus); с — пяточная кость (calcaneum); с1, с2 — центральные кости стопы (centralia); n — ладьевидная кость стопы (naviculare); d1-d5 — дистальные предплюсневые кости (tarsalia distalia); cb — IV-я предплюсневая кость (cuboideum); еc — III-я предплюсневая кость (ectocuneiforme); iс — I-я предплюсневая кость (entocuneiforme); mc — II-я предплюсневая кость (mesocuneiforme); ph — прегаллюкс (prehallux) (редкий у млекопитающих).
Таким образом, для млекопитающих характерны приведенные к телу трехзвенные Z-образные конечности, верхний сгиб которых обращен вперед, а нижний — назад. За пределами этого класса подобная конфигурация встречается только у динозавров и птиц. В задней конечности верхний сгиб образован коленным суставом, но в передней конечности гомологичное сочленение, т. е. локтевой сустав, по независимым причинам обращен назад и действует в качестве нижнего сгиба. Роль верхнего сгиба, аналогичного колену, досталась плечевому суставу, что потребовало выполнения некоторых морфофункциональных условий. Свобода этого сустава (подобно коленному) по отношению к осевому скелету достигнута полной редукцией коракоида (а у хороших бегунов — и ключицы), а на роль верхнего звена в схему передней конечности включена лопатка. В задней конечности, где голень, гомологичная предплечью, образует среднее звено, нижнего звена исходно не хватало; оно сформировано за счет стопы с пяточным бугром и вытянутыми плюсневыми костями. Кости пясти также вытянуты, но кисть не образует самостоятельного рычага, а примыкает к предплечью в составе нижнего звена.
Плечо и бедро типичного млекопитающего двигаются в передне-задней плоскости, действуя как рычаг с точкой опоры (колено или локоть) на одном конце, с грузом близ другого конца у сочленения с поясом и с главной силой (мышцы), приложенной у самой верхушки (большой бугорок плеча и большой вертлуг бедра). Поэтому главные мышцы являются по своему положению дорзальными, вследствие чего пояса конечностей редуцируются в своей вентральной части и увеличиваются в верхней (что особенно резко выражено в плечевом поясе).
В связи с приспособлением к различным условиям существования и со специализацией в совершении тех или иных движений, конечности многих млекопитающих претерпели специфические изменения: изменилась относительная длина их отделов, конфигурация и толщина костей, число пальцев и т. п.
вперед
в оглавление