в оглавление
назад
Конечности (продолжение)
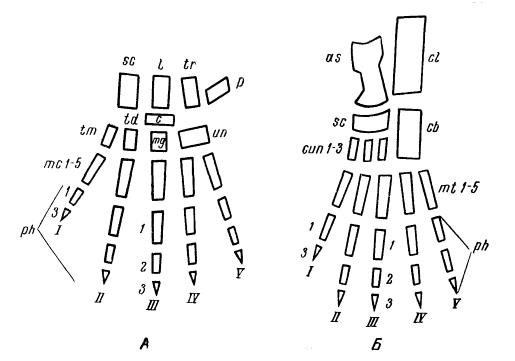
Схемы исходного строения кисти (А) и стопы (Б) млекопитающих. Кости: sс — ладьевидная (naviculare); l — полулунная (lunatum); tr — трехгранная (triquetrum); р — гороховидная (pisiforme); с — центральная (centrale); tm — многоугольная (trapezium); td — трапециевидная (trapezoideum); mg — большая (magnum): un — крючковатая (unciforme); сl — пяточная (calcaneus); аs — таранная (astragalus); cun 1-3 — I-III-и клиновидные (cuneiforme 1-3); сb — кубовидная (cuboideum); mc — пястные (metacarpalia); mt 1-5 — I-V-и плюсневые (metatarsalia); ph 1-3 — I-III-я фаланги пальцев (phalanges); I-V — I-V-е пальцы.
У примитивных форм ноги короткие, достаточно сильно согнутые в суставах, их распластанная по субстрату ступня несет 5 широко расставленных пальцев. Такие конечности еще не специализированны в каком либо одном направлении и способны к разнообразным движениям: к умеренно быстрому бегу, к схватыванию пищи и поднесению ее ко рту, к плаванию, лазанью по деревьям и т. д. В связи с этим их суставы относительно свободны, допускают движение в разных направлениях: сгибание и разгибание, отведение в сторону и приведение к телу, вращение вокруг продольной оси и комбинацию всех этих движений. Соответственно устроены и суставные поверхности. Головка плечевой кости довольно сильно выпукла, бедренной — почти шарообразна, лучевая и локтевая свободно вращаются одна относительно другой вокруг продольной оси. В предплюсне верхняя поверхность таранной кости и, соответственно, нижняя — большой берцовой слаборельефны, почти плоские; в нижнем ряду — 4 косточки. Для запястья у наиболее примитивных форм характерно сохранение полного числа костей; раньше других исчезает свободная центральная кость, у большинства млекопитающих во взрослом состоянии сращенная с ладьевидной. Суставные поверхности костей запястья рельефно-выпуклые, вогнутые, седловидные, что, в связи со значительным рельефом суставов костей пясти и фаланг пальцев, обеспечивает необходимую свободу движении кисти и относительную независимость отдельных пальцев; в стопе эта свобода выражена несколько меньше. Пальцы конечностей немногим отличаются по величине друг от друга; короче всех I-й, длиннее других — III-й, а иногда и IV-й. Такой тип конечностей обычно свойствен небольшим наземным животным с всеядным режимом питания, например, ежам (Erinaceus) и крысам (Rattus). Сходные конечности имели и древнейшие млекопитающие; немногим отличаются они у медведей (Ursus) и барсуков (Meles), у которых, однако, ладьевидная кость уже срослась с полулунной.
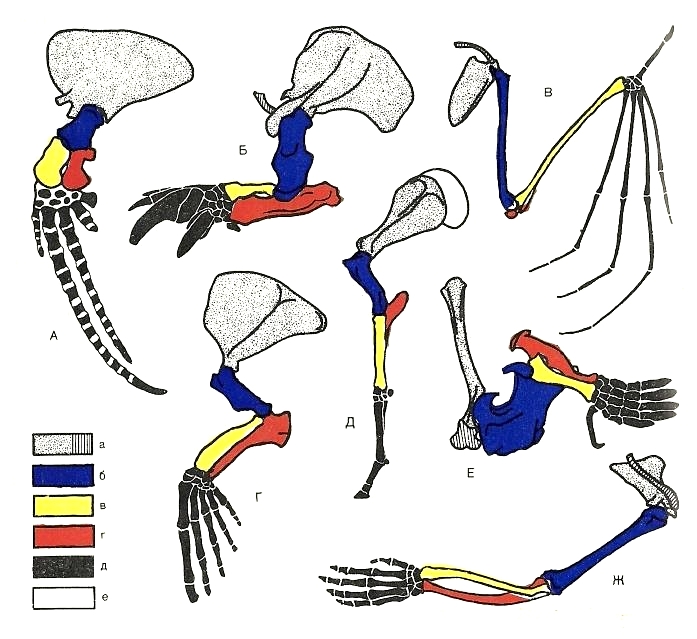
Схематическое изображение строения левой передней конечности различных млекопитающих: А — лопатка и ключица; б — плечевая кость; в — лучевая кость; г — локтевая кость; д — кисть; А — финвал (Balaenoptera physalus); Б — гигантский броненосец (Priodontes maximus); В — рыжая вечерница (Nyctalus noctula); Г — сивуч (Eumetopias jubatus); Д — лошадь (Equus ferus); Е — обыкновенный крот (Talpa europaea); Ж — западная горилла (Gorilla gorilla).
Для ранних и значительной части современных млекопитающих (большинства насекомоядных, приматов, медведей (Ursidae), енотов (Procyonidae), некоторых других хищных и грызунов) при передвижении свойствен упор на всю кисть и стопу, т. е. стопохождение. Такое положение ступни оказалось весьма полезным для лазания, ведь один из способов фиксации на деревьях — охватывание ветвей кистью и стопой, которые распластываются по их поверхности. Соответственно, адаптация к древесному образу жизни в большинстве случаев способствовала сохранению данного признака.
У некоторых насекомоядных, части грызунов и многих хищных приподнятые над землей запястье, пясть, предплюсна и плюсна стали более или менее вертикальными продолжениями голени и предплечья, а конечности опираются только на пальцы — наблюдается пальцехождение. При этом I-е пальцы, как более короткие, перестают достигать земли, а в дальнейшем могут совсем исчезать — на задних конечностях раньше, чем на передних. Специализация такого рода весьма характерна для хищников, перешедших к питанию теплокровной добычей, в погоне за которой необходим быстрый бег или прыжки. Начальный этап подобной специализации прослеживается у некоторых гиенодонтов (Hyaenodonta), в более ясной форме она проявляется у собак (Canidae) и части кошек (Felidae), в особенности у гепарда (Acinonyx jubatus). Ограничение подвижности кисти проявляется при этом в слиянии некоторых костей запястья (ладьевидной с полулунной и центральной). В стопе ограничение подвижности проявляется в образовании более глубокой борозды блока таранной кости и в ослаблении рельефа суставных поверхностей других элементов.
Иногда между стопохождением и пальцехождением выделяется промежуточный тип положения ступни — полустопохождение, которое свойственно, например, некоторым виверрам (Viverridae) и куницам (Mustelidae). Муравьеды (Myrmecophagidae) при ходьбе подгибают длинные когти и опираются на тыльную сторону ступней; подобным же образом передвигались вымершие наземные ленивцы. Для крупных человекообразных обезьян (Hominidae) и халикотериев (Chalicotheriinae) характерна опора на костяшки подогнутых пальцев передних конечностей.
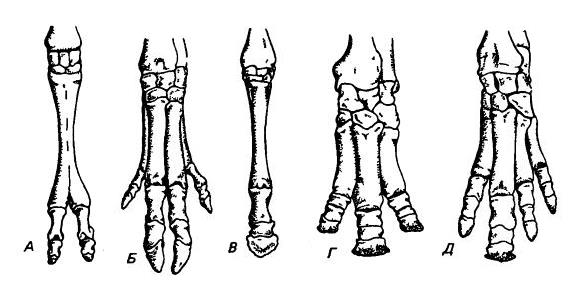
Левая кисть двугорбого верблюда (Camelus bactrianus) (А), кабана (Sus scrofa) (Б), лошади (Equus ferus) (В), индийского носорога (Rhinoceros unicornis) (Г) и чепрачного тапира (Tapirus indicus) (Д). Верблюд и свинья относятся к парнопалым, у этих животных ось симметрии проходит между III-м и IV-м пальцами. У свиньи боковые пальцы, II-й и V-й, представлены полностью, но малы. У большинства парнопалых, как и у верблюда, два главных элемента пясти срослись в os cannon. Три остальных формы — непарнопалые, у которых ось проходит через III-й палец. У тапира I-й палец утрачен, но остальные четыре пальца сохраняются. У современных носорогов V-й палец исчез. У современных лошадей II-й и IV-й пальцы редуцированы до состояния грифельных косточек.
Жесткие конечности с малой опорной площадкой более экономно передают энергию при резком толчке и потому сопутствуют адаптации к быстрому бегу. Наиболее полного выражения это достигает у копытных, особенно населяющих открытые ландшафты, для которых бег служит главной защитой от хищников. Конечности, преимущественно кисть и стопа, у этих бегунов еще сильнее удлиняются; не только запястье и предплюсна, но и фаланги пальцев, за исключением конечных с развитыми на них копытами, отвесно поднимаются от земли — животные становятся фалангоходящими. Опора тела все больше концентрируется на средних пальцах — III-м (у непарнопалых), или III-м и IV-м (у парнопалых). В первом случае ось конечности проходит через опорный палец, во втором — между этими пальцами. Ширина таких пальцев увеличивается, остальные пальцы делаются значительно тоньше, а при крайней специализации перестают касаться земли и могут совсем исчезнуть (у лошадей (Equidae), верблюдов (Camelidae), полорогих (Bovidae) и некоторых других). Примечательно, что в большинстве случаев редукция пальцев происходит быстрее в задней конечности. Вместе с тем сужаются и удлиняются пясть и плюсна, а средние пястные и плюсневые кости нередко срастаются в одну, чем достигается большая прочность этого звена и большая устойчивость (у тушканчиков (Dipodidae), большинства жвачных (Ruminantia)).
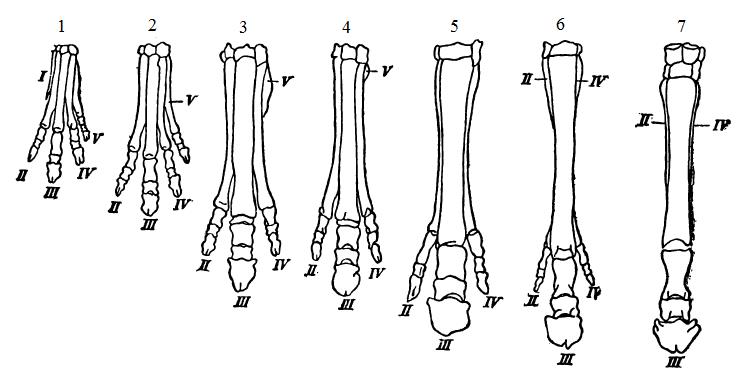
Филогенетическое развитие передней конечности лошадей; пальцы пронумерованы римскими цифрами: 1 — эогиппус (Eohippus pernix); 2 — орогиппус (Orohippus agilis); 3 — мезогиппус (Mesohippus celer); 4 — миогиппус (Miohippus anceps); 5 — гипогиппус (Hypohippus equinus); 6 — неогиппарион (Neohipparion whitneyi); 7 — протогиппус (Protohippus pernix).
В суставах, по возможности, исключаются все движения, кроме маятникообразных спереди назад, в связи с чем изменяется форма их поверхностей. Головка плечевой кости уплощается, бедренной — приближается по форме к поперечноцилиндрической; лучевая и локтевая кости утрачивают взаимную подвижность, причем локтевая становится тонкой, а при крайней специализации от нее сохраняются только верхний и нижний концы, вполне срастающиеся с лучевой (лошади, верблюды). Сильно утоньшается и малая берцовая кость, в крайних случаях от нее также сохраняются только концы, прирастающие к большой берцовой или свободные. В суставе между плечом и предплечьем развиваются направленные спереди назад гребни и борозды, взаимно вклинивающиеся друг в друга и исключающие боковые сдвиги и вращение. Косточки в запястье и предплюсне, относящиеся к уменьшенным или утраченным боковым пальцам, исчезают или срастаются с соседними; косточки двух рядов взаимно вклиниваются друг в друга, а верхние — также во впадины на нижней поверхности лучевой кости; таранная кость приобретает глубокую борозду верхнего блока, соответствующую высокому гребню большой берцовой кости. Все это препятствует боковым сдвигам в суставах. При этом верхние и нижние поверхности косточек часто теряют рельеф, уплощаются, особенно в предплюсне, где может также происходить их частичное слияние (например, ладьевидной с кубовидной у большей части жвачных).
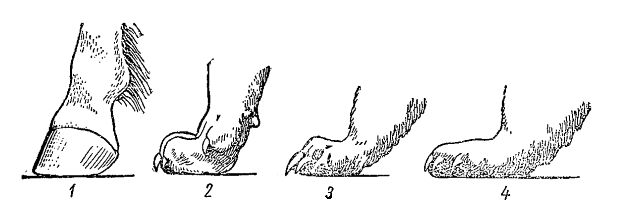
Положение стопы у различных млекопитающих: 1 — фалангохождение (лошадь (Equus ferus)); 2 — пальцехождение (волк (Canis lupus)); 3 — полустопохождение (лесная куница (Martes martes)) (по другим данным — пальцехождение); 4 — стопохождение (бурый медведь (Ursus arctos)).
Итак, в рассмотренном ряду происходит относительное удлинение конечностей, и в первую очередь их дистальных отделов — кисти и стопы. Это не только увеличивает длину шага, но и способствует повышению скорости. Если стопохождение является относительно медленным типом передвижения, пальцехождение и особенно фалангохождение дают возможность очень быстрого бега. С конечностями бегательного типа значительное сходство имеют конечности прыгающих животных, особенно задние (кенгуру (Macropodidae), тушканчики (Dipodidae)). Параллельно удлинению конечностей редуцируется или полностью исчезает ключица, и тем самым плечо освобождается от всякого костного соединения с туловищем, что ослабляет сотрясение тела при ударе передних ног о землю после прыжка. Первоначально гибкий скелет становится приспособленным почти исключительно к передне-заднему движению конечностей, так что другие их движения оказываются неловкими или даже невозможными.
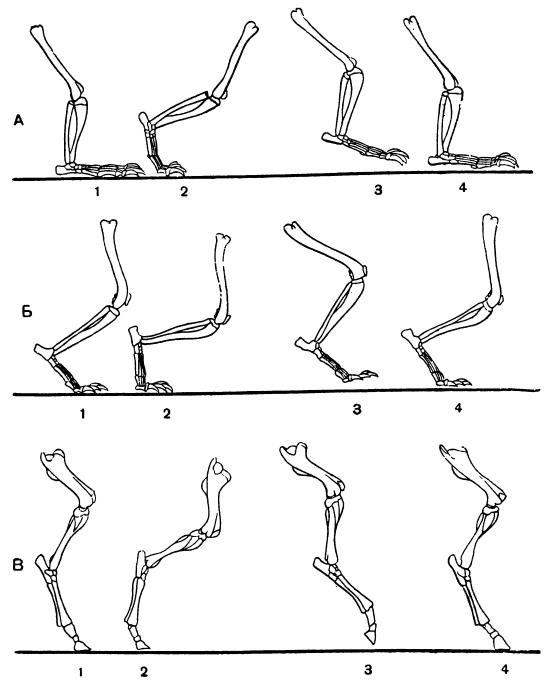
Скелет конечностей стопоходящих (А), пальцеходящих (Б) и фалангоходящих (В) млекопитающих и схематическое изображение четырех последовательных фаз (1-4) их движения при ходьбе.
В конечностях очень крупных наземных зверей (слоны (Elephantidae), индрикотерии (Indricotheriinae), некоторые бронтотерии (Brontotheriidae) и др.) имеются приспособления к поддержанию большой тяжести тела. Во избежание лишней траты мускульной энергии при стоянии или при разгибании в суставах конечности крупных животных становятся столбообразными, слабо согнутыми в суставах (тяжесть тела поддерживается преимущественно самими костями, а не мышцами), что отражается на строении всех костей. Локтевая кость, на которую при отвесном положении конечностей приходится основное давление тяжести тела, делается более массивной; лучевая — иногда тоньше локтевой (у слонов); кисть и стопа большей частью короткие и широкие (исключение — индрикотерии с удлиненной кистью и стопой, что имело особое биологическое значение); в суставах значительно ограничена свобода движений.
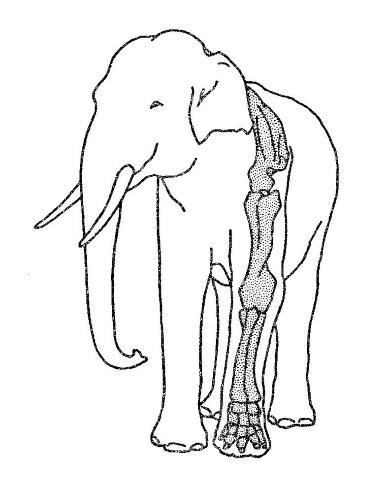
Скелет левой передней конечности азиатского слона (Elephas maximus), врисованный в контуры тела.
Слоны могут показаться стопоходящими, но на самом деле они пальцеходящи, поскольку земли у них касаются лишь пальцы, а остальные костные элементы кисти и стопы приподняты. Пальцы расходятся в стороны, охватывая крупную волокнистую подушку и формируя широкую округлую опорную площадку. Она ограничивает погружение конечности при движении по топкому грунту и, что очень важно, облегчает ее отрывание, поскольку сужается при поднимании, впуская в лунку воздух. Даже при самом скоростном передвижении слонов хотя бы одна из их конечностей сохраняет контакт с субстратом. Быстрый бег невозможен, но в нем и нет необходимости: огромные животные защищены от врагов уже своим ростом, а часто, кроме того, и разного рода вооружением.
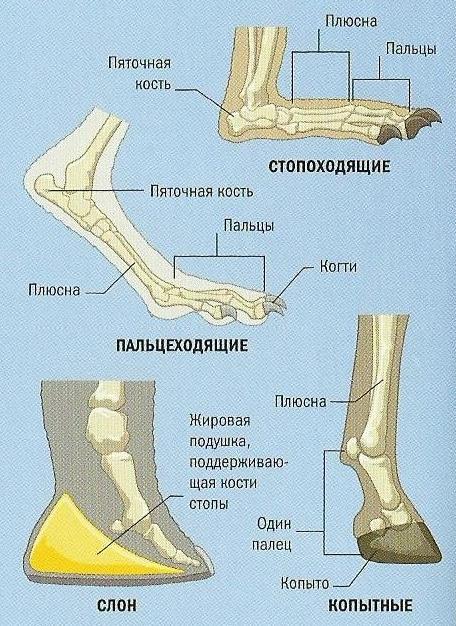
Постановка ступни слона в сравнении с другими млекопитающими.
У роющих млекопитающих, вскапывающих и отбрасывающих землю при помощи передних конечностей (некоторые броненосцы, насекомоядные и грызуны), кости сохраняют примитивную подвижность в суставах и соответственно изменены. Прежде всего, они сильно расширены. Гребни и бугры, служащие для прикрепления мышц-сги6ателей плечевого сустава и кисти и разгибателей локтевого сустава, разрастаются; в частности, значительно удлиняется локтевой отросток локтевой кости (плечо силы разгибателей). Наибольшего развития достигают передние конечности кротов. Они направлены в стороны и характеризуются максимальным укорочением и расширением всех отделов. Необходимую опору в корпусе конечность получает на переднем членике грудины (рукоятке) через посредство толстой короткой ключицы, конструктивно уподобившейся коракоиду. Плечевая кость обладает мощными мышечными отростками и дополнительным сочленением верхнего конца с ключицей, чем достигается большая крепость плечевого сустава. Локтевой сгиб направлен вверх, роющую лопату образуют короткое предплечье и пятипалая кисть, обращенная ладонью вбок. Кисть получает дополнительное увеличение роющей поверхности путем образования серповидной кости (falciforme). Огромные конечные фаланги продольно расщеплены на конце для более прочного укрепления в них роговых когтей. Конечности скребут и отгребают грунт мощными боковыми усилиями.
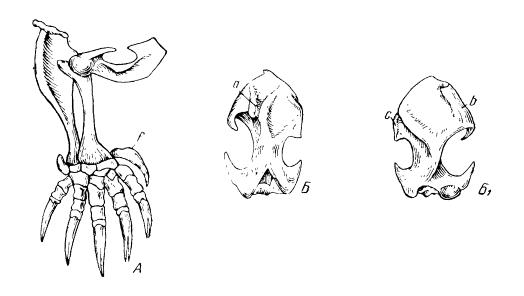
Передняя конечность обыкновенного крота (Таlpa europaea): А — передняя конечность; Б — плечевая кость сзади, Б1 — плечевая кость спереди; f — серповидная кость (falciforme): а — головка; b — дополнительное сочленение с ключицей; c — малый бугорок.
Своеобразное приспособление для лазанья по деревьям — способность I-го пальца в той или иной мере противопоставляться остальным — развивается в конечностях большей части приматов, некоторых сумчатых и грызунов (здесь необходимо заметить, что отставленный I-й палец встречается и у наземных форм). При сохранении свободы движений в большинстве суставов, сочленение между многоугольной и I-ой пястной костью, а иногда — и между I-й плюсневой и клиновидной костями, становится седловидным. Это дает возможность I-му пальцу кисти, а порой — и стопы, не только приближаться к другим и отклоняться от них (приведение и отведение) и не только сгибаться и разгибаться, но и подводиться под остальные пальцы, поворачиваясь к ним своей нижней (ладонной или подошвенной) стороной. Последующее сгибание подведенного таким образом пальца, вместе со сгибанием остальных, дает возможность крепко охватывать предметы, в частности ветви деревьев.
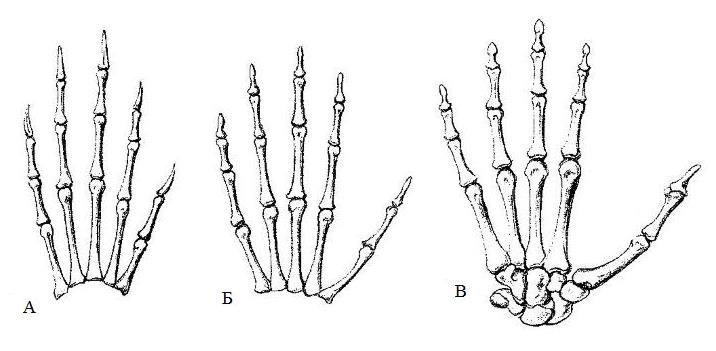
Противопоставление I-го пальца на кисти тупайи (А), капуцина (Б) и человека (В).
Приспособление конечностей к полету в полной мере выражено только у рукокрылых. Кости их передних конечностей очень вытянутые и тонкие. Поскольку движения при полете производятся в основном сгибанием и разгибанием в плечевом суставе, подвижность остальных суставов ослаблена; в связи с этим локтевая кость редуцируется до верхнего и нижнего концов, более или менее неподвижно сращенных с лучевой, а кости первого ряда запястья сращены между собой, кроме трехгранной (иногда), и с центральной. Только I-й палец передних конечностей развит нормально. Пястные кости и фаланги пальцев с II-го по V-й, поддерживающие кожистую летательную перепонку, очень длинные и тонкие. В связи с утратой способности к ходьбе, когтевые фаланги на них исчезают, кроме II-го пальца (в некоторых случаях), сохраняясь лишь в форме хряща. Верхний конец малой берцовой кости также исчезает; для поддержки заднего отдела летательной перепонки на пяточной кости развивается длинная шпора.
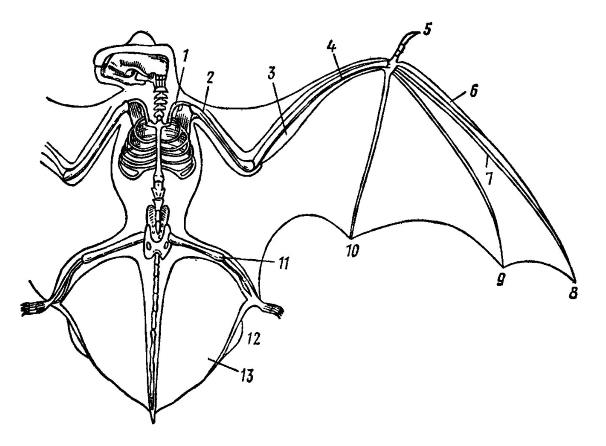
Скелет летучей мыши, врисованный в контуры тела: 1 — ключица; 2 — плечевая кость; 3 — лучевая кость; 4 — локтевая кость; 5 — I-й палец; 6 — II-й палец; 7 — III-й палец; 8 — концевая фаланга III-го пальца; 9 — IV-й палец; 10 — V-й палец; 11 — бедренная кость; 12 — шпора пяточной кости; 13 — тазобедренная перепонка.
При приспособлении к плаванию конечности превращаются в ласты; их пальцы охвачены общей кожистой плавательной перепонкой, нередко опускающейся дальше концов пальцев. В остальном приспособление к плаванию идет разными путями. У ластоногих конечности служат органом продвижения вперед в воде, а отчасти (особенно передние) — и для ползания по суше. В связи с этим плечо и бедро укорачиваются, а пясть, плюсна и пальцы удлиняются. При этом конечности достаточно подвижны в суставах, что обеспечивает разнообразные движения при плавании и передвижении по суше. Менее всего подвижны суставы фаланг пальцев, фиксированных в вытянутом и раздвинутом состоянии для поддержки плавательной перепонки.
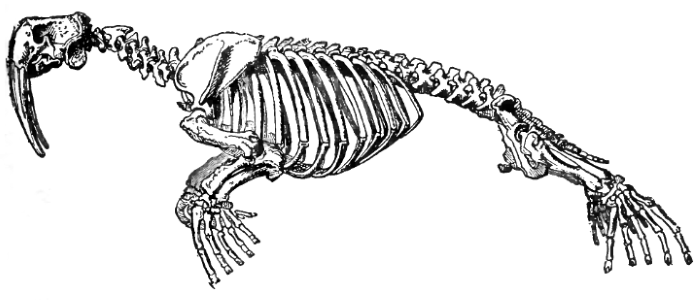
Скелет моржа (Odobenus rosmarus).
К Современные китообразные и сирены перешли к полностью водному образу жизни и совершенно утратили способность выходить на сушу. Их передние конечности преобразованы в ласты, а задние совершенно редуцированы (их закладка появляется на ранней эмбриональной стадии, но вскоре исчезает). Перемещение в толще воды происходит с помощью горизонтальных движений мощного хвоста, не имеющего в своем составе никаких скелетных частей. У вымерших представителей обеих групп, стоящих более близко к наземным предшественникам, для плавания использовались все четыре конечности. Сегодня передние конечности китообразных служат органами равновесия и поворота. Они сохраняют подвижность только в основном суставе, остальные суставы теряют рельеф своих сочленяющихся поверхностей и срастаются при помощи хряща. Плечо и предплечье резко укорочены и расширены, а пальцы нередко удлинены путем увеличения числа фаланг (гиперфалангия), порой очень значительного — до 14 фаланг в одном пальце; в этом отношении китообразные сходны с вымершими морскими рептилиями. Сирены с помощью своих менее специализированных передних конечностей не только управляют движением, но и ползают по дну, обнимают детенышей, зажимают обеими кистями водные растения и подносят их ко рту. Ласты водных форм перемещаются относительно тела в основном как единое целое, перемещение же их отделов относительно друг друга развито слабо.
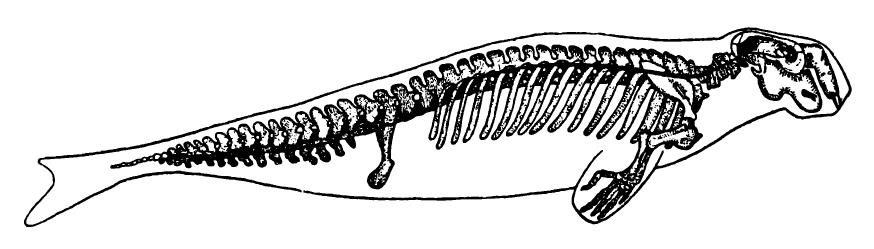
Скелет дюгоня (Dugong dugong), врисованный в контуры тела.
Кроме описанных основных типов приспособлений конечностей и их, часто существенных, видоизменений, имеются и другие (например, к подвешиванию на ветвях деревьев и др.). Характер движения животного, даже при незначительном отличии от движения близких форм, кладет отпечаток на все кости конечностей; кроме того, их строение отражает путь, пройденный предками в филогенетическом развитии. Можно сказать, что у каждого рода млекопитающих все кости конечностей, а особенно кисти и стопы, отличаются от костей любого другого рода. В частности, — очень показательно строение таранной и пяточной кости.
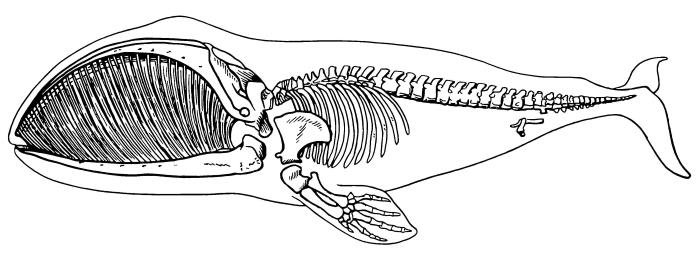
Скелет (и китовый ус) гренландского кита (Balaenа mysticetus), врисованный в контуры тела.
Как и каждая техническая конструкция, скелет обладает определенным резервом прочности, т. е. как бы имеет коэффициент безопасности, обеспечивающий в экстремальных ситуациях выдерживать нагрузки, значительно превосходящие обычные. Если масса спокойно стоящего животного равномерно распределена на все четыре конечности, то при усилении нагрузки, например при беге или когда приходится тащить тяжелую жертву, вся тяжесть тела переносится на одну конечность. Зарегистрированы случаи, когда лев (Panthera leo) тащил ношу в 250-300 кг, а пума (Puma concolor) — тушу бычка весом в 270 кг. Коэффициент безопасности должен быть рассчитан на подобные напряжения костей во что бы то ни стало. Так, бедренная кость человека (Homo sapiens) в состоянии покоя нагружена массой приблизительно 35 кг, но известно, что она ломается лишь при нагрузке около 600 кг. Бедренная кость мыши (Mus musculus) выдерживает нагрузку, в 750 раз большую, чем в состоянии покоя, бедренная кость суслика (Spermophilus citellus) — в 325 раз, слона (Elephas maximus) — в 120 раз. Из приведенных цифр понятно, что у крупных животных коэффициент безопасности значительно ниже, чем у мелких. В значительной мере это обусловливается тем, что кости многих крупных млекопитающих пневматизируются. Облегчение костей снижает резерв, который дает им возможность выдерживать увеличивающиеся нагрузки. И кости конечностей носорога (Rhinoceros unicornis), например, уже не могут выдержать нагрузки, возникающие при скачках, а конечности слона разрешают лишь быструю рысь как самый скорый для него способ передвижения.
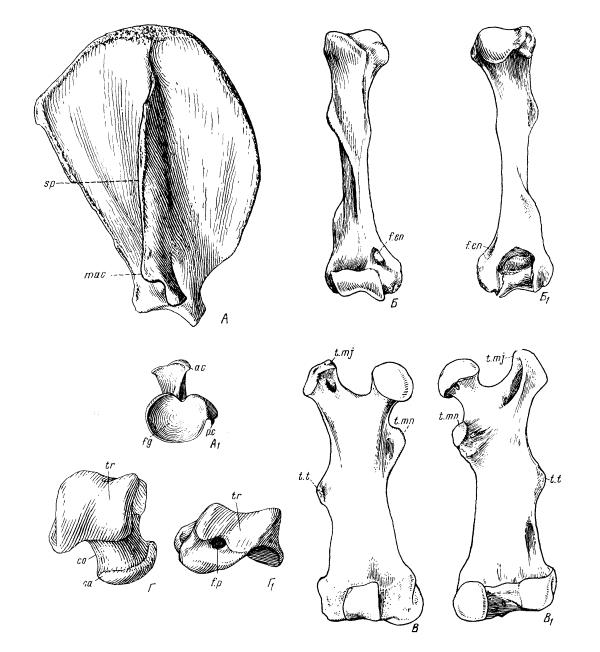
Элементы строения некоторых костей. Лопатка кошки (Felis silvestris) снаружи (А) и снизу (Аl); плечевая кость барсука (Meles meles) спереди (Б) и сзади (Бl); бедренная кость бобра (Casfor fiber) спереди (В) и сзади (Вl); таранная кость смилодона (Smilodon fatalis) сверху (Г) и сзади (Г1). ас — акромион; mас — метакромион; р. с — клювовидный отросток; f. g — суставная впадина; sp — ость лопатки; f. еn — внутреннее надмыщелковое отверстие; t. mj — большой вертлуг бедренной кости; t. mn — малый вертлуг бедренной кости; t. t — III-й вертлуг бедренной кости; tr — блок таранной кости; со — шейка таранной кости; са — головка таранной кости, f. р — заднее отверстие таранной кости.
Гетеротропные элементы
Помимо обыкновенных хрящей и костей, которые объединены в общую скелетную конструкцию, имеются разнообразные кости и хрящи, которые не являются частью собственно скелета, а появляются в качестве вспомогательных элементов. Обычно они замещают волокнистую соединительную ткань, присутствующую в данном месте у других форм; кость может формироваться прямо из такой ткани или проходить промежуточную хрящевую стадию. У млекопитающих маленькие кости и хрящи часто образуются в сухожилиях — в точках, где возможно трение, например там, где сухожилие перекидывается через костный гребень или сустав. Такие структуры, часто встречающиеся в кисти и стопе, называются сесамовидными, или сесамоидами (sesamoideum). Коленная чашка, или надколенник (patella) является «переросшим» сесамоидом. Примечательно, что у антилопы канны (Taurotragus oryx) во время ходьбы чашечки издают громкие щелкающие звуки, которые люди слышат на расстоянии до 100 м. Кости, которые разбросаны в других местах и представляются позднейшими приобретениями, именуются гетеротопными костями.
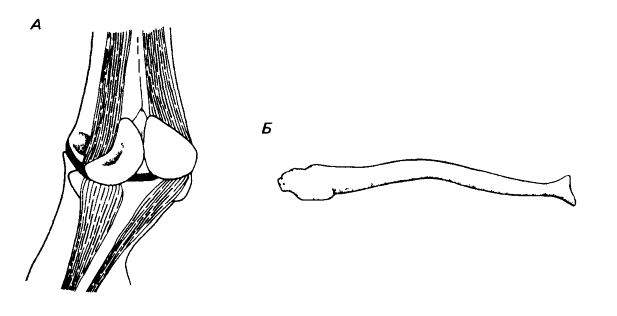
Гетеротропные кости. А — 2 сесамовидные кости на задней стороне сустава между пястным элементов (наверху) и проксимальной фалангой (внизу) у лошади (Equus ferus); штрихами показаны сухожилия, идущие к сесамовидным костям и от них. Б — бакулюм выдры (Lutra lutra); проксимальный конец обращен влево.
В половом члене самцов ряда групп млекопитающих обнаруживается специальная поддерживающая кость — бакулюм (baculum). У самок в пределах тех же систематических групп его заменяет баубеллюм (baubellum), поддерживающий клитор. Прочие гетеротопные элементы имеют более ограниченное распространение. Так, у панд (Ailuropodinae) и малых панд (Ailuridae) сесамовидная косточка пясти — преполлекс — образует «VI-й», или ложный, палец. Он способен сгибаться и разгибаться согласованно с настоящим большим пальцем, функционируя наподобие клещей. Пандам этот добавочный палец позволяет крепко обхватывать бамбуковые стебли во время кормежки, а малым пандам помимо этого помогает достигать относительно тонких ветвей. У кротов (Talpidae) разросшийся преполлекс, именуемый серповидной костью (falciforme), служит для увеличения площади роющей поверхности кисти. На конце морды некоторых млекопитающих обнаруживаются ростральные кости (rostralia): пятачковая косточка у свиней (Suidae) и пекари (Tayassuidae), хоботковая — кротов (Talpidae) и прыгунчиков (Macroscelidea). У парашютирующих форм присутствует специальная кость (osrea patagii), служащая для растягивания летательной кожной складки; у летяг (Petauristinae) она примыкает к кистевому сгибу, у шипохвостов (Anomaluridae) — к локтевому сгибу. У летучих мышей (Microchiroptera) имеется костная шпора, поддерживающая межбедренную летательную перепонку. В перегородке сердца оленей (Cervidae), полорогих (Bovidae) и верблюдов (Camelidae) сформированы маленькие сердечные косточки (cordis), предоставляющие дополнительную опору напряженно работающим мышцам и клапанам сердца. Имеются и другие подобные примеры.
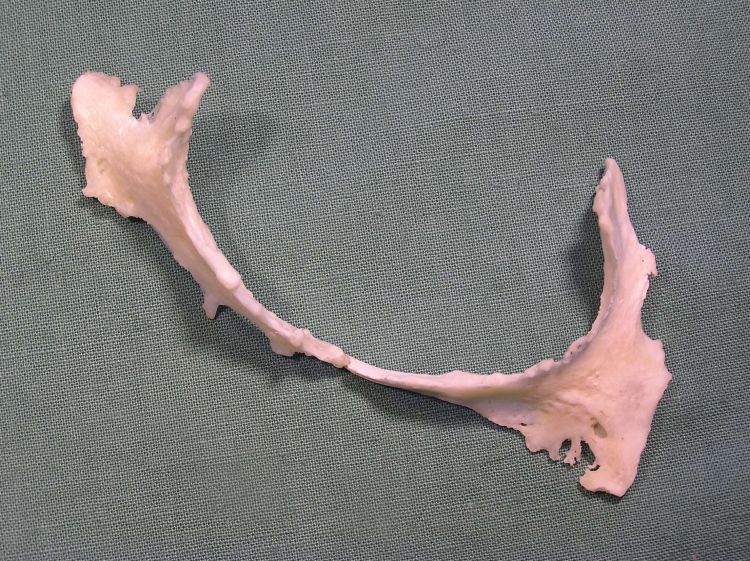
Окостенение из сердечной перегородки крупного рогатого скота (Bos primigenius taurus).
Кожные окостенения
Остеодермы представляют собой вторичные окостенения, развивающиеся в мезодермальном слое кожи. Весьма характерные для многих рептилий, среди млекопитающих они присутствуют только у ксенартр. У броненосцев развит костный панцирь, а у многих наземных ленивцев имелись внутрикожные окостенения, играющие роль органов пассивной защиты.

К окостенениям дермы относятся и т. н. кожные кости (cornua), которые прирастают к лобным костям и формируют костную основу рогов жвачных (Ruminantia). У полорогих (Bovidae) и жирафов (Giraffidae) они постоянные. У оленей (Cervidae) кожные кости сменяемые; ежегодно они отпадают из-за разрушения их оснований остеокластами, после чего кожа смыкается над местом прикрепления отпавшего рога и под кожей развивается новая кость более сложной формы.

Смонтированный скелет и часть костного панциря девятипоясного броненосца (Dasypus novemcinctus). Панцирь состоит из костяных пластинок, покрытых роговыми щитками. На спине пластинки образуют подвижные пояса, обеспечивающие панцирю эластичность.
вперед
в оглавление