в оглавление
назад
Периферическая нервная система
Спинномозговые нервы
Спинномозговые, или спинальные, нервы расположены симметрично слева и справа в каждом сегменте тела. Количество их велико — у человека, к примеру, насчитывается 31 пара спинномозговых нервов. Каждый нерв связан со спинным мозгом двумя корешками — спинным и брюшным (задним и передним по терминологии, принятой в анатомии человека). Брюшной корешок (radix ventralis) выходит наружу из вентролатеральной части стенки спинномозговой трубки. Спинной корешок (radix dorsalis), несущий крупный ганглий (ganglion spinale), входит в боковую стенку трубки. Брюшной и спинной корешки объединяются, образуя главный ствол нерва, в канале позвоночного столба. За пределами позвоночника он делится на разнообразные ветви (rami). Крупная спинная ветвь (ramus dorsalis) посылает волокна к дорсальной части осевой мускулатуры и к коже спины. Брюшная ветвь (ramus ventralis) снабжает более латерально и вентрально расположенные участки кожи и гипаксиальную мускулатуру стенки тела. Также имеется ветвь, следующая к внутренностям.
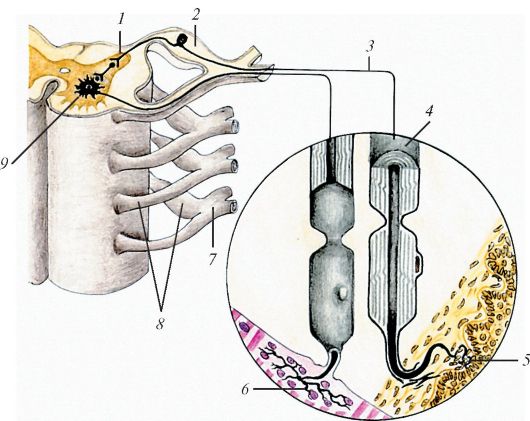
Схема строения периферического нерва: 1 — задний рог спинного мозга; 2 — спинномозговой узел; 3 — отросток (дендрит) чувствительной нервной клетки; 4 — миелиновое нервное волокно; 5 — нервно-чувствительное окончание; 6 — нервно-мышечное окончание; 7 — спинномозговой нерв; 8 — корешки спинномозговых нервов; 9 — передний рог спинного мозга.
Главный ствол спинномозгового нерва и его основные ветви включают в свой состав как афферентные, так и эфферентные волокна. Однако корешки имеют четкое разграничение функций. Брюшные корешки проводят импульсы центробежно, по эфферентным волокнам нейронов, клеточные тела которых лежат в спинном мозге. Спинные корешки, напротив, включают преимущественно афферентные волокна, проводящие центростремительные импульсы. Тела соответствующих нейронов лежат в ганглии спинного корешка. Эти клетки имеют ножку, которая почти сразу Т-образно разветвляется: один отросток доставляет импульсы от сенсорных структур, другой передает их центростремительно к центральной нервной системе.
В большинстве сегментов туловища спинномозговой нерв представляет собой дискретную структуру, не имея соединений ни с задними, ни с передними такими же нервами. Он иннервирует туловищные мышцы определенного сегмента, а афферентными компонентами обеспечивает соответствующую полоску кожи с некоторым перекрыванием по отношению к нервам соседних сегментов. Однако в конечностях имеет место соединение ветвей соседних спинномозговых нервов с образованием сплетений — плечевого (plexus brachialis) и пояснично-крестцового (plexus lumbosacralis). В результате мышцы каждой конечности обслуживаются волокнами нескольких спинномозговых нервов.
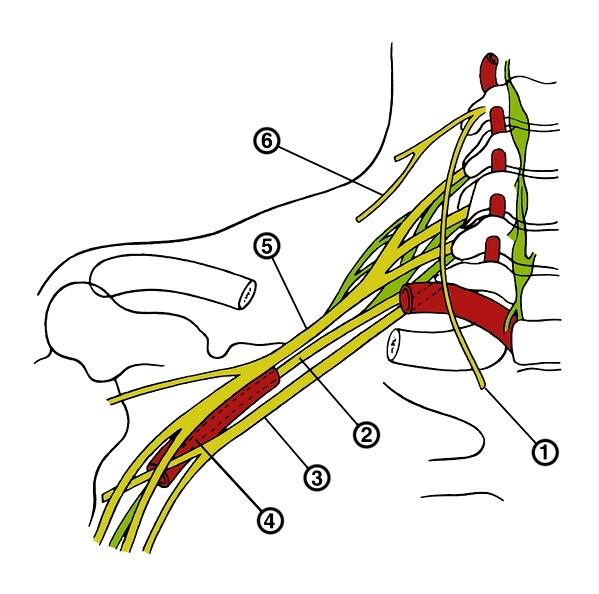
Схема строения плечевого сплетения человека: 1 — диафрагмальный нерв; 2 — задний ствол; 3 — медиальный ствол; 4 — подмышечная артерия; 5 — латеральный ствол; 6 — надключичный нерв.
Черепные нервы
У млекопитающих, как и у других амниот, имеется 12 парных черепных нервов (nervi craniales), получивших свои номера и названия в соответствии с их расположением и выполняемыми функциями.
I) Обонятельный нерв (nervus olfactorius) — чувствующий; иннервирует обонятельные рецепторы. Имеет своеобразную структуру. Обычно тела нейронов чувствующего нерва находятся в ганглии, расположенном вблизи спинного и головного мозга. В данном же случае нервные волокна принадлежат не нормальным нейронам, а клеткам обонятельного эпителия, от которых их тонкие пучки тянутся внутрь, проходят сквозь продырявленную пластинку (lamina cribrosa) решетчатой кости и входят в обонятельную луковицу, лежащую на переднем конце головного мозга. У некоторых форм носовая полость находится на значительном расстоянии от обонятельной луковицы, в связи с чем оформлен единый (парный) обонятельный нерв; обычно, однако, головной мозг вытянут вперед и нерв состоит из множества коротких пучков волокон. У млекопитающих с хорошо развитым вомероназальным органом для его обслуживания обонятельный нерв формирует отдельную ветвь.
II) Зрительный нерв (nervus opticus) — чувствующий; иннервирует сетчатку. Собственно говоря, эта структура представляют собой не нерв, а внутримозговой тракт, поскольку сетчатка формируется в качестве мозгового выроста. Тела чувствительных клеток расположены периферически — в ганглионарном слое сетчатки. Их волокна собираются в нервный тяж, который попадает в полость черепа через зрительное отверстие (foramen opticum) глазоклиновидной кости. Далее левый и правый нервные тяжи пересекаются в зрительной хиазме и входят в промежуточный мозг.
III) Глазодвигательный нерв (nervus oculomotorius) — двигательный; иннервирует нижнюю косую, верхнюю, нижнюю и внутреннюю прямую мышцу глаза, а также ресничную мышцу и мышцу радужины. Отходит от среднего мозга и покидает череп через переднее рваное отверстие (foramen lacerum anterius) между основной крыловидной и глазоклиновидной костями внутри глазничной полости.
IV) Блоковый нерв (nervus trochlearis) — двигательный; иннервирует верхнюю косую мышцу глаза, называемую также блоковой мышцей. Этот небольшой нерв проходит весьма необычно. С каждой стороны он берет начало вентрально внутри среднего мозга, но вместо того, чтобы сразу выйти наружу, он поворачивает в толще мозга вверх и пере-ходит на противоположную сторону, где появляется на поверхности дорсально на границе между средним мозгом и мозжечком. Как и предыдущий, описываемый нерв покидает череп через переднее рваное отверстие.
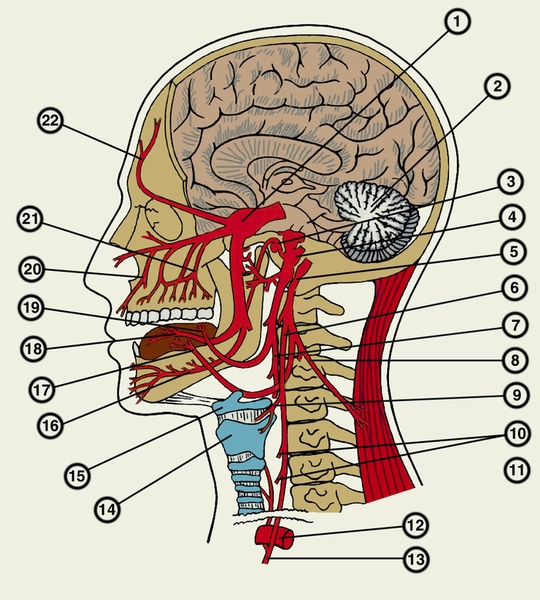
Схематическое изображение стволов и основных ветвей черепных нервов на примере человека: 1 — ганглий тройничного нерва; 2 — мозжечок; 3 — барабанная струна; 4 — лицевой нерв; 5 — нижний ганглий блуждающего нерва; 6 — глоточная ветвь блуждающего нерва; 7 — глоточная ветвь языкоглоточного нерва; 8 — добавочный нерв; 9 — верхний гортанный нерв; 10 — сердечные ветви блуждающего нерва; 11 — возвратный гортанный нерв; 12 — дуга аорты; 13 — ствол блуждающего нерва; 14 — щитовидный хрящ гортани; 15 — подъязычная кость; 16 — нижний альвеолярный нерв; 17 — подъязычный нерв; 18 — язычный нерв; 19 — язычные ветви; 20 — ветвь подглазничного нерва; 21 — верхние альвеолярные нервы; 22 — лобный нерв.
V) Тройничный нерв (nervus trigeminus) — смешанный; чувствующий для кожи передней части головы и слизистых оболочек носовой и ротовой полостей, околоносовых пазух и зубов; двигательный для жевательных мышц; здесь же проходят часть своего пути секреторные волокна в слезную и слюнные железы. Разделяется на три основные ветви: глазничную (ramus ophthalmicus), верхнечелюстную (ramus maxillaris) и нижнечелюстную (ramus mandibularis). Глазничная ветвь проходит вперед в глубине глазницы и иннервирует кожу и слизистые. Две другие ветви иннервируют мускулатуру верхней и нижней челюстей. Общий ствол нерва отходит от продолговатого мозга, мозговую полость ветви покидают соответственно через переднее рваное (foramen lacerum anterius), круглое (foramen rotundum) и овальное (foramen ovale) отверстия внутри глазничной полости. Ветвь, отвечающая за иннервацию верхнечелюстной мускулатуры, затем выходит за пределы черепа сквозь подглазничное отверстие (foramen infraorbitale).
VI) Отводящий нерв (nervus abducens) — двигательный; иннервирует наружную прямую мышцу глаза, которая отводит глаз назад или вбок. Этот небольшой нерв отходит вентрально от переднего конца продолговатого мозга и выходит из черепа через переднее рваное отверстие.
VII) Лицевой нерв (nervus facialis) — смешанный; двигательный для всей мимической мускулатуры, т. е. для губных, щечных, носовых, ушных и мышц век; в нем же проходит часть вкусовых волокон с языка и парасимпатические секреторные волокна для слезной железы, а также нижнечелюстной и подъязычной слюнных желез. Нерв выходит из мозга в области трапециевидного тела, проходит через каменистую часть височной кости и покидает черепную полость через наружное отверстие лицевого канала (foramen stylomastoideum) позади слухового барабана. Делится на большое количество ветвей.
VIII) Слуховой нерв (nervus acuslicus) — чувствующий; иннервирует равновесно-слуховой сенсорный орган. Представляет собой обособившуюся часть лицевого нерва. В отличие от обонятельного и зрительного нервов, нервные волокна здесь принадлежат настоящим ганглионарным клеткам, тела которых располагаются периферически, в слуховой капсуле, поблизости от рецепторов. У млекопитающих имеются две главные ветви слухового нерва, каждая с собственным ганглием: преддверный нерв (nervus vestibularis), обслуживающий переднюю часть системы каналов и мешочков, и улитковый нерв (nervus cochlearis), который помимо задней части органов равновесия обслуживает слуховой орган улитки. С поверхности мозговой коробки слуховой нерв не виден, потому что его волокна идут прямо из слуховой капсулы через внутренний слуховой проход (meatus acusticus internus) в черепную полость, где сверху входят в продолговатый мозг.
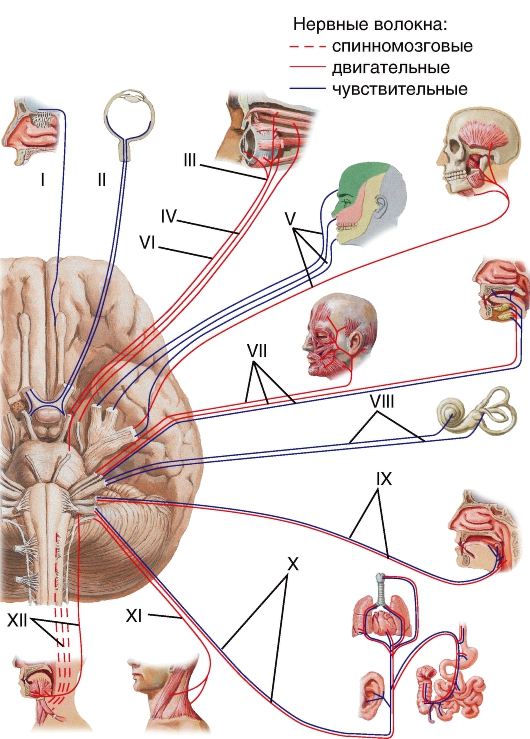
Схема функциональности черепных нервов на примере человека (по Сапину и Биличу, 2007): I — обонятельный; II — зрительный; III — глазодвигательный; IV — блоковый; V — тройничный; VI — отводящий; VII — лицевой; VIII — слуховой; IX — языкоглоточный; X — блуждающий; XI — добавочный; XII — подъязычный.
IX) Языкоглоточный нерв (nervus glossopharyngeus) — смешанный; содержит чувствующие волокна со слизистой оболочки корня языка, миндалины, глотки и среднего уха, вкусовые волокна с корня языка и двигательные волокна для поперечнополосатой мускулатуры глотки; здесь же проходят часть своего пути парасимпатические секреторные волокна для околоушной слюнной железы. Этот небольшой нерв отходит от продолговатого мозга, полость черепа покидает через заднее рваное отверстие (foramen lacerum posterius).
X) Блуждающий нерв (nervus vagus) — смешанный; парасимпатические волокна иннервирует сердце, пищеварительные и дыхательные органы, также имеется небольшое количество чувствующих волокон от кожи шеи и наружного уха. Отходит от продолговатого мозга, из черепа выходит через заднее рваное отверстие. В туловище блуждающий нерв проникает в качестве внутренностной ветви (ramus splanchnicus). Это самый крупный черепной нерв, обслуживающий наиболее обширную область.
XI) Добавочный нерв (nervus аccessorius) — двигательный; иннервирует трапециевидную, плечеголовную и грудинночелюстную мышцы. Является частью блуждающего нерва, обособившейся в связи с мощным развитием у млекопитающих трапециевидной мышцы. Отходит от спинного мозга в области шеи, но выходит через заднее рваное отверстие.
XII) Подъязычный нерв (nervus hypoglossus) — двигательный; иннервирует мускулатуру языка и подъязычной кости. Берет начало от задней части продолговатого мозга. Обычно проходит через отдельное, иногда множественное, подъязычное отверстие (foramen hypoglossi) в дне затылка.
Как правило, нервы очень хрупки и растяжение любого из них приводит к травме. Даже активно двигающиеся животные просто имеют волокна достаточной длины; если она не используются полностью, то нервы «провисают». В отличие от сказанного, челюстные нервы полосатиковых китов (Balaenopteridae) весьма эластичны. Сами по себе их волокна не растягиваются, но они намотаны на центральную часть и покрыты белком эластином. При усилии поверхностный белок растягивается, а нервы разматываются с сердцевины. Как известно, процесс питания полосатиков требует фильтрации огромных объемов океанской воды, насыщенной планктоном. Для подобных действий им необходимо значительно растягивать ротовую полость. Можно предположить, что эластичные нервы позволили этим животным управлять процессом заглатывания, увеличить объем фильтруемой воды, а в итоге дали возможность достичь гигантских размеров.
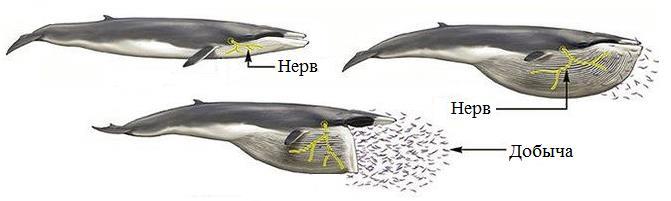
Схема растяжения челюстного нерва (обозначен желтым) полосатика в процессе питания.
Соматический и вегетативный отделы нервной системы
Нервные волокна — и афферентные, и эфферентные — функционально могут быть разделены на соматические и вегетативные, входящие в состав одноименных отделов нервной системы. В спинномозговых нервах присутствуют четыре типа волокон: два афферентных в спинном корешке и два эфферентных в брюшном. В стволе спинномозгового нерва на выходе за пределы позвоночного столба также имеется все четыре указанных типа. Однако далее наиболее характерные крупные стволы включают, как правило, только два типа соматических волокон, а большая часть вегетативных волокон в составе вегетативной ветви направляется в вентральном направлении. В головной области наряду с указанными четырьмя типами можно выделить еще три. Вдобавок к обычной гладкой висцеральной мускулатуре, которая есть и в туловище, в голове и области глотки имеется особая поперечнополосатая висцеральная мускулатура; иннервирующие ее нервные компоненты объединяют под названием особых вегетативно-двигательных. Волокна от органов вкуса выделяют в качестве особых вегетативно-чувствующих. Далее, в голове присутствуют особые соматические сенсорные структуры, отвечающие за обоняние, зрение и слух; идущие от них нервы называют особыми соматически-чувствующими.
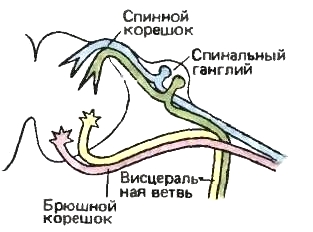
Схема распределения различных типов волокон в спинных и брюшных корешках спинномозговых нервов млекопитающих. Соматически-чувствующие волокна окрашены голубым; вегетативно-чувствующие — зеленым; вегетативно-двигательные — желтым; соматически-двигательные — красным. Спинной корешок почти исключительно чувствующий, и почти все двигательные волокна сосредоточены в брюшном корешке.
В целом соматическая система у млекопитающих очень хорошо развита. Относящиеся к ней афферентные волокна проводят импульсы от чувствительных окончаний кожи, скелетных мышц, сухожилий и сенсорных органов (экстероцептивная и проприоцептивная группы по физиологической терминологии). В двигательном компоненте эфферентные волокна идут к соматической мускулатуре — поперечнополосатым произвольным мышцам тела и конечностей. Данной системе принадлежит большинство (11 пар из 12) черепных и часть спинномозговых нервов. Большую часть соматической активности контролирует головной мозг. Соматическая система отвечает за восприятие внешних стимулов и положения частей тела в пространстве. В результате ее работы достигается тонкая координация движений, которая во многом подлежит контролю.
Вегетативная система именуется также автономной, или висцеральной (т. е. внутренностной). Ее афферентные волокна передают информацию от интероцептивных чувствительных окончаний, которые находятся в сосудах, пищеварительном тракте и других внутренних органах. Эфферентные волокна, в свою очередь, проводят сигналы к гладкой мускулатуре пищеварительного тракта и сосудов, сердечной мышце и т. п., а также к различным железам. В состав вегетативной системы входит черепной блуждающий нерв, простирающийся вдоль большей части полости тела, и специальные спинномозговые нервы. Значительная часть вегетативной активности контролируется спинным мозгом. Наивысшим центром, непосредственно связанным с вегетативной рецепцией и соответствующими двигательными ответами, является гипоталамус головного мозга. Таким образом, вегетативная система регулирует работу органов кровообращения, дыхания, пищеварения, выделения, размножения, процессы роста и обмена веществ. Рефлексы, затрагивающие обслуживаемые системой анатомические структуры, в большинстве своем носят характер самоуправления и не подчиняются произвольному контролю. Результатом работы вегетативной системы является обеспечение целостного реагирования организма на различные воздействия и поддержание уровня интенсивности основных жизненных процессов соответственно его текущим потребностям.
Организация вегетативных нервов спинного мозга отличается от ранее описанной для нервов, проводящих двигательные импульсы к поперечнополосатым мышцам (включая поперечнополосатые мышцы особой висцеральной системы головы) в одном примечательном отношении. Соматические двигательные импульсы проводятся от спинного мозга к органу-эффектору длинным аксоном единственного нейрона. Типичный вегетативный эфферентный путь, напротив, представляет собой цепь из двух нейронов. Первый, тело которого лежит в спинном мозге, именуется преганглионарным нейроном. Его аксон имеет типичную миелиновую оболочку. Однако эффектора этот аксон не достигает. В определенной точке цепи он входит в ганглий вегетативной системы, где двигательные импульсы передаются клеткам второго звена — постганглиоарным нейронам — аксоны которых (со слабо развитой миелиновой оболочкой или без нее) завершают путь до объекта иннервации (эффектора). В вегетативной системе единственным отклонением от двухнейронной схемы является особый способ иннервации мозгового вещества надпочечников.
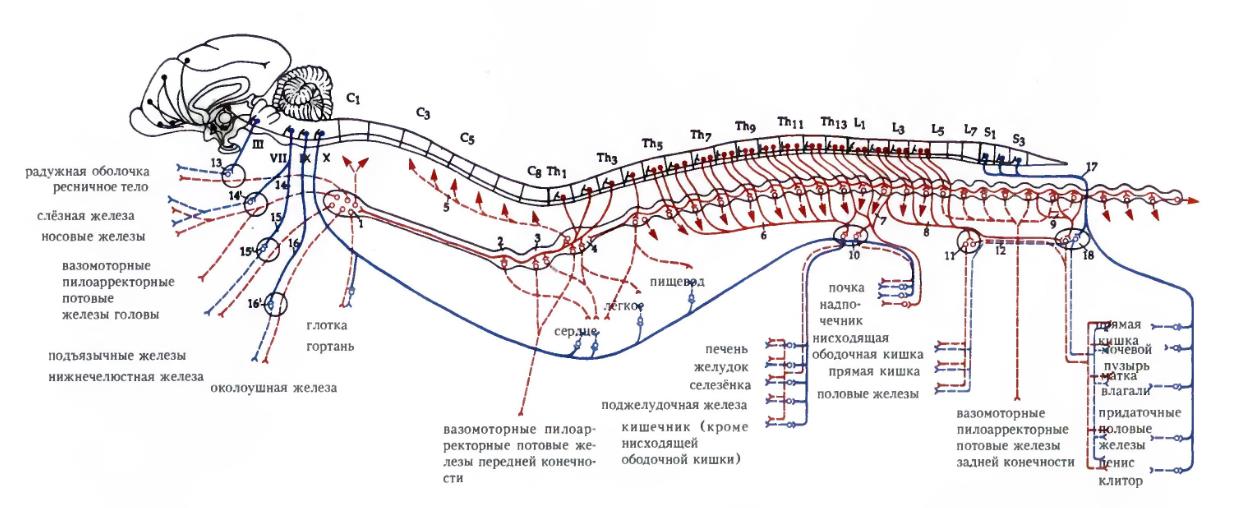
Схематическое изображение эфферентных симпатических и парасимпатических путей млекопитающего.
Объединение физиологических и анатомических данных дает основания для подразделения вегетативной системы на три части: симпатическую, или пояснично-грудную систему, парасимпатическую, или крестцово-черепную систему, а также метасимпатическую систему. Они отличаются как по своим функциям, так и по топографии. Как правило, большинство внутренних органов млекопитающих снабжено окончаниями двух, а то и всех трех означенных систем. Это является прогрессивной чертой; у рыб симпатическая и парасимпатическая системы не перекрываются, деля органы между собой.
Стимуляция собственно симпатических нервов оказывает возбуждающее влияние — ускоряет кровообращение, замедляет пищеварительные процессы, — т. е. повышает активность животного, подготавливая его к борьбе или к играм. Напротив, стимуляция парасимпатических нервов снижает общее возбуждение, но усиливает секрецию и перистальтику кишечника, благоприятствуя спокойной фазе существования. Совершенствуя регуляцию, подобный антагонизм значительно расширяет способность выносить угнетающие или чрезмерно возбуждающие внешние влияния (стресс), увеличивая шансы выживания организма в широком диапазоне условий. В то же время, некоторые органы (сосуды, потовые железы, мозговой слой надпочечников) находится под контролем только симпатической нервной системы.
В обоих системах кончиками нервных волокон выделяются нейромедиаторы. В собственно симпатической системе почти во всех случаях это два амина сходной структуры — норадреналин (норэпинефрин) и реже — адреналин (эпинефрин). Они почти идентичны веществам, которые выделяет в кровяное русло мозговое вещество надпочечников. В парасимпатической системе (как и в большинстве других отделов нервной системы) нейромедиатором служит ацетилхолин. Однако следует учитывать, что четкая картина химической и функциональной дифференцировки систем прослеживается не всегда, и резкого разграничения между ними зачастую нет.
Две части вегетативной системы млекопитающих можно анатомически можно различить по положению на нерве места переключения с преганглионарного волокна на постганглионарное и по положению на нервной трубке места выхода преганглионарного волокна. В симпатических нервах переключение, как правило, происходит рядом с центральной нервной системой, и соответственно преганглионарные волокна короткие, постганглионарные — длинные. В парасимпатической системе, напротив, преганглионарные волокна длинные, постганглионарные — короткие, а переключение происходит в обслуживаемом органе или рядом с ним.
Волокна симпатической системы выходят из спинного мозга в области I-го грудного – IV-го поясничного сегментов. Они прерываются в узлах пограничного симпатического ствола или в несколько дальше расположенных ганглиях, откуда распространяются по всему телу. Парасимпатические волокна выходят из среднего и продолговатого отделов головного мозга, включаясь в состав некоторых черепных нервов (в первую очередь, важного блуждающего нерва), а также из нескольких сегментов крестцового отдела. Волокна заканчиваются далеко на периферии в узлах у иннервируемого органа или внутри него.
У млекопитающих симпатические волокна выходят из позвоночного канала в составе главного ствола нерва каждого грудного и поясничного сегмента. Но сразу за пределами позвоночника они покидают этот ствол, поворачивают вентрально в составе короткой висцеральной, или белой соединительной ветви (ramus visceralis, или communicans albus), получившей такое название из-за ярко-белого цвета толстых миелиновых оболочек входящих в ее состав преганглионарных волокон, и входят в небольшой симпатический ганглий (ganglion sympathicum). У большинства млекопитающих эти ганглии с каждой стороны позвоночного столба соединены продольными пучками волокон, так что получается симпатическая цепь, или ствол (truncus sympathicus), вдоль которого преганглионарные волокна могут проходить некоторое расстояние до того сегмента, где они оканчиваются. В симпатических ганглиях импульсы, предназначенные для гладкой мускулатуры и желез кожи, передаются постганглионарным нейронам. У млекопитающих их периферические аксоны возвращаются из ганглия назад в главный ствол нерва в составе дополнительной соединительной ветви — серой (ramus communicans griseus), обязанной своим цветом и названием скудости миелина вокруг ее волокон.
Симпатические нервы, переключающиеся в ганглиях симпатического ствола, составляют лишь часть симпатической системы млекопитающих. Большинство из тех преганглионарных волокон, которые направляются к собственно внутренним органам, проходит сквозь эти ганглии (т. е. без переключения в них) и поворачивает вентромедиально. Под позвоночным столбом левые и правые преганглионарные симпатические волокна сплетаются в общую сеть, включающую постганглионарные клетки — это подпозвоночные ганглии (ganglia subvertebralia) или подпозвоночные сплетения (plexus subvertebrales), например чревное сплетение (plexus coeliacus), лежащее под передними поясничными позвонками, и расположенные позади него брыжеечные сплетения (plexus mesenterices). Здесь каждое преганглионарное волокно может передавать импульсы множеству постганглионарных клеток, длинные аксоны которых спускаются главным образом по брыжейкам к различным частям кишки.
Большая часть парасимпатических волокон распространяется в составе блуждающего нерва, главная ветвь которого следует вдоль значительной части длины пищеварительного тракта (благодаря чему этот нерв и получил название блуждающего); если не считать незначительного парасимпатического компонента других черепных нервов, остальные парасимпатические нервы берут начало в нескольких сегментах крестцового отдела. В этой системе нет проксимальных ганглиев; длинные миелинизированные преганглионарные волокна проходят все или почти все расстояние до различных внутренних органов; переключение происходит в сплетениях, обычно замурованных в обслуживаемом органе.
Наибольшей независимостью от центральной нервной системы отличается метасипатическая система. К ней относят комплекс микроганглиев и соединяющих их нервных волокон, расположенных в стенках внутренних органов, обладающих моторной активностью (пищеварительный тракт, сердце, мочеточники, матка и др.). Какие-либо ядерные образование в метасимпатической системе отсутствуют.
вперед
в оглавление