Внешний облик
Размеры ныне живущих однопроходных невелики: длина их тела колеблется от 30 до 80 см, вес — от 0,7 до 16,5 кг. Длина самых мелких ископаемых форм составляла приблизительно 15 см, тогда как наиболее крупные достигали около 1 м в длину и весили порядка 30 кг. Современные однопроходные самым неожиданным образом совмещают общую консервативность строения с признаками высокой специализации.
Голова небольшая, с вытянутым рылом, богатым нервными окончаниями. Мясистых губ нет. Глаза маленькие, наружные ушные раковины едва заметны или вовсе отсутствуют. Телосложение тяжелое. Конечности стопоходящие, короткие и сильные. Их передняя пара уступает по длине задней (утконос), либо они примерно одинаковой длины (ехидны). Широкие лапы с мощными когтями специализированы для копания почвы (утконос, ехидны) и плавания в воде (утконос). Конечности не подведены под тело, как у типичных териев (Theria), а расставлены в стороны подобно рептилиям. Голень со стопой повернуты назад постоянно (ехидны) или только во время плавания (утконос). Брюхо во время ходьбы касается субстрата (утконос) или расположено сравнительно высоко над ним (ехидны). Тело покрыто мягким густым мехом (утконос) или грубыми волосами и иглами (ехидны).
Железы внешней секреции
Имеются апокриновые и эккриновые потовые железы, но есть особенности их распределения внутри отряда. Млечные железы сохраняют простое трубчатое строение. Они представляют собой толстостенные извитые канальцы, которые ко времени лактации получают просветы, расширяются на концах и образуют тонкостенные альвеолы, продуцирующих молоко. В период выкармливания потомства железы могут достигать значительных размеров. Настоящих сосков нет и многочисленные выводные протоки желез открываются мелкими отверстиями на 2 плоских и покрытых волосками млечных полях по бокам брюха.
На пятке стопы находится экстратарзальная шпора. Она состоит из шпорной кости (os calcaris) и находящегося на ней кератинового шпорного рога (cornu calcaris), в которую выходит проток особой железы, расположенной позади бедра в месте его соединения с тазом (утконосы) или в области подколенной ямки (ехидны). По происхождению эти железы, вероятно, связаны с апокриновыми потовыми железами. У ныне живущих однопроходных функционирующие шпоры и железы присутствуют лишь у взрослых самцов; у самок шпорные кости отсутствует, а зачаточные роговые шпоры есть только в молодом возрасте. Утконос, железы которого вырабатывают ядовитый секрет, использует шпоры во внутривидовых поединках и для защиты от хищников. У ехидн шпоры имеют меньшие размеры, а секрет желез не ядовит и служит другим целям — предположительно, для запаховой коммуникации. Очень редко можно обнаружить одну или обе недействующие шпоры у взрослой самки.
Общий предок ехидны и утконоса, общий предок всех однопроходных и даже, возможно, общий предок однопроходных и териевых млекопитающих уже обладали какими-то ядом и ядопроводящим аппаратом. Учитывая наличие шпорной кости и костного шпорного рога у ряда мезозойских млекопитающих, можно считать, что экстратарзальная шпора была унаследована представителями данного класса от общего предка. Судя по всему, она развилась у него для защиты от хищников. У предков териев шпоры и яд были утрачены еще до разделения сумчатых и плацентарных. Присутствие рудиментарной шпоры у самок ехидны и утконоса означает, что яд и ядопроводящий аппарат у их дальних предков имелся у обоих полов. Вероятно половой диморфизм в отношении наличия ядовитой железы и ядопроводящего аппарата появился уже после отделения линии однопроходных от остальных млекопитающих. Яд был утрачен предками ехидны уже после разделения линий ехидн и утконосов.
Скелетная система
Череп уплощенный. Его лицевой отдел сильно вытянут, плоский и лопатообразно расширенный (утконосы) или цилиндрически заостренный (ехидны). Мозговой отдел довольно больших размеров, круглый и гладкий. В отличие от териев, кости черепа срастаются друг с другом очень рано, практически не оставляя разделительных швов. Хрящевой череп и соотношение костей в крыше и основании костного черепа в известной степени сходны с таковыми у рептилий.
Удлиненная морда образована вытянутыми верхнечелюстными и предчелюстными костями. Для носовых костей тоже характерны большие размеры. Хорошо развит сошник, который идет назад к основной части клиновидной кости. Подобно цинодонтам и млекопитающеобразным, но в отличие от териев (кроме ксенартр), имеется крупная септомаксилла, а предчелюстная кость несет отросток (processus ascendus), сходный с таковым у терапсид. У утконосов также присутствует непарная гантелевидная кость (paradoxum), которая по различным предположениям может являться рудиментом предсошников, небных отростков межчелюстных костей или уникальным новообразованием. Вдоль костей морды открывается несколько небольших отверстий для прохождения ветвей V- черепного (тройничного) нерва для обслуживания чувствительного клюва или рыльца.
Подобно прочим млекопитающим, свод ротовой полости сформирован соединенными друг с другом широкими и плоскими элементами, начиная от парных верхнечелюстных и небных костей и заканчивая направленным вниз гребнем сошника. Они формируют парные носоглоточные проходы и сильно протяженное вторичное костное небо. Спереди носоглоточные проходы открываются наружными ноздрями, а в ротовую полость — хоанами, которые отодвинуты далеко назад длинными пластинками небных костей, лишенными в задних отделах мелких отверстий. Сбоку и позади каждой небной кости расположена крыловидная кость, которая перекрывается массивным эктоптеригоидом, также характерным для терапсид, но отсутствующим в виде отдельного элемента у взрослых териев. В отличие от териев и подобно другим нетериевым млекопитающим и млекопитающеобразным, крылоклиновидная кость относительно мала, а передняя пластина каменистой кости очень велика. Присутствует основная крылоклиновидная кость (alisphenoideum), тоже имеющая своеобразное строение.
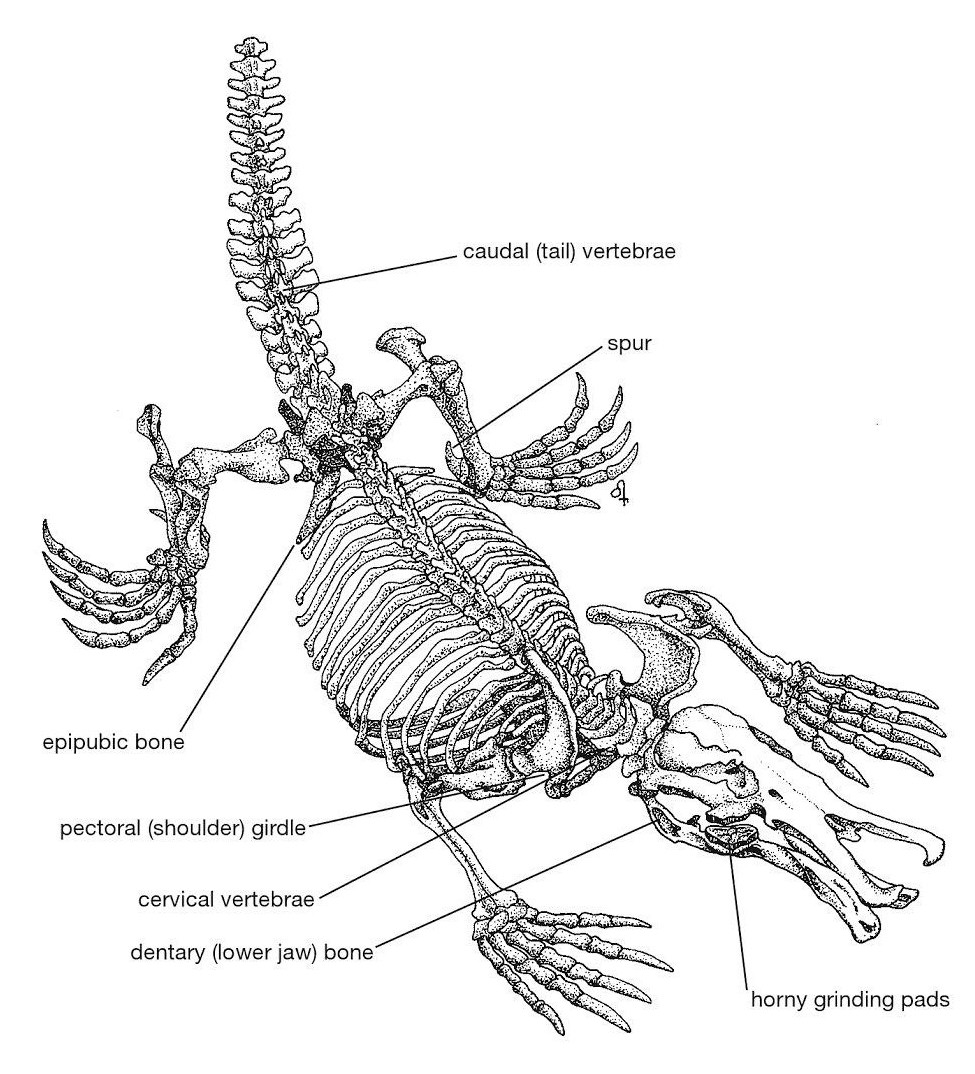
Прорисовка скелета утконоса (Ornithorhynchus anatinus).
Передняя стенка черепной коробки образована поперечной мезетмоидальной костью (mesethmoideum), носовыми капсулами и lamina infracribrosa, которая сверху несет простирающуюся далеко вперед горизонтальную продырявленную пластинку (lamina cribrosa). Подобно териям, у ехидн эта пластинка пронизана множеством мелких отверстий для прохождения нервных волокон от обонятельного эпителия носовых раковин к обонятельным луковицам мозга. Напротив, у утконоса с каждой стороны в продырявленной пластинке имеется только по 1 отверстию, что отличает его от всех других современных млекопитающих. В отличие от большинства териев, слезная кость отсутствует. Глазоклиновидная кость (orbitosphenoideum) большая. С каждой стороны имеются чешуйчатая и каменисто-сосцевидная кости. Крышу черепа образуют сильно укороченные лобные, а также теменные кости. В отличие от териев, сохраняются рудименты предлобных и заднелобных костей.
Основание черепной коробки уплощено. Снизу в ее состав входит комплекс, включающий сросшиеся основную часть клиновидной кости и пресфеноид, а также pilae antoticae, проходящие вдоль боков турецкого седла. Pilae antoticae представляют собой парные полоски хряща, развивающиеся из основания хрящевого черепа и затем окостеневающие. У рептилий они формируют плевросфеноидные кости (pleurosphenoideum), образующие значительную часть черепной коробки, а у однопроходных имеют рудиментарный характер. У териев pilae antoticae отсутствуют. Спереди от базисфеноидно-пресфеноидного комплекса находится сошник, а позади него — основная затылочная кость. Задняя часть черепа содержит парные боковые и непарную верхнюю затылочную кость, окружающие большое затылочное отверстие. Каждая боковая затылочная кость несет мыщелок, сочленяющийся с I-м шейным позвонком.
Барабанная полость расположена на задне-нижней поверхности каменисто-сосцевидной кости, лежащей между чешуйчатой и основной затылочной костями. Барабанная кость в форме кольца (утконос) или полукольцевидная (ехидны), относительно слабо соединенная с окружающими костями черепа (не сращена с к каменистым отделом височной кости черепа (petrosum), как у большинства териев). У утконосов барабанная полость снизу полностью открыта, у ехидн углубление для среднего уха частично прикрыто продолжениями эктоптеригоида и каменистой кости. В обоих случаях костный слуховой барабан не образуется. В отличие от териев, костный слуховой проход не сформирован.
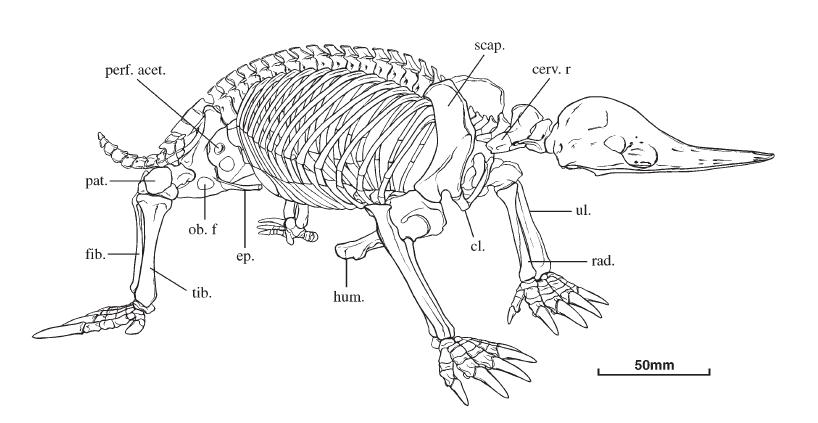
Прорисовка скелета австралийской ехидны (Tachyglossus aculeatus): cerv. r. — шейное ребро; cl. — ключица; ep. — предлобковая кость; fib. — малая берцовая кость; hum. — плечевая кость; ob. f. — запирательное отверстие; pat. — коленная чашечка; perf. acet. — отверстие вертлужной впадины; rad. — лучевая кость; scap. — лопатка; tib. — большая берцовая кость; ul. — локтевая кость.
Подобно всем настоящим млекопитающим, звукопроводящий аппарат среднего уха состоит из 3 слуховых косточек — молоточка, наковальни и стремени. В отличие от териев, молоточек и наковальня плотно соединяются (утконос) или даже срастаются (ехидны) и имеют длинный отросток (processus folii), жестко прикрепленный к каменистой кости. Молоточек большой относительно наковальни. У ехидн молоточек посредством отростка (processus gracilis) плотно присоединяется, а часто даже срастается с барабанной костью. Стремя крупное, столбикообразное, без отверстия. Близкая к столбикообразной форма стремени и отсутствие в нем отверстия отмечены также у ряда териев (например, у сумчатых и панголинов). Слуховые косточки полностью включены в череп, а не входят в состав нижней челюсти, как это наблюдается у цинодонтов и млекопитающеобразных. Судя по палеонтологическим данным (в частности, ископаемым остаткам Teinolophos), конструкция из 3 нижнечелюстных косточек была вовлечена в передачу звука до разделения на предков однопроходных и териев (Theria), но независимо в обеих линиях сформировала полностью отделенное от челюсти среднее ухо (явление гомоплазии).
Примечательной особенностью уха однопроходных является то, что овальное окно у них круглое по форме, а круглое окно — овальное. Затянутое перепонкой овальное окно расположено в верхней части куполовидной барабанной полости. К этой перепонке прикреплено расширенное круглое (архаичная особенность) основание стремени. Барабанная перепонка с прилегающей к ней рукояткой молоточка остаются видимыми при взгляде снизу. Наружное слуховое отверстие расположено у основания нижней челюсти.
Тонкая скуловая дуга образована длинным скуловым отростком верхнечелюстной кости, доходящим до суставной впадины для нижней челюсти, и скуловым отростком чешуйчатой кости. Скуловая кость у взрослых современных однопроходных отсутствует; у молодого утконоса имеется очень маленькая скуловая косточка, прилегающая к скуловой дуге сверху. Основание скулового отростка чешуйчатой кости пронизано широким височным каналом (рудимент зачешуйчатого отверстия терапсид, также встречающийся у некоторых архаичных млекопитающих) для затылочной артерии. Сочленовная ямка для нижней челюсти образована чешуйчатой костью. Подобно прочим млекопитающим, нижняя челюсть образована лишь парной зубной костью. Спереди обе ее ветви неподвижно сращены друг с другом.
Нижняя челюсть в значительной мере редуцирована. Ее венечный и угловой отростки выражены слабо или полностью отсутствуют. В отличие от сумчатых, угловой отросток не загнут внутрь. У утконосов имеется внутренний челюстно-подъязычный отросток, расположенный чуть выше нижнечелюстного отверстия. Внутри зубной кости пролегает канал для прохождения нервов — расширенный (утконосы) или узкий (ехидны).

Жевательные поверхности правого верхнего предкоренного зуба однопроходных, с соблюдением масштаба: A — Stirtodon elizabethae; B — Kollikodon ritchei; C — Teinolophos trusleri; D — Obdurodon dicksoni; e — гипотетический предкоренной Obdurodon tharalkooschild.
У взрослых особей ныне живущих видов зубы совершенно отсутствуют. У современного молодого утконоса (Ornithorhynchus) с каждой стороны имеется 2 верхних и 3 нижних рудиментарных щечных зуба — низких, широких, с многочисленными неправильно расположенными бугорками. Вскоре они утрачиваются и заменяются роговыми пластинками. Древние утконосы (Steropodon, Monotrematum, Obdurodon) на протяжение всей жизни имели щечные зубы с прямоугольной жевательной поверхностью и с 4-5 разными по высоте бугорками, образующих 2 хорошо выраженные поперечные режущие лопасти, разделенные глубокой выемкой.
Для обозначения характерных элементов жевательной поверхности озубленных однопроходных разработана альтернативная номенклатура. Согласно ей, бугорок верхнего зуба называют акисом, а его лезвие — дрепаном, тогда как на нижнем зубе они именуются соответственно акидом и дрепанидом. Триакидидрепаноны (триакидидрепаниды на нижних зубах) формируют режущие лопасти, состоящие из 3 бугорков, V-образно соединенных 2 гребнями. Диакидрепаноны (диакидрепаниды на нижних зубах) состоят из 2 бугорков, соединенных единственным гребнем. Между режущими лопастями расположена широкая и глубокая долинка для вхождения аналогичной структуры противостоящего зуба.
Зубы ископаемых и молодого ныне живущего утконоса отдаленно напоминают щечные зубы гарамиевых (Haramiyidae) и многобугорчатых (Multituberculata), но в то же время сходны с трибосфеническими зубами более продвинутых групп, таких как метатерии (Metatheria) и эвтерии (Eutheria). Окончательно неясно, приобретена ли такая форма зубов однопроходными независимо (что делает их зубы псевдотрибосфеническими), или была унаследована от общего с териями предка с примитивными трибосфеническими зубами. Возможно, утрата зубов у однопроходных была связана с развитием электрорецепции.
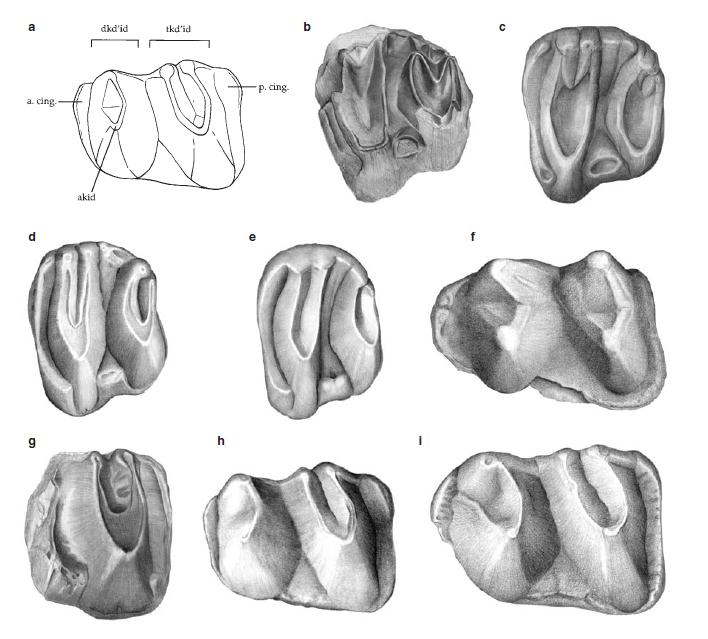
Прорисовки жевательных поверхностей коренных зубов утконосов, без соблюдения масштаба: a — схема левого m1 Obdurodon insignis, иллюстрирующая строение режущих лопастей; b — m2 Monotrematum sudamericanum; c — m2 ‘Obdurodon sp. A’; d — m2 Obdurodon insignis; e — m2 Obdurodon dicksoni; f — Steropodon galmani; g — задняя половина m1 Monotrematum sudamericanum; h — m1 Obdurodon insignis; i — m1 Obdurodon dicksoni. Сокращения: dkd’id — диакидрепанид (одиночное лезвие); tkd’id — триакидрепанид (V-образное лезвие); a. cing. — передний цингулюм; p. cing. — задний цингулюм.
Посткраниальный скелет утконоса и ехидн имеет много общих черт строения. В большинстве своем они являются плезиоморфными, но в обоих случаях помимо этого отчетливо просматривается специализация к рытью. Утконос — более легко сложенное животное с рядом специализаций к водному образу жизни (что, в частности, выражено в уплощенной, обтекаемой форме тела), тогда как ехидны — массивно сложенные копатели.
Позвоночный столб состоит из 7 шейных, 15-17 грудных, 2-3 поясничных, 2 крестцовых, 0-2 копчиковых и 11-20 хвостовых позвонков. Как и у других амниот, 2 первых шейных позвонка значительно специализированы. Атлант имеет форму кольца, его передняя поверхность вогнута для сочленения с затылочными мыщелками черепа. Эпистрофей на переднем конце несет зубовидный отросток (processus odontoideus), представляющий собой очень поздно прирастающее тело атланта. Этот отросток входит сзади в кольцо атланта. Атланто-затылочный сустав позволяет совершать сгибательно-разгибательные движения, а атланто-аксиальное сочленение допускает вращение. Эпифизы на телах позвонков имеются только в хвостовом отделе. Сочленения туловищных позвонков напоминают таковые у продвинутых цинодонтов.
В противоположность териям, ребра лишены головок и бугорков, а присоединяются непосредственно к телам позвонков. В отличие от териев и подобно многим базальным группам, имеются рудиментарные шейные ребра. В молодом возрасте они не слиты с позвонками, но затем с срастаются с их поперечными отростками, все же оставаясь заметными. В результате срастания с каждой стороны позвонка формируется поперечное отверстие, совокупность которых образует канал для прохождения позвоночной артерии, вены и нервного сплетения. Грудные ребра большие и плоские. В противоположность териям и подобно птицам, реберные хрящи, соединяющие истинные ребра с грудиной и ложные ребра с последними истинными, полностью окостеневают. Подобно ряду древних млекопитающих, ребра имеются и в поясничном отделе.
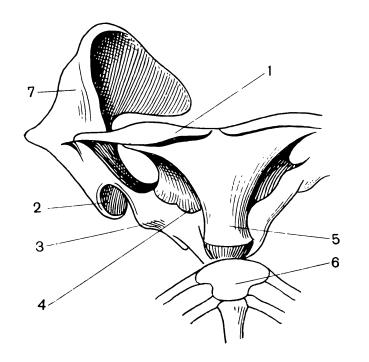
Прорисовка плечевого пояса утконоса (Ornithorhynchus anatinus), снизу: 1 — ключица; 2 — сочленовная ямка для головки плечевой кости; 3 — метакоракоид; 4 — прокоракоид; 5 — межключица; 6 — грудина; 7 — лопатка.
Плечевой пояс прикреплен к осевому скелету гораздо жестче, чем у териев. Его строение сходно с таковым у продвинутых цинодонтов и млекопитающеобразных, но некоторые элементы (кости грудины) уже достигают характерного для териевого уровня развития. В плечевом поясе сохраняются прокоракоид и метакоракоид — самостоятельные столбчатые кости. Они представляют собой преобразованные передний и задний коракоиды терапсид, в ходе эволюции утраченные большинством ветвей млекопитающих. У современных териев они редуцируются в эмбриональном возрасте, причем рудимент метакоракоида прирастает к лопатке, образуя коракоидный отросток, а прокоракоид полностью исчезает. Ключица очень крупная. Между ключицами и грудиной продолжает существовать большое Т-образное срединное окостенение — межключица, также утрачиваемая териями. У ехидн имеется мечевидный отросток грудины, тогда как у утконоса он отсутствует.
Лопатка расширяется кверху, а спереди несет акромиальный отросток. Она сохраняет много плезиоморфных черт. Так, лопаточная ость отсутствует либо зачаточна. Нет также предостной ямки, которая у териев расположена впереди ости и образует передний край лопатки. Есть подобие небольшой заостной ямки. Предостная ямка териев служит для прикрепления m. supraspinatus, стабилизирующей положение плеча во время ходьбы. Снизу лопатка сочленена с прокоракоидом и метакоракоидом. Сверху этот блок прикрепляется к грудной клетке, а снизу — к межключице, которая сзади сочленена с грудиной. Поперечная часть межключицы плотно зажата между лопатками и ключицами. Суставная впадина лопатки представляет собой ориентированную латерально клиновидную впадину в месте сочленения лопатки и метакоракоида.
Громоздкий плечевой пояс однопроходных в известной мере ограничивает подвижность передних конечностей и делает невозможной асимметричную походку. В то же время, жесткий плечевой пояс обеспечивает конечностям надежную опору во время плавания и рытья, а обширные костные площади служат для прикрепления мощной грудной мускулатуры. Возможно, именно из-за этой необходимости плечевой пояс не только не утратил большую часть своих элементов, как это произошло у териев, но даже стал более массивным (в особенности, у ехидн), чем был у многих мезозойских млекопитающих (например, у эвтриконодонтов (Eutriconodonta) и многобугорчатых (Multituberculata)).
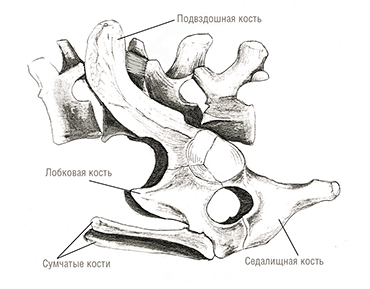
Прорисовка тазового пояса утконоса (Ornithorhynchus anatinus).
Тазовый пояс построен менее архаично, но тоже сохраняет ряд примитивных черт, демонстрируя в то же время некоторые специализации. Короткий и широкий таз состоит из 3 парных костей — верхней подвздошной, нижне-передней лобковой и нижне-задней седалищной. Подобно некоторым цинодонтам, лопасть подвздошной кости направлена вперед и вверх. Лобковая и седалищная кости разделены сверху запирательным отверстием и соединены снизу сильно удлиненным симфизным сращением. В вертлужной впадине сращение 3 костей неполное и имеется отверстие — по-видимому, это связано с какой-то локомоторной специализацией. У представителей обоих полов к каждой лобковой кости спереди подвижно причленяется длинная, палочковидная предлобковая кость (prepubica). Эти кости также имеются у некоторых продвинутых цинодонтов, многих базальных млекопитающих, большинства метатериев, ряда базальных эвтериев и служат у них для прикрепления мышц бедер и брюха. Передняя часть подвздошной кости удлинена, сужена и проходит вперед и вверх, где сочленяется с 2 крестцовыми позвонками.
Подобно цинодонтам и примитивнейшим мезозойским млекопитающим, конечности относительно короткие, направленные более или менее в стороны от туловища. Плечевая кость отстоит от средней линии тела не менее чем на 45º, так что кисть контактирует с субстратом сбоку от плечевого пояса. Размещение конечностей под таким углом приводит к более рептильному способу ходьбы, когда тело волнообразно выгибается из стороны в сторону. Напротив, у многих мезозойских и всех современных млекопитающих (за исключением специализированных) конечности находятся ближе к средней линии тела (в парасагиттальной позиции), что способствует более быстрой, «маммальной» походке. По-видимому, латеральное положение конечностей у однопроходных является следствием их специализации к рытью нор, поскольку подобное положение также характерно для ныне живущих териев с подземным образом жизни.
Плечевая кость короткая, очень массивная, скрученная вокруг оси и с внутренним надмыщелковым отверстием. Некоторые черты ее строения общие с цинодонтами, включая форму головки, взаиморасположения гребней на проксимальном конце и очертания выемки для прикрепления двуглавой мышцы. Дистальный конец расширен наподобие того, как это наблюдается у терапсид и некоторых мезозойских млекопитающих, но не как у териев (за исключением специализированных землероев типа кротов). Этот исключительно широкий дистальный конец плечевой кости является областью прикрепления сильной мускулатуры передней конечности. Локтевая кость значительно длиннее лучевой. Ее расширенный проксимальный конец соединяется с плечом с помощью отростка. Эта структура, образовавшаяся из спиралевидного локтевого отростка цинодонтов, больше всего напоминает локоть некоторых юрских млекопитающих. У териев развилась выемка между отростками локтевой и лучевой костей, формирующая блок плечевой кости (указание на парасагиттальное положение передних конечностей). Запястье однопроходных короткое и широкое. Ладьевидная и полулунная кости запястья сращены, центральной кости нет.
Короткая и мощная бедренная кость демонстрирует ряд архаических особенностей: головка не отклоняется к средней линии тела, шейка очень короткая, вертлуги расходятся в сторону, III-й вертлуг отсутствует. В целом бедро по строению напоминает таковое у прогрессивных цинодонтов и млекопитающих позднего триаса - юры, у которых задняя нога еще не была подведена под тело. У предков териев бедро претерпело ряд изменений, в результате которых конечность приняла парасагиттальное положение: головка отклонилась внутрь, развилась шейка, вертлуги сместились для прикрепления преобразовавшихся мышц. Бедренная кость утконоса по форме архаичнее, чем у ехидн. Коленный сустав однопроходных имеет несколько приобретенных ими черт, включая широкую выемку для коленной чашечки и разделенные, плоские, равные по величине мыщелки на бедренной кости.
Малоберцовая кость скручена назад, что частично объясняет задне-боковую ориентацию стопы. На уплощенном проксимальном конце малоберцовой кости располагается направленный вверх длинный отросток (perconecranon), служащий для прикрепления мышц; у утконоса он уплощен и удлинен особенно значительно. У ехидн большая и малая берцовые кости обращены назад, так что задняя конечность вместе со стопой повернута в наружно-заднем направлении. В строении предплюсны сочетаются черты плезиоморфности и высокой специализации не-териевого типа. Подобно некоторым мезозойским млекопитающим и млекопитающеобразным, голеностопный сустав содержит у самцов шпорную кость. Все конечности стопоходящие, широкие лапы изначально пятипалые. У ехидн I-й и V-й пальцы на передних и задних лапах могут редуцироваться. Дистальные фаланги когтевидные, очень крупные. Пальцы оканчиваются длинными когтями.
В локомоции отмечается уникальная механика движения, отличающаяся от таковой у других тетрапод. Широкая суставная впадина лопатки принимает крупную головку плечевой кости, расположенной почти горизонтально и отведенной под углом 90° к парасагиттальной плоскости тела. Лучевая и локтевая кости причленяются к мыщелку плечевой кости, имеющему квазиспиральную конфигурацию. Согласно результатам кинорадиографического анализа, основным локомоторным движением плеча является ротация (т. е. вращение) вокруг продольной оси, а не антеропостериорная протракция (движения вперед и назад, наблюдаемые у териев с парасагиттальным положением конечностей). Подошвенный контакт кисти с субстратом осуществляется под суставной впадиной лопатки — подобно тому, как это происходит у типичных териев.
Бедро также занимает практически горизонтальное положение и расположено по отношению к туловищу под прямым углом. Главные локомоторные движение бедра — это ротация вокруг проксимодистальной оси (от головки до паза для надколенника) и, в меньшей степени, элевация и депрессия; полная ротация составляет приблизительно в 45°. Длинная ось бедра во время движения также смещается назад, начиная примерно с 35° в сагиттальной плоскости и заканчивая на 50° или более. При этом в момент финального отталкивающего движения ступня направлена назад. Ретракция бедра — один из важнейших компонентов локомоции для большинства тетрапод. У клювоголовых (Sphenodontida), чешуйчатых (Squamata) и крокодилов (Crocodilia) она превышает 70°, а у териев (Theria) — 50-70°. В то же время, у однопроходных во время наземного передвижения ретракция не превышает 15°, замещаясь пронацией.
Основные различия в движении задних конечностей однопроходных следующие: во время движения бедро утконоса расположено в сагиттальной плоскости под большим углом, чем у ехидны и, кроме того, дистальный конец бедренной кости утконоса находится значительно более дистально относительно вертлужной впадины. Дистальный конец бедренной кости ехидны в начале движения находится немного выше, чем вертлужная впадина, а в конце движения оказывается ниже. Таким образом, при наземной локомоции туловище ехидны находится заметно выше над субстратом, чем у утконоса.
Задняя конечность ехидны имеет специфическое строение — результат ее приспособленности к копанию. Это животное способно зарываться в субстрат с удивительной скоростью, используя задние и передние конечности, вооруженные сильными кривыми когтями. При этом кисти быстро вращаются в различных направлениях вокруг вертикальной оси. Бедро ехидны сглажено и округлая головка бедренной кости расположена несколько медиально, содействуя большей пронации. Таким образом, ось вращения бедра расположена близко к среднему краю кости. Пронация также усиливается приводящими мускулами аддуктора удлиненного медиального мыщелка.
У ехидны вся подошвенная поверхность стопы находится в контакте с поверхностью с начала до конца фазы движения. Кроме того, сферическая поверхность мыщелка ее лодыжки обеспечивает изменчивость движений конечностей во время ходьбы и копания. Не последнюю роль здесь играет и пяточный бугор. Его удлинение обычно связывали с усилением флексии голеностопного сустава. Считается, что удлинение пяточного бугра первоначально было связано с абдукцией в голеностопном суставе, что помогало противостоять скольжению в сторону при симметрично-диагональных походках у животных с широко расставленными конечностями. Пяточный бугор разрастался у предков млекопитающих (так же как аналогичное образование у крокодилов), увеличивая нагрузку на боковые пальцы. При наблюдаемой в эволюции млекопитающих тенденции к использованию асимметричных походок, первичная функция пяточного бугра постепенно менялась в сторону увеличения роли флексии голеностопного сустава, одновременно с переводом конечностей в парасагиттальное положение. Существует несомненная связь между ролью пяточного бугра и пятой плюсневой у однопроходных, так же как и у многих продвинутых цинодонтов.
Мышечная система
Мускулатура весьма своеобразна, отражая архаичность и специализацию скелетной системы. Жевательные мышцы развиты слабо. У утконоса нижнюю челюсть опускает парная m. detrahens mandibulae, с каждой стороны прикрепляющаяся к нижней части ее внутренней поверхности. Она иннервируется ветвью тройничного (V-го черепного) нерва и является производным жевательной мышцы (m. masseter), сместившейся назад к челюстному суставу и взявшей на себя функцию открывания рта. У ехидн m. detrahens mandibulae практически не функционирует, а рот открывается и закрывается вследствие вращения зубных костей относительно их продольных осей. M. detrahens mandibulae однопроходных может являться пережитком той давней поры, когда функция нижнечелюстного сустава переходила от сочленовной и квадратной костей к зубной и чешуйчатой костям.
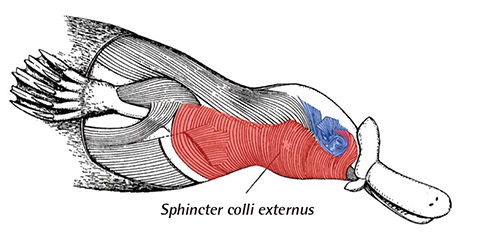
Мышцы передней части тела утконоса (Ornithorhynchus anatinus).
Имеются предшественники мимических мышц, берущие начало из общего сжимателя шеи (m. sphincter colli). Этот мускул присутствует уже у рептилий и птиц, но ограничен у них областью шеи, так что их лицо представляет собой лишенную выражения маску. У однопроходных сжиматель шеи вторгается в лицевую область и дифференцируется в подкожную мышцу, проходящую сбоку головы и прилегающую к ушному отверстию, лобную подкожную мышцу вокруг глаз и боковые щечные мышцы, расположенные под нижней челюстью. Задняя часть сжимателя шеи прикрепляется к начальным отделам грудной клетки и плеча. Сзади от верхних и боковых поверхностей черепной коробки к плечевым костям тянутся широкие, напоминающие ленты мышцы, особенно выраженные у ехидн и придающие им «безшеий» облик. Эта специализированная мускулатура существенно отличается от поверхностной мускулатуры териев.
Туловище покрыто слоем сильно развитой подкожной мышцы (panniculus carnosus). Подкожная мускулатура не развита лишь в районе конечностей, млечных желез, клоаки и хвоста. Мускулатура плечевого пояса почти наверняка близка по строению к грудной мускулатуре цинодонтов и базальных млекопитающих, у которых плечевой пояс был построен сходным образом. Очевидна специализация мышц к копанию. Некоторые мышцы, по-видимому, не изменились со времен терапсид. Уникальное расположение мускулатуры затрудняет гомологизацию мышц плечевого пояса однопроходных с ныне живущими териями и предполагает длительную независимую эволюцию. Гортань примитивна и лишена голосовых связок.