в оглавление
назад
Нервная система
Нервная система млекопитающих высоко интегрирована и упорядочена, благодаря чему достигается совершенство взаимосвязей составных частей организма и его отношений с внешней средой.
По морфологическим признакам нервную систему подразделяют на центральную и периферическую. Центральная часть представлена головным и спинным мозгом. Сложно организованный головной мозг находится в мозговой полости черепа, поблизости сосредоточены основные органы чувств. Сзади он переходит в дорсально расположенный спинной мозг. Периферическую нервную систему образуют парные нервы, отходящие от головного и спинного мозга, а также их ответвления (всего их насчитывают свыше 200). По ходу нервов могут располагаться скопления нервных клеток — нервные узлы, или ганглии — также относящиеся к периферической нервной системе. Подобно другим позвоночным, нервная система млекопитающих происходит из внешнего зародышевого листа — эктодермы. Сначала образуются нервная трубка и примыкающие к ней нервные гребни, затем из них формируются мозг и проникающие в разные части тела периферические компоненты.
Нервные клетки и волокна
Основным структурным и функциональным элементом нервной системы является нервная клетка — нейрон, осуществляющий генерирование и распространение нервных импульсов. Во взрослом организме не наблюдается признаков митоза нейронов, что указывает на приобретение полного, или, по крайней мере, почти полного набора нервных клеток уже ко времени появления на свет (примечательная особенность нервной системы). Вследствие этого разрушение нервных клеток в результате ранения или болезни представляет собой невосполнимую утрату — отростки клеток способны к регенерации, если сохранно тело клетки, но коль скоро утрачен нейрон, возмещение его невозможно.
У нейрона имеется тело и отходящие от него отростки. Ткань головного и спинного мозга, а также ганглиев содержит многочисленные тела нейронов. Главная их функция — питание и энергетическое обеспечение отростков. Тела разнообразны по своей форме, однако чаще всего она бывает звездчатой благодаря многочисленным отросткам. В их протоплазме имеются различные характерные структуры, не встречающиеся в клетках других тканей. В первую очередь это тельца Ниссля, которые содержат большое количество РНК и многочисленные рибосомы. Все процессы химического синтеза протекают в теле нейрона, откуда продукты поступают в различные отростки.
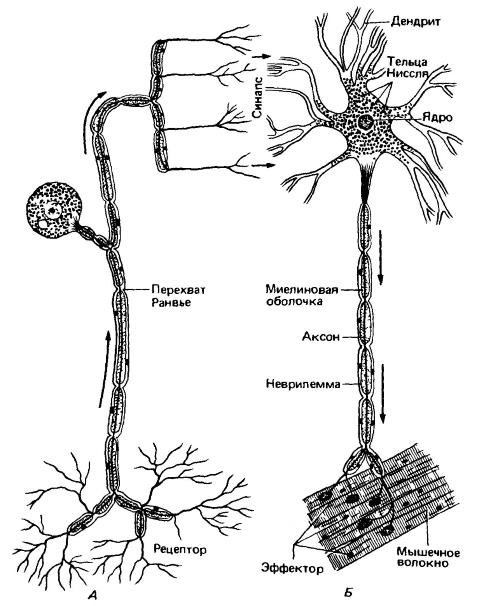
Два типа нейронов. А — чувствующий (афферентный) нейрон спинномозговых и черепных нервов. От рецептора к телу клетки, помещающемуся в спинномозговом ганглии, тянется длинный аксоноподобный отросток. От тела клетки отросток направляется в спинной мозг и там ветвится. Б — двигательный (эфферентный) нейрон. Тело клетки расположено в спинном мозге. Длинный аксон соединяется с эффектором (мышечным волокном). На обоих рисунках периферический отросток (аксон двигательной клетки) изображен сильно укороченным. При соблюдении данного масштаба они должны были достигать нескольких метров в длину.
Отростки нейронов представляют собой тончайшие нервные волокна, собственно и служащие для передачи нервных импульсов. Центростремительные, или афферентные, волокна проводят импульсы от периферии к телу нейрона, а центробежные, или эфферентные, — от тела нейрона к иннервируемому им органу или к другим нейронам. Пучки нервных волокон образуют нервы, окруженные соединительнотканными оболочками и пронизывающие почти все части тела. Кроме того, эти волокна составляют основную массу ткани во всех отделах центральной и периферической нервной системы. Большая часть нервных волокон имеет настолько большую длину, что бывает трудно установить, где лежат тела клеток, от которых они отходят. Тем не менее, данные волокна всегда представляют собой отростки нейронов, а не самостоятельные образования. В нервной ткани хорошо развито капиллярное кровообращение.
Длина и пространственная организация отростков достаточно изменчива. Типичными обычно считаются двигательные (эфферентные) нейроны, иннервирующие поперечнополосатые мышцы. Короткие тонкие ветвистые отростки этих нейронов, проводящие импульсы к телу клетки, в соответствии с их древовидной формой именуются дендритами. Единственный практически неразветвленный относительно толстый и длинный отросток называется аксоном; у крупных животных он нередко достигает нескольких метров в длину. По аксону импульсы идут от тела клетки. Второй распространенный тип — чувствующие (афферентные) нейроны, проводящие импульсы по направлению к центральной нервной системе. Здесь оба отростка — длинный отросток, идущий от точки восприятия к телу клетки, расположенному неподалеку от спинного мозга, и второй длинный отросток, входящий в спинной мозг, — сходны по своему строению с аксоном двигательного нейрона. Поскольку у двигательных нейронов помимо аксона имеется по нескольку дендритов, эти нейроны называют мультиполярными (подобные нейроны встречаются также и внутри центральной нервной системы). Типичные чувствующие нейроны биполярны, так как обладают двумя длинными отростками.
С функциональной точки зрения наиболее важной частью крупного нервного волокна является аксоплазма — тонкий тяж протоплазмы, продолжающий протоплазму тела клетки. В аксоплазме имеются многочисленные нитевидные продольные нейрофибриллы, которые являются продолжением фибрилл, расположенных в теле нейрона. Аксоплазма окружена очень тонкой плазматической мембраной и поверх нее почти всегда бывает одета дополнительными оболочками. Над плазматической мембраной располагается прочная, неэластичная многоклеточная оболочка — неврилемма. В центральной нервной системе клетки этой оболочки принадлежат к нейроглии. Вне мозга эта оболочка образована особыми шванновскими клетками, происходящими из нервного гребня. Шванновские клетки окружают волокно миелиновой оболочкой из жирового вещества.
Полностью сформированная миелиновая оболочка придает нервным волокнам ярко-белый цвет. У большинства периферических аксонов миелиновая оболочка имеется, но иногда встречаются немиелинизированные волокна, имеющие серый цвет. Обычно шванновская клетка наматывается на участок отростка нейрона, образуя подобие рулета, но с тем отличием, что слоев очень много и они очень тонкие. В промежутке между двумя соседними шванновскими клетками миелиновая оболочка периферического волокна прерывается. Такая область именуется перехватом Ранвье. Если перерезать нервное волокно, то его часть, лежащая дистальнее разреза, дегенерирует, а проксимальная часть и тело клетки могут повреждаться в различной степени. Однако часто периферическое волокно регенерирует, вновь вырастая из «обрезка», связанного с телом клетки. По-видимому, восстановлению былого пути прохождения волокна способствует сохранение обкладочных клеток, окружавших его разрушившийся участок.

Строение нервных волокон на светооптическом (А, Б) и ультрамикроскопическом (а, б) уровнях: А, а — миелиновое волокно; Б, б — безмиелиновое волокно; 1 — осевые цилиндры; 2 — миелиновый слой; 3 — соединительная ткань; 4 — насечка миелина; 5 — ядро нейролеммоцита; 6 — узловой перехват; 7 — микротрубочки; 8 — нейрофиламенты; 9 — митохондрии; 10 — мезаксон; 11 — базальная мембрана.
Нервный импульс
Нейроны могут быть стимулированы другими нейронами, повреждением или сенсорными рецепторами. Во всех случаях принцип один и тот же. Мембрана нейрона обычно поляризована. Иными словами, между ее наружной и внутренней поверхностью имеется электрический потенциал, который в неактивном нейроне называется потенциалом покоя и создает устойчивое состояние готовности, сходное с состоянием электрической батареи, накопившей энергию, которая высвобождается в случае надобности. Потенциал покоя обусловлен неодинаковыми концентрациями катионов калия внутри и снаружи клетки. Когда клетка находится в состоянии покоя, ее внутренний заряд отрицателен по отношению к наружному. При деполяризации клетки ее мембранный потенциал снижается по направлению к нулю. Когда же мембранный потенциал становится еще более отрицательным, говорят, что клетка «гиперполяризована».
Если потенциал покоя падает ниже определенного порогового значения, вдоль мембраны распространяется потенциал действия. Он длится недолго и вызывается закономерными изменениями относительных концентраций катионов натрия и калия по обе стороны мембраны, проникающие сквозь нее через специальные каналы. Для нейронов млекопитающих известно больше 12 типов потенциалозависимых ионных каналов. Необходимая энергия (как и в случае сокращения мышечного волокна) освобождается в результате расщепления аденозинтрифосфата в аксоплазме. Потенциал действия проходит к концу аксона в виде электрической волны. Он всегда имеет одну и ту же амплитуду (высоту), обычно зависящую от диаметра аксона. По более крупным аксонам распространяются более высокие потенциалы действия, чем по более тонким; диаметр же аксонов варьирует от 1 до 20 мкм.
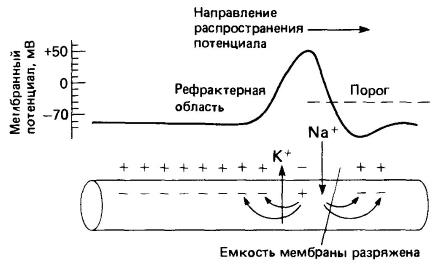
Распространение потенциала действия благодаря прохождению ионов калия и натрия через мембрану аксона.
Скорость распространения нервного импульса вдоль волокна все же существенно меньше скорости распространения электрического импульса. В среднем аксоны млекопитающих обеспечивают более быстрое проведение, чем аксоны низших позвоночных — например, вдвое быстрое, чем у лягушки. Однако даже для млекопитающих наибольшая зарегистрированная скорость распространения нервного импульса составляет лишь 130 м/с, а для некоторых аксонов не превышает 0,5 м/с. Большие скорости характерны для аксонов большего диаметра. Несмотря на то, что скорость проведения в некоторых аксонах и достигает значительных величин, все же у таких крупных животных, как, например, слон, наблюдается очевидный разрыв во времени между восприятием стимула и ответом. Объясняется это тем, что даже в случае простейшего рефлекса сигнал должен пройти вдоль нервных волокон многие метры.
После прохождения каждого потенциала возникает рефрактерный период, в течение которого мембрана восстанавливает свое нормальное ионное равновесие и нормальный потенциал покоя. Поскольку во время рефрактерного периода новый потенциал действия возникнуть не может, рефрактерные свойства аксона определяют максимальную частоту потенциалов действия.
Когда возникает потенциал действия, говорят, что нейрон «разряжается». Этот потенциал часто проявляется в виде пика на экране осциллографа, приспособленного для измерения мембранных потенциалов с помощью электродов, введенных в нервную ткань. Нейрон разряжается по принципу «все или ничего». Не существует сильных или слабых импульсов: нейрон или включается полностью, или не включается вовсе; соответственно, на осциллографе либо наблюдается полный пик, либо его совсем нет (в высших мозговых центрах специальные типы нейронов, в виде исключения, могут действовать по иному принципу). Конечно, импульсы, передающиеся по нервам, различаются по силе, но эти различия обусловлены иными причинами. В разное время в нерве может включаться различное число аксонов. Чрезвычайно важным является то обстоятельство, что импульсы не бывают единичными, а следуют один за другим сериями с различной частотой и могут вызывать кумулятивный эффект. Частота импульсов ограничена рефрактерными свойствами нейрона и зависит от силы его стимуляции.
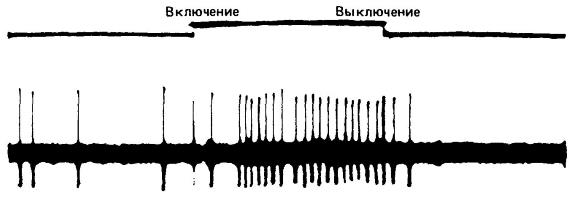
Внеклеточное отведение нервных импульсов от нейрона в зрительной системе кошки (Felis silvestris). При включении стимула частота импульсации повышается.
Нервный импульс неспецифичен и не имеет обратного адреса. Природа «ощущений», возникающих в мозге, зависит от того, какой центр принял сигнал, а не от каких-либо особенностей полученных импульсов. В норме импульсы передаются вдоль волокон только в одном направлении, но экспериментально можно выявить способность к проведению импульсов в обоих направлениях. Односторонняя передача, наблюдаемая в работающей нервной системе, обеспечивается особой схемой соединения волокон: нейроны «поляризованы» морфологически.
У позвоночных никогда не бывает так, чтобы связь между чувствительным органом (рецептором) и реагирующей двигательной или железистой структурой (эффектором) осуществлялась одним-единственным нейроном. Она осуществляется посредством цепи нейронов, состоящей по крайней мере из двух, а обычно и из большего числа клеток. Место контакта двух смежных нейронов именуется синапсом. Кончик аксона первого из них (пресинаптического нейрона) разделяется на тонкие, как правило, оканчивающиеся синаптическими бляшками, или «пуговками», веточки, которые переплетаются с дендритами второго (постсинаптического) нейрона или обвивают его тело. Несмотря на то, что отростки таких двух клеток могут располагаться очень близко друг от друга (на расстоянии всего лишь 20 нм), их протоплазма никогда не объединяется.
Передача импульса через синапс осуществляется с помощью электрических сигналов или, в большинстве случаев, с помощью особых химических веществ-посредников — нейромедиаторов. Нейромедиатор секретируется телом нейрона и в микроскопическом количестве выделяется его пресинаптическими окончаниями. Диффундирует через синаптическую щель, он влияет на потенциал покоя воспринимающей мембраны следующего нейрона и, следовательно, на готовность этого нейрона генерировать потенциалы действия. В роли основного нейромедиатора выступает ацетилхолин. Иногда отдельно рассматривают нейромодуляторы, отличающиеся от нейромедиаторов тем, что выбрасываются в синаптическую щель в меньших количествах и действуют медленнее.
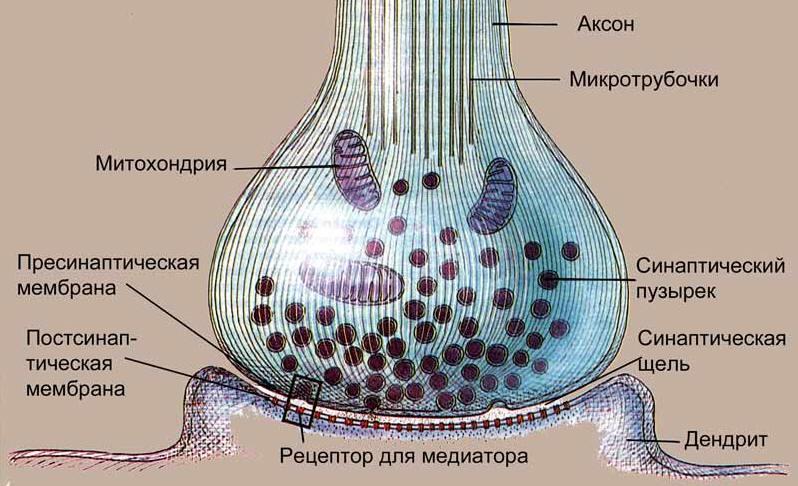
Схема строения синапса.
Простейшей функцией нервной системы является рефлекс. К рефлекторным реакциям принадлежат, например, «автоматическое» отдергивание пальца, прикоснувшегося к раскаленной печке, или быстрый и резкий выдох (кашель) в ответ на раздражение слизистой оболочки дыхательных путей. Как правило, сигнал поступает от рецепторных клеток и по длинному афферентному нервному волокну проводится к центральной нервной системе. Тело чувствующего нейрона, которому принадлежит волокно, расположено наряду с многими телами других нейронов в ганглии неподалеку от спинного или головного мозга. Волокно продолжается далее прямо в центральную нервную систему. Здесь оно обычно соединяется не с одним лишь нейроном, но, разветвившись, передает стимул через синапсы целой серии нейронов. Вместе с тем каждый из таких нейронов может получать импульсы от многих афферентных волокон, и таким образом, могут осуществляться различные сочетания связей между рецепторами и эффекторами. По-видимому, единичный импульс редко способен «включить» постсинаптический нейрон: возбуждение наблюдается только в том случае, если импульсы поступают из нескольких источников.
В случае простейшего рефлекса афферентные нейроны могут передавать центростремительные импульсы прямо эфферентным, или двигательным, нейронам. Тела последних лежат в центральной нервной системе. От них отходят длинные тонкие аксоны к эффекторам («концевым органам»); обычно ими бывают мышечные волокна. Однако почти все рефлекторные дуги, за исключением тех, которые связаны с поддержанием позы, на одну ступень сложнее, т. е. состоят не из двух, а из трех последовательных нейронов. В этом случае идущие к центральной нервной системе волокна не имеют синаптических контактов непосредственно с двигательными нейронами, а соединяются с нейронами, целиком лежащими в пределах центральной нервной системы. Эти последние, называемые ассоциативными (или вставочными) нейронами, подобно афферентным дают разветвленные отростки, вступающие в синаптические контакты со многими двигательными нейронами. Таким образом, число потенциальных ответов на определенный импульс от рецептора и, кроме того, число импульсов, от рецепторов, вызывающих один и тот же ответ, еще сильнее увеличивается. Стимулы могут суммироваться, и, образно говоря, появляется возможность выбора одного из вариантов ответа. До сих пор говорилось, что через синаптические контакты осуществляется возбуждение клеток. Однако нередко на синаптическом уровне происходит торможение, блокирующее стимулы, которые клетка может получить от других источников. Более того, уже на таком относительно простом уровне нервной деятельности существуют механизмы обратной связи, которые еще больше усложняют картину. Увеличение числа ассоциативных нейронов и более сложных обратных связей — модуляторов, резонансных блоков — привело к возникновению высших ассоциативных центров, получающих большое количество разнообразных сигналов и после их сортировки, сравнения и интеграции генерирующих большое количество разнообразных ответов.
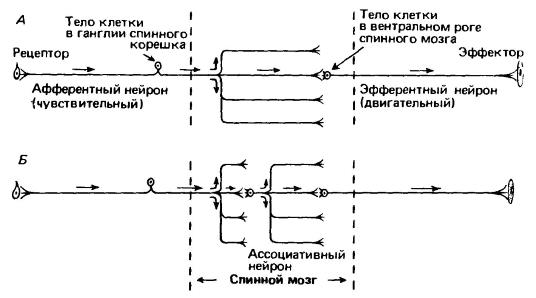
Схемы простых рефлексов (область между штриховыми линиями отграничивает ту часть рефлекторной дуги, которая располагается в спинном мозге). А — двухнейронный рефлекс. Б — введение ассоциативного нейрона увеличивает число возможных путей распространения импульса. Представленная схема сильно упрощена. Большая часть эфферентных (и ассоциативных) нейронов в действительности получает сигналы от большого числа разнообразных нейронов, обеспечивая оценку суммарного импульса.