в оглавление
назад
Мышечная система (продолжение)
Скелетная мускулатура млекопитающих устроена гораздо сложнее, чем у других позвоночных животных. Она высоко дифференцирована и характеризуется большим числом разнообразно расположенных мускулов, выполняющих самые разные функции. У зверя средних размеров насчитывается 200-250 различных мышц. Движения тела и его частей редко выполняются за счет сокращения единственного мышечного элемента; обычно взаимодействует несколько мышц, силы которых складываются. Таким образом, функция отдельной мышцы зависит от того, в составе какой группы она действует. Мышцы работают либо в одном направлении (тогда они синергисты), либо в противоположном (антагонисты). Противодействующие группы мышц дополняют друг друга. Их синхронное действие под строгим контролем центральной нервной системы обеспечивает широкий спектр сложных движений челюстей, головы и шеи, аппарата дыхания, туловища и конечностей, особенно в связи с быстрым передвижением по земле и с древесным образом жизни. Мускулатура у представителей разных отрядов и семейств существенно отличается в зависимости от характерных способов передвижения. За исключением водных форм, наибольшего развития она достигает в области конечностей и их поясов.
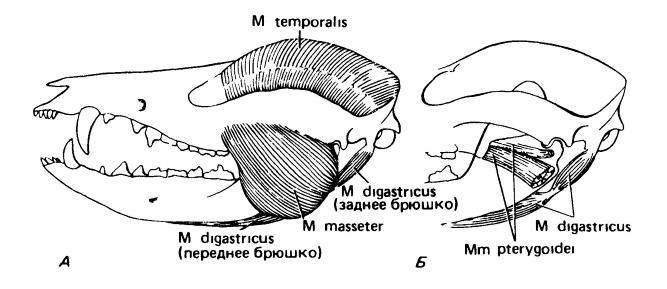
Челюстная мускулатура виргинского опоссума (Didelphis virginiana): А — вид с поверхности; Б — более глубокий слой. На Б челюсть изображена как бы прозрачной, чтобы были видны мышцы, прикрепляющиеся к ее внутренней поверхности.
У млекопитающих очень хорошо развита жевательная мускулатура, связанная с захватом и механической обработкой пищи и достигающая особенно сильной дифференцировки у плотоядных и растительноядных форм. Опускается нижняя челюсть, помимо тяжести самой кости, при помощи челюстно-подъязычной мышцы (m. mylohyoideus), отделившейся от межчелюстной мышцы, и комплексной двубрюшной мышцы (m. digastricus), составленной из мускулатуры 2 висцеральных сегментов. Переднее брюшко образовано волокнами, идущими назад под ветвью нижней челюсти. Позади оно соединяется с пучком мускулатуры, который идет вверх позади нижней челюсти и прикрепляется к черепу около уха. 2 брюшка лежат под углом друг к другу, но вместе с челюстно-подъязычной мышцей успешно выполняют не слишком трудную задачу размыкания челюстей. Обе этих мышцы иннервируются тройничным (V-м черепным) нервом и отчасти принадлежат системе лицевого (VII-го) нерва.
Из мускулатуры подъязычной дуги, иннервируемой VII-м черепным нервом, у низших тетрапод развивается мускульный листок — сжиматель шеи (m. sphincter coli). У млекопитающих он распадается на подкожную мышцу шеи (platysma mypides) и мускульный комплекс, распространившийся на значительную часть поверхности головы и образовавший подкожный пласт лицевых, или мимических мышц. Эти мышечные пучки, хотя и распределены на значительной площади, как правило, особенно сконцентрированы вокруг глазниц, наружного уха, вибрисс, ноздрей и губ (в случае, если они развиты). Соответствующими мышцами приводятся в движение веки, ушные раковины, нос, хоботок многих насекомоядных. Более 40 000 мышц насчитывается в хоботе слонов. Ротовой сфинктер делает возможным акт сосания молока детенышем. Также мышцы лица обеспечивают мимику, играя важную роль в общении и передаче информации, что особенно важно для хищных и приматов. По-видимому, лицевые мышцы появились на ранних этапах эволюции млекопитающих, однако зверообразные их еще не имели; у однопроходных они принципиально отличаются от мышц сумчатых и плацентарных. Комплекс особых мышц обслуживает и подвижный язык млекопитающих. Мышцы лица иннервируются VII-м черепным нервом, языка — XII-м.
Лицевая мускулатура виргинского опоссума (Didelphis virginiana).
Группа находящихся в глазнице маленьких мышц обеспечивает движения глазного яблока. Сокращение их в различных сочетаниях поворачивает глаз в любом нужном направлении. 4 прямые мышцы начинаются сзади, рядом с глазным стебельком или зрительным нервом. 2 косые мышцы отходят от передней части глазницы. Верхняя прямая (m. rectus superior) и верхняя косая (m. obliquus superior) мышцы прикрепляются к верхнему краю глазного яблока, пара нижних мышц (m. rectus inferior, m. obliquus inferior) — к его нижнему краю, а 2 оставшиеся прямые мышцы — к переднему, или внутреннему (m. rectus аnterior), и заднему, или наружному (m. rectus posterior), краям. Прямые и нижняя косая мышцы сохраняются у млекопитающих в том же виде, как у нижестоящих форм, тогда как верхняя косая мышца своим прикреплением уходит вглубь глазницы и перекидывается к месту окончания у края глазницы через блок (trochlea), которому обязан своим названием иннервирующий ее тонкий нерв. 4 из 6 указанных мышц иннервируются III-м черепным нервом, верхняя косая — IV-м (блоковым, nervus trochlearis), а задняя прямая — VI-м. Кроме указанных мышц, у большинства млекопитающих (исключение составляют приматы) имеется ретрактор глазного яблока (m. retractor bulbi), втягивающий его в глазную впадину и обслуживаемый VI-м черепным нервом. От верхней прямой мышцы обособляется подниматель верхнего века (m. levator palpebrae superioris), также иннервируемый III-м черепным нервом. Пучки ретрактора закрывают глаз мигательной перепонкой (в случае ее наличия).
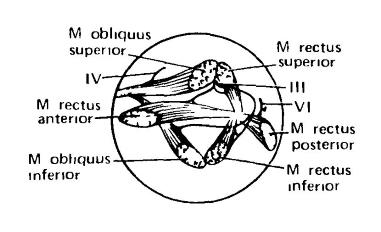
Глазные мышцы млекопитающего. Вид сбоку при удаленном глазном яблоке (обозначено окружностью); овалы — это окончания мышц на глазном яблоке. Показаны 3 черепных нерва (III-й, IV-й, VI-й), идущие к глазным мышцам. Передний конец обращен влево.
Мышцы системы языкоглоточного (IX-го черепного) и блуждающего (X-го черепного) нервов. т. е. мускулы жаберного аппарата, сохраняются лишь в виде мышц подъязычного и глотательного аппаратов. Помимо грудино-подъязычной (m. sternohyoideus) и лопаточно-подъязычной (m. omohyoideus) имеется еще несколько мышц языка — подбородочно-язычная (m. genioglossus) для выдвигания и подъязычно-язычная (m. hyoglossus) для втягивания языка. Обе язычных мышцы обслуживаются XII-м черепным нервом.
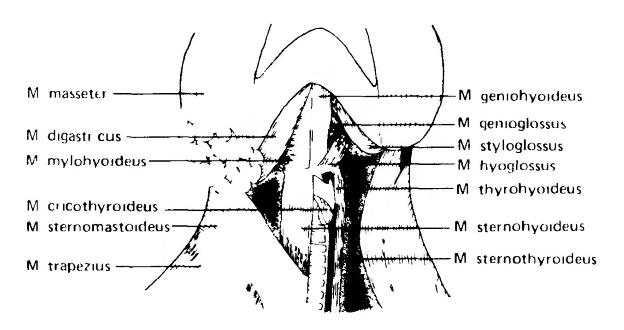
Мышцы горла и шеи домашней кошки (Felis silvestris catus). На правой стороне рисунка поверхностные мышцы и другие образования удалены, чтобы были видны более глубокие слои
Спинная мускулатура развита хорошо. Многие мышцы ограничены областью позвоночника, исполняя задачу его сгибания и фиксации в определенном положении. Вдоль спины над поперечными отростками позвонков тянется главная поверхностная мышца в дорсальной группе — длиннейшая мышца спины (m. longissimus dorsi). Более медиально, между ней и верхними остистыми отростками, находятся сложно перекрещивающиеся пучки мелких мышц, которые связывают следующие друг за другом верхние остистые и поперечные отростки — остистая и полуостистая мышца спины (m. spinalis dorsi и m. semispinalis dorsi) и др. Вместе они составляют систему поперечно-остистой мышцы (m. transversospinalis). От поперечных отростков к ребрам идут поперечно-реберные мышцы (mm. transversocostales). На протяжении поясничного отдела 2 более латеральные части мускулатуры могут объединяться в сильную крестцово-остистую мышцу (m. sacrospinalis), которая помогает разгибать позвоночник. В задней части туловища имеется подвздошно-реберная мышца (m. ileocostalis), которая идет от подвздошной кости к латеральной стороне ребер, простираясь вниз поверх мышц боков туловища. Наконец, присутствует хвостовая система мышц.
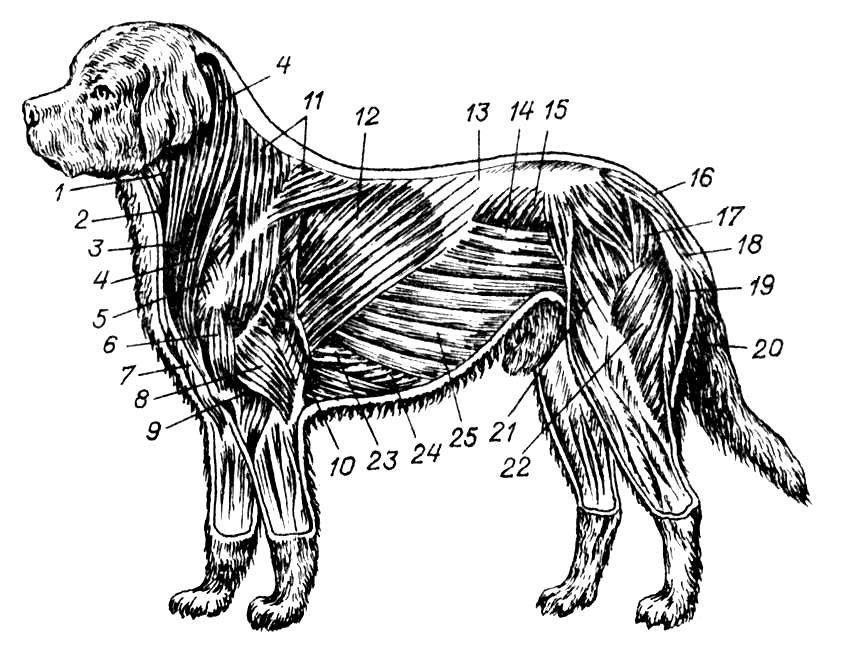
Поверхностная мускулатура домашней собаки (Canis lupus familiaris): 1 — грудино-щитовидная мышца; 2 — грудино-подъязычная мышца; 3 — грудино-головная мышца; 4 — IV-я плечеголовная мышца; 5 — плечеатлантная мышца; 6 — дельтовидная мышца; 8 — латеральная головка трехглавой мышцы плеча; 9 — плечевая (внутренняя) мышца; 10 — длинная головка трехглавой мышцы плеча; 11 — трапециевидная мышца; 12 — широчайшая мышца спины; 13 — ее пластинчатое сухожилие; 14 — косая брюшная внутренняя мышца; 15 — пояснично-спинная фасция; 16 — ягодичная средняя мышца; 17 — ягодичная поверхностная мышца; 18 — хвостовая мышца; 19 — полуперепончатая мышца; 20 — полусухожильная мышца; 21 — мышца-напрягатель широкой бедренной фасции (спереди от нее — портняжная мышца); 22 — двуглавая мышца бедра; 23 — грудная глубокая мышца; 24 — прямая брюшная мышца; 25 — косая брюшная наружная мышца.
Комплекс основных и дополнительных мышц отвечает за расширение грудной клетки и совершение вдоха. Это грудино-ключично-сосцевидная мышца (m. sternocleidomastoideus), прямая грудная мышца (m. rectus thoracis), большая (m. pectoralis major) и малая (m. pectoralis minor) грудные мышцы и ряд других. Лестничная мышца (m. scalenus) несколькими лентами спускается от мощной шеи к ребрам и при вдохе поворачивает их вперед, эффективно увеличивая объем грудной клетки. Особенно велика по размерам собственно трапециевидная мышца (m. trapezius), которая имеет висцеральное происхождение. Эта мышца разделяется на 5 порций, идущих от головы и позвоночного столба к акромиальному отростку и гребню лопатки; иннервируется она XI-черепным нервом. Помимо дыхательной, указанными мышцами выполняются и другие функции.
Специфична для млекопитающих сухожильно-мускульная стенка — диафрагма (diaphragma), разделяющая грудную и брюшную полости. Она представляет собой систему сухожилий и поперечнополосатых мышц, сходящихся к ее центру со всех сторон от грудины, ребер и поясничных позвонков; через нее проходит пищевод и крупные кровеносные сосуды. Мышцы диафрагмы иннервируются особым диафрагмальным нервом (nervus phreniсus), начинающимся в шейном отделе. Последнее является веским аргументом в пользу того, что грудобрюшная перегородка образовалась из передней части прямой мускулатуры живота и переместились назад по мере разрастания легких. Расслабленная диафрагма куполообразно выпукла. Становясь более плоской при сокращении, она оттесняет органы брюшной полости назад, тем самым растягивая расположенную в грудной клетке плевральную полость, а вместе с ней и всасывающие воздух легкие. Совершается вдох. Возникновение подвижной грудобрюшной преграды позволило предкам млекопитающих резко интенсифицировать вентиляцию легких и повысить общий метаболизм.
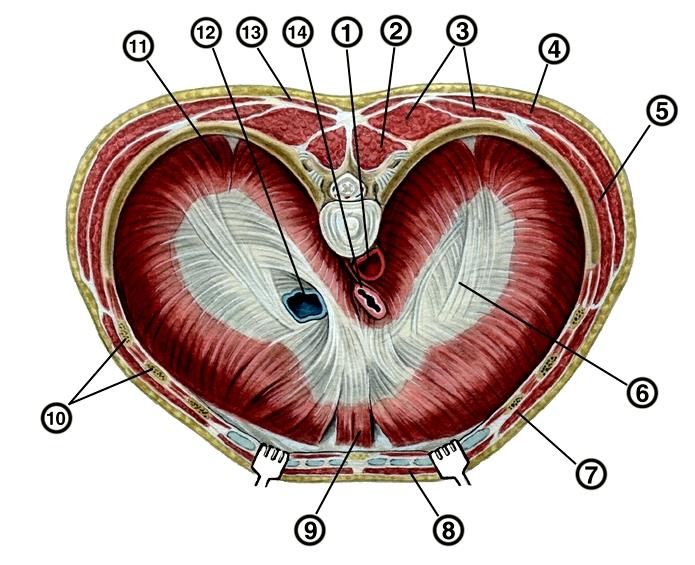
Диафрагма человека (Homo sapiens), вид сверху: 1 — аорта; 2 — поперечная остистая мышца; 3 — мышца, выпрямляющая позвоночник; 4 — широчайшая мышца спины; 5 — передняя зубчатая мышца; 6 — сухожильный центр; 7 — наружная косая мышца живота; 8 — прямая мышца живота; 9 — грудинная часть диафрагмы; 10 — реберная часть диафрагмы; 11 — поясничная часть диафрагмы; 12 — нижняя полая вена; 13 — трапециевидная мышца; 14 — пищевод.
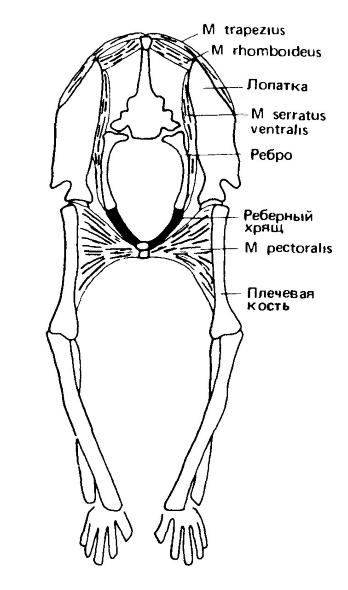
Тазовая конечность поддерживает вес тела за счет прочного соединения таза с крестцом, но у плечевого пояса такого соединения нет. Он лишен прямой связи с черепом и может лишь косвенно соединяться с позвоночником через грудину и ребра. Как и у амфибий, к осевому скелету плечевой пояс подвешен исключительно на мышцах (сильная трапециевидная мышца — одна из них), расположенных в парасагиттальной плоскости. Значение данного явления в системе локомоторных адаптаций вполне понятно. Функционирование конечностей млекопитающих сопряжено с вертикальными движениями и усилиями (взаимодействие с силой тяжести), а также с продольными (разгон и торможение), совместно определяющими сагиттальную плоскость. Использование поперечной оси амфибиями и рептилиями можно рассматривать как наследие рыбообразных предков с их ундуляционным плаванием. В условиях отказа от использования поперечных сил чисто мышечная подвеска пояса передних конечностей оказалась вполне удачной.
Главный элемент мускульной подвески плечевого пояса — сложная зубчатая мышца (m. serratus ventralis), которая широким веером отдельных пучков («зубцов») тянется от редуцированных шейных и от передних грудных ребер к вершинам обеих лопаток. Здесь она крепится очень компактно и допускает свободные повороты лопатки вперед и назад относительно центра зоны крепления, контролируемые другими мышцами. Самый передний пучок зубчатой мышцы называется поднимателем лопатки (m. levator scapulae). Дорсальнее располагается ромбовидная мышца (m. rhomboideus), которая удерживает верхний конец лопасти лопатки, притягивая его к средней линии. Расположенная вентрально грудная мышца (m. pectoralis) удерживает основание лопатки. Таким образом, указанные мышцы удерживают лопатку и конечность в нужном положении, поддерживая целостность всей конструкции. Использование мышц в системе прикрепления плечевого пояса сообщает ей эластичность, которая восполняет отсутствие костной прочности таза. Движения конечностей и туловища здесь взаимно амортизируются, а происходящие при локомоции толчки и сотрясения значительно ослабляются.
Схема, показывающая, как тело млекопитающих поддерживается мышцами передних конечностей.
В связи с особенностями бега млекопитающих, освоивших прыжковые аллюры типа галопа, у многих из них позвоночник в области поясницы приспособлен к вертикальному сгибанию и разгибанию. При прыжках его разгибает расположенный в пояснице мускул распрямитель туловища (m. errector trunci). В нем слиты воедино 3 системы мышечных пучков, которые в грудной области четко разделены. Хвост млекопитающих обычно достаточно тонкий, в связи с чем хвостовая мускулатура уменьшена в объеме. Основное место в ней занимает хвостовая мышца (m. coccygeus). Из вентральных мышц сразу позади тазового пояса у однопроходных развивается сфинктер, замыкающий клоаку; у остальных млекопитающих ему соответствует анальный сфинктер.
Несколько дорсальных мышц грудной конечности отвечают за многие движения плечевой кости относительно плечевого пояса, прикрепляясь около ее головки. На поверхности находятся крупные веерообразные мышцы этого типа: широчайшая спины (m. latissimus dorsi) и дельтовидная (m. deltoideus); первая начинается от поверхности спины и бока, а вторая — от лопатки и ключицы (часто в виде 2 частей). Лопаточная часть дельтовидной мышцы отходит от ости, расположенной по центру лопатки. Один из пучков широчайшей мышцы приобрел связь с лопаткой и отделился в виде большой круглой мышцы (m. teres major). Связанная с широчайшей мышцей у ее окончания глубоко залегающая подлопаточная мышца (m. subscapularis) отходит от внутренней поверхности пояса. Малая круглая мышца (m. teres minor) обосабливается из дельтовидной мышцы и находится у заднего края лопаточной лопасти. Предостная мышца (m. suprаspinаtus) отвечает за разгибание плечевого сустава; для ее размещения на лопатке дополнительно сформировалась передняя костная полка. На лопаточной лопасти также крепится заостная мышца (m. infrаspinаtus). Дорсальная поверхность плечевой кости покрыта крупной трехглавой мышцей плеча (m. triceps brachii), которая начинается от этого элемента, и одной или более головками от прилежащих частей пояса; дистально она прикрепляется к локтевому отростку локтевой кости (это прикрепление фактически объясняет его существование) и служит для разгибания локтя. Дорсальная мускулатура грудной конечности продолжается дистальнее локтя серией мышц-разгибателей.
Большая грудная мышца (m. pectoralis major), возникающая из грудной мышцы, простирается далеко назад поверх грудины и ребер и оканчивается на мощном отростке у проксимального конца плечевой кости. Она с большой силой тянет эту кость вниз и назад, что имеет большое значение при локомоции. От заднего конца коракоида вдоль нижней стороны плечевой кости идет коракоидно-плечевая мышца (m. coracobrachialis). 2 мышцы сгибают конечность в локте: плечевая (m. brachialis), идущая в дистальном направлении от наружного и переднего края плечевой кости, и двуглавая плеча (m. biceps brachii), начинающаяся от коракоида. Результирующее действие рычага стремится поворачивать конечность вниз и вперед или, наоборот, толкать тело вверх и назад при опоре на кисть.
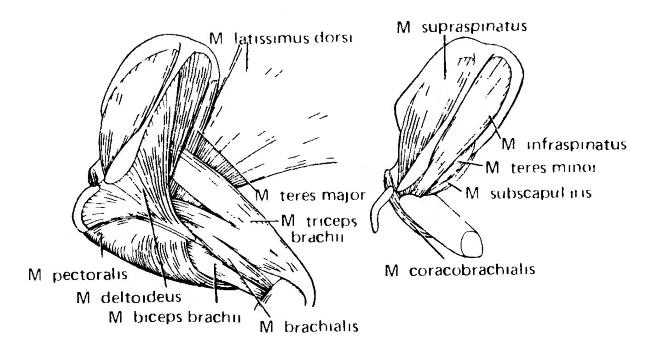
Мышцы левого плечевого пояса и плеча виргинского опоссума (Didelphis virginiana); справа показаны глубокие слои после удаления широчайшей, дельтовидной, грудной, трехглавой, двуглавой и плечевой мышц.
На предплечье располагаются сгибатели и разгибатели кисти, пальцев и пронаторы и супинаторы предплечья. Сухожилия общего разгибателя тянутся в пальцы. Специальные длинные мышцы управляют I-м и V-м пальцами. Однако в дистальной части конечности основное пропульсивное усилие — предплечья и пальцев, выполняется мышцами вентральной, сгибательной поверхности, которые вследствие этого мощнее разгибателей. Длинные сгибатели начинаются от внутреннего надмыщелка плечевой кости и расходятся к нижнему концу предплечья и к кисти. Пальцы сгибает самая глубокая мышца (m. flexor palmaris profundus), идущая от локтя и огибающая с нижней стороны запястье. За запястьем поверхностнее пальцевых сухожилий образуется ее апоневроз, к которому другие длинные сгибатели крепятся проксимально, а некоторые из коротких пальцевых мышц — дистально. Апоневроз расщеплен на 2 налегающих друг на друга сухожильных листка. Длинная ладонная мышца оканчивается поверхностным апоневрозом, который лежит «над» запястьем, а под ним поверхностный сгибатель пальцев имеет (как и лежащий под ним глубокий) широкую систему пальцевых сухожилий. Глубоко в кисти расположены различные короткие сгибатели пальцев.

Мышцы левого предплечья и кисти виргинского опоссума (Didelphis virginiana), несколько схематизированные и упрощенные: А — внешний вид разгибательной поверхности; Б — поверхностные слои сгибательной стороны; В — более глубокие слои сгибательной стороны.
Так как тазовый пояс у млекопитающих срастается с позвоночником, то у них, естественно, отсутствует мускулатура, приводящая в движение тазовые кости, однако, как было показано выше, имеются мышцы туловища, которые прикрепляются к тазу. Дорсальная мускулатура бедра образована поясничной (m. psoas), подвздошной (m. iliacus) и гребешковой (m. pectineus) мышцами, идущими соответственно от поясничного отдела позвоночника, подвздошной и лобковой костей. Группа мышц-разгибателей идет к головке большой берцовой кости, — прямая мышца бедра (m. rectus femoris), начинающаяся от подвздошной кости, и широкие мышцы (mm. vasti), отходящие от бедренной кости; они часто бывают скомпонованы в четырехглавую мышцу бедра (m. quadriceps femoris), оканчиваясь сухожилием, в котором помещается коленная чашечка. Иногда с четырехглавой объединена и портняжная мышца (m. sartorius). Мощные глубокие ягодичные мышцы (m. gluteus medius, m. gluteus minimus) и грушевидная мышца (m. piriformis) идут назад и вниз от подвздошной кости к бедренной и прикрепляются к проксимальному концу кости, за который они посредством рычага поворачивают колено назад, или, наоборот, толкают тело вверх и вперед при опоре на стопу. Кроме того, к малой берцовой кости от подвздошной идет глубокая и часто утрачиваемая тончайшая мышца (m. tenuissimus). Дистальнее колена план строения разгибателей тазовой конечности в целом такой же, что и в грудной конечности.
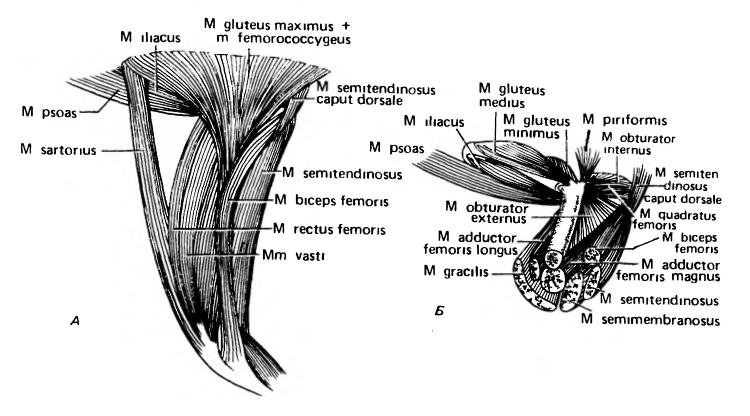
Мышцы таза и бедра виргинского опоссума (Didelphis virginiana): А — вид с поверхности; Б — более глубокие слои мускулатуры. Передний конец обращен влево.
Вентральные мышцы бедренной области главным образом приводят бедренную кость и сгибают колено, т. е. при локомоции поднимают тело от земли и толкают его вперед, в связи с чем они велики и сложны. По расположению выделяются 3 основные их группы.
I. Мышцы, идущие от таза к бедренной кости, по большей части очень глубокие. Они включают 3 основных элемента. а) Крупный мышечный комплекс, мясисто отходящий от значительной части наружной поверхности лобковой и седалищной костей (образование здесь окна связано именно с этим мышечным креплением) и оканчивающийся на нижней стороне бедренной кости около ее головки, тянет бедро вниз. У млекопитающих это главным образом наружная запирательная мышца (m. obturator externus); квадратная мышца бедра (m. quadratus femoris) является обособленной частью этой же мышечной массы. б) Меньший мышечный комплекс, выходящий сзади с внутренней стороны седалищной кости и идущая к головке бедренной кости, представлен внутренней запирательной (m. obturator internus) и меньшими по размеру близнечными мышцами (mm. gemelli). в) Начинающиеся от лобковой и седалищной костей и идущие далеко вниз по вентральной стороне бедренной кости 2 из 2 приводящих мышц бедра (mm. adductor femoris).
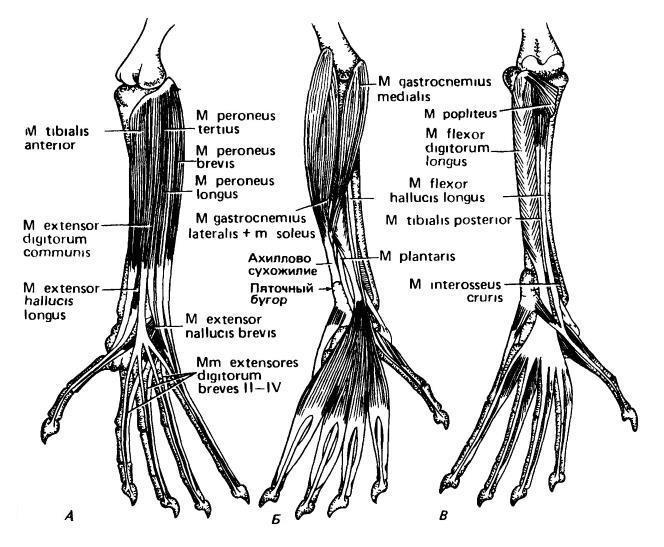
Мышцы левой голени и стопы виргинского опоссума (Didelphis virginiana), несколько схематизированные и упрощенные: А — внешний вид разгибательных поверхностей; Б — поверхностные слои сгибательной стороны; В — более глубокие слои сгибательной стороны.
II. Большая и сложная группа длинных мышц, сгибающих колено, покрывает нижнюю поверхность бедра. Самый поверхностный их пласт — это стройная мышца (m. gracilis). Глубже залегают двуглавая мышца бедра (m. biceps femoris), полусухожильная (m. semitendinosus) и полуперепончатая (m. semimembranosus) мышцы.
III. От нижней стороны хвостовых позвонков идут вперед 2 вентральные хвостовобедренные мышцы — короткая (m. caudofemoralis) и длинная (m. semitendinosus caput dorsale), переходящие в сухожилия, которые оканчиваются посередине длины бедренной кости. У далеких предков млекопитающих они были хорошо развиты; мощно тяня бедро назад, эти мышцы вносили большой вклад в перемещение тела вперед. Однако у млекопитающих эта группа мышц редуцировалась, и окончание короткой мышцы сдвинулось дистальнее.
Длинные вентральные мышцы более дистальных отделов тазовой конечности образуют мощную группу. Большую часть их массы составляет очень важная икроножная мышца (m. gastrocnemius), выпрямляющая стопу. Ее брюшки образуют «икру» ноги, оканчивающуюся ахилловым сухожилием на бугре пяточной кости. Подобно грудной, в тазовой конечности образуются пальцевые сухожилия общего разгибателя и развиваются длинные мышцы, действующие на I-й и V-й пальцы. Длинный сгибатель пальцев переходит в апоневроз, от которого тянутся дистальные сухожилия и мышцы пальцев. На сгибательной стороне имеются глубокие короткие мышцы пальцев.
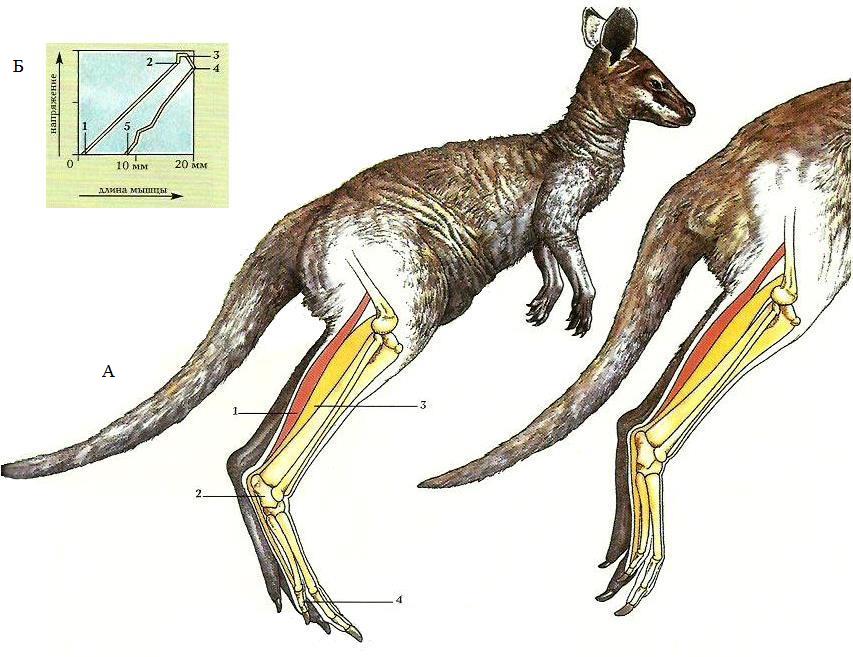
А. У болотного валлаби (Wallabia bicolor) конечности хорошо приспособлены для прыжков. Икроножная мышца (1) оканчивается сухожилием, которое прикрепляется к пятке (2), а подошвенная мышца (3) проходит сверху икроножной и прикрепляется сухожилием к пальцам (4). Напряжение этих мышц приводит к сильному сокращению лодыжки, обеспечивая прыжок.
Б. График показывает изменение длины подошвенной мышцы у прыгающего валлаби по отношению к напряжению мышц. Когда конечности соприкасаются с землей после прыжка (1), мышцы полностью расслаблены. Между (1) и (2) позициями мышцы напрягаются, а конечности сгибаются. Когда конечности разгибаются для нового прыжка, напряжение достигает максимума (3). Мышцы лодыжки сокращаются (4), и животное отталкивается от земли (5).
Адаптивные преимущества трехзвенной Z-образной конструкции парасагиттальных ходильных конечностей связаны с энергетическими проблемами, сопутствующими использованию рычажных систем для перемещения тела животного над грунтом. При этом мышцы конечностей ритмически сокращаются, преодолевая силу тяжести, и неизбежно производят работу, однако в то же время не изменяются ни потенциальная, ни кинетическая энергия животного (высота центра масс над субстратом и скорость движения постоянны). Из этого следует, что производимая мышцами работа поглощается другими мышцами; таким образом, передвижение животного на рычажных конечностях неизбежно сопровождается противоборством мышц и чревато непроизводительными тратами энергии. Между прочим, от подобных проблем полностью свободно колесо: как известно, езда на велосипеде вчетверо экономичнее пешей ходьбы или бега.
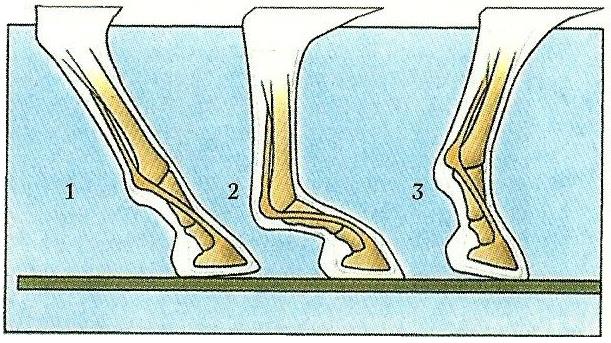
Передвижение лошади (Equus ferus) осуществляется за счет эластичного сухожилия, прикрепленного к копыту. Когда конечность находится на земле, сухожилие напряжено, сохраняя энергию (1). При полном растяжении с максимальной нагрузкой на конечность, сухожилие сохраняет максимальную энергию (2). Когда конечность поднимается, запасенная энергия освобождается, выпрямляя и отталкивая копыто (3).
В цикле движения ноги человека, как показывают расчеты, первая половина фазы опоры, пока нога остается впереди от вертикали тазобедренного сустава, значительно экономичнее второй, когда колено было бы выгоднее обратить назад. В первой половине нагруженной фазы обладатели Z-образной конечности также используют ее как обращенную коленом вперед, а подвижность в пяточном сгибе блокируют при помощи мышц. Но во второй половине они блокируют коленный сустав, используя в качестве действующего сгиба обращенный назад сустав — голеностопный. Трехзвенная конечность, имеющая больше степеней свободы, чем двухзвенная, допускает более пластичный алгоритм движения, позволяя своим обладателям получить некоторую экономию энергии за счет поведенческих реакций.

Подкожные мышцы, обволакивающие тело крота (Talpa europaea). Те, что находятся спереди от передней конечности, являются частью лицевой мускулатуры; позади нее расположена большая подкожная мышца туловища.
Для большинства млекопитающих (высшие приматы составляют исключение) характерно наличие хорошо развитой подкожной мускулатуры. Нередко туловище и шея почти полностью окутаны сплошной оболочкой, которая представляет собой сильно разросшиеся и обособившиеся поверхностные слои грудной мышцы. Этот тонкий пласт получил название подкожной мышцы туловища (m. panniculus carnosus). Волокна других мышц (например, широчайшей мышцы спины) также могут участвовать в формировании этой или иных подкожных мышц, но их роль редко бывает существенна. Зачастую подкожные мышцы приводят в движение те или иные участки кожи. Особенно подвижна кожа у копытных; это можно наблюдать на примере подергивания кожи лошади в том месте, где уселась муха. Подкожная мускулатура участвует и в терморегуляции, отвечая за изменение положения волосяного покрова и свертывание тела в клубок. Поднятие игл (у ехидн, ежей, дикобразов) происходит вследствие сокращения поперечнополосатых мышц, которыми снабжены основные части самих игл. Такие же поперечнополосатые мышцы подходят к вибриссам. Очень сильна подкожная мускулатура у ехидн, броненосцев, панголинов и ежей; у них она имеет кольцевую форму, что обусловливает способность этих животных свертываться в защитный шар.

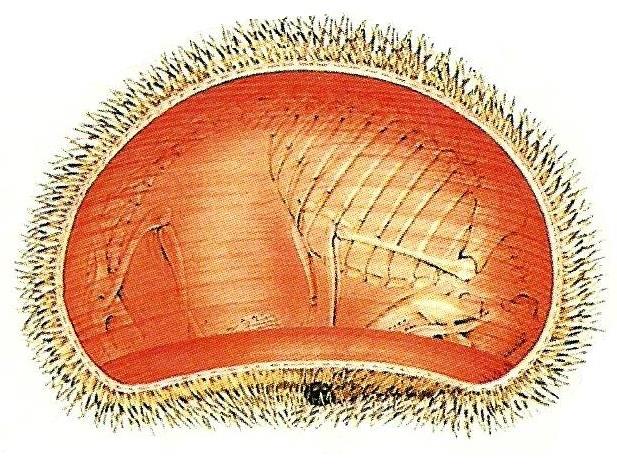
Подкожная мускулатура свернувшегося ежа (Erinaceus europaeus).
Отдельно следует упомянуть мускулатуру желудка и кишечника, образованную гладкими мышечными волокнами. Обычно имеется слой продольных и слой кольцевых волокон, идущих вдоль тракта. Они иннервируются нервами вегетативной (непроизвольной) нервной системы, которые могут или стимулировать, или подавлять их активность; на них также могут воздействовать гормоны. Однако в большинстве случаев волокна работают вполне независимо от какого бы то ни было центрального управления, стимулируемые сокращением соседних волокон или местной нервной сетью. Следующие друг за другом волны мышечного сокращения вызывают сужения желудочно-кишечного тракта, которые перемещаются назад и толкают пищу перед собой. Данный процесс — перистальтика — служит основным средством транспортировки пищи.
вперед
в оглавление