Нервная система
Головной мозг в общем крупный и обладает многими характерными для териев чертами, но сохраняет при этом ряд примитивных признаков. Обонятельные луковицы относительно небольшие (утконос) или очень крупные (ехидны); они не имеют слоя митральных клеток, характерного для териев. Обонятельные пути головного мозга в целом сходны с таковыми у других представителей класса. Подобно териям, наиболее развитой частью больших полушарий является кора, а не полосатое тело, как у рептилий и птиц.
Новая кора очень обширна по сравнению с древней корой, или обонятельной долей, и старой корой, или гиппокампом. В целом новая кора занимает 40-50 % поверхности головного мозга. В месте, где оттесненная вниз обонятельная доля встречается с новой корой, находится хорошо выраженная продольная, или сильвиева, борозда. Снаружи новая кора покрыта довольно многочисленными извилинами (ехидны) или, напротив, полностью их лишена (утконос). Примечательно, что у ехидн рисунок извилин отличается от рисунка, характерного для териевых млекопитающих. Внутренняя структура коры примитивна. Подобно рептилиям и сумчатым, отсутствует мозолистое тело, которое у плацентарных является основным маршрутом коммуникации между правым и левым полушариями. Имеются только спайка свода (commissura fornicis) и передняя спайка (commissura anterior). У однопроходных и сумчатых передняя спайка содержит все спайковые волокна, идущие из новой коры, тогда как у плацентарных она включает только некоторые из этих волокон.
Как и у териев, на крыше среднего мозга расположено четверохолмие (corpora quadrigemina), в отличие от двухолмия (corpora bigemina) большинства рептилий. Мозжечок прикрыт большими полушариями лишь частично. Он имеет крупные размеры и разделяется на ряд долей, поверхность которых покрыта складками. Боковые части мозжечка соединены друг с другом варолиевым мостом — массивной полосой проходящих снизу нервных волокон, имеющейся только у млекопитающих. Характерной особенностью однопроходных являются очень крупные тройничные нервы. Это смешанные чувствительно-двигательные нервы, но их чувствительные компоненты развиты значительно больше, поскольку проводят импульсы от многочисленных рецепторов морды к коре через перераспределяющий их таламус. Подобно териям и в отличие от всех других позвоночных, связь между двигательной областью новой коры и спинным мозгом осуществляет пирамидный путь. Спящий мозг поочередно проходит через фазы медленного и быстрого сна.
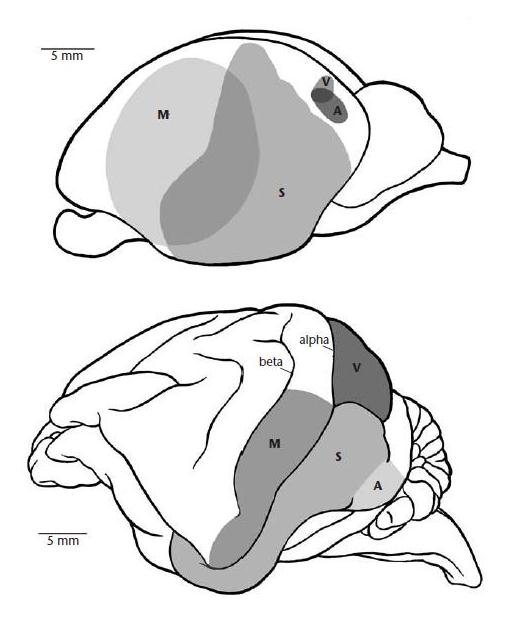
Вид слева на головной мозг утконоса (Ornithorhynchus anatinus) (вверху) и австралийской ехидны (Tachyglossus aculeatus) (внизу) (по Augee et Gooden, 1997):
А — слуховая область коры; М — двигательная область коры; S — сенсорная область коры; V — зрительная область коры, alpha — альфа-извилина; beta — бета-извилина.
Органы чувств
Обоняние развито достаточно хорошо у утконоса и превосходно у ехидн. Имеется вомероназальный орган, открывающийся в ротовую полость. По сторонам верхней части языка расположены углубления со скоплениями вкусовых луковиц.
Слух посредственный (утконос) или хороший (ехидны). Строение органов слуха архаично. Ушная раковина отсутствует или очень невелика. Состоящий из хрящевых колец изогнутый наружный слуховой проход проводит звуковые волны от ушного отверстия к барабанной перепонке в основании нижней челюсти. Изнутри к барабанной перепонке примыкают слуховые косточки. Вследствие плотного соединения между собой и с прилежащей стенкой черепа они могут вибрировать только как один элемент и практически не приспособлены для восприятия воздушных звуковых колебаний. С другой стороны, внутреннее ухо хорошо улавливают вибрации, проводимые телом. Подобно мезозойским группам млекопитающих, ушная улитка завита сравнительно слабо — примерно на 270°; у современных териев изгиб составляет 360º. Судя по всему, эта извитость развилась у однопроходных независимо. Внутри канала улитки отсутствуют костные пластинки, имеющиеся у териев. В остальном внутреннее строение улитки (включая устройство кортиева органа) вполне сходно (хотя и не идентично) с таковым у териев, в то же время достаточно сильно отличаясь от рептильного. В отличие от типичных териев, ухо наиболее чувствительно к низкочастотным звукам (примерно 4-5 кГц).
Вестибулярный аппарат развит хорошо. Как и у териев, он представлен ампулами 3 полукружных каналов и макулами овального и круглого мешочков. Кроме того, подобно рептилиям и в отличие от териев имеется макула улитки.
Зрение слабое. Для него характерно необычное сочетание черт, присущих как териям, так и рептилиям. Глазное яблоко маленькое, сферической формы. В задней части склеры залегает чашевидное образование из хряща — структурный элемент, не встречающийся у териев, но обычный для части рептилий. Полагают, что его функция — укрепление глазного яблока, особенно во время аккомодации. Сосудистая оболочка толстая тонкая. Хрусталик сильно уплощен. В отличие от териев, отсутствует ресничная мышца, влияющая на кривизну хрусталика. Высказано предположение, что для обеспечения фокусировки зрения (весьма ограниченной) с помощью окружающих мышц изменяет форму все глазное яблоко. Сетчатка лишена сосудов. Подобно другим позвоночным, она состоит из тонкого слоя нервных волокон, слоя ганглионарных клеток и рецепторного слоя. Содержащиеся в последнем двойные колбочки заключают внутри капельки жира. Такое строение колбочек не встречается у териев, но присуще рептилиям. В оптической хиазме перекрещивается большинство зрительных нервных волокон; лишь малая их часть продолжает идти по той же стороне мозга. Мигательная перепонка имеется (утконосы) или отсутствует (ехидны).
В толще кожи на морде у всех современных видов располагаются высокочувствительные электрорецепторы, которые среди прочих млекопитающих пока обнаружены лишь у гвианского дельфина (Sotalia guianensis). Считается, что у однопроходных они представляют собой специализированные эккриновые железы, выводные протоки которых снабжены чувствительными нервными окончаниями. Этот вид чувствительности позволяет выявлять жертву, слабые электрические поля, вызванные ее мышечной активностью. При этом наибольшее число электрорецепторов характерно для утконоса (40 000), охотящегося в водной среде. Меньшим их количеством обладают проехидны (2000), которые роются во влажной лесной подстилке. Наконец, меньше всего рецепторов у австралийской ехидны (400), обитающей главным образом в засушливых условиях. Также на морде однопроходных всегда присутствуют несенсорные эккриновые железы, выделяющие слизистый (утконос) или серозно-слизистый (ехидны) секрет. В отличие от териев, вибриссы совершенно отсутствуют.
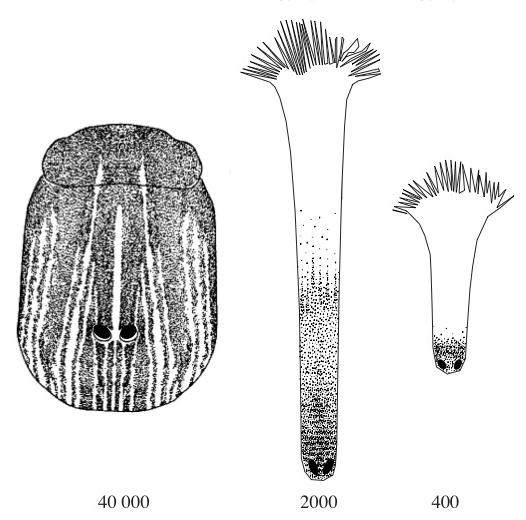
Схема расположения электрорецепторов (обозначены точками) на морде утконоса (Ornithorhynchus anatinus), проехидны Брюйна (Zaglossus bruijnii) и австралийской ехидны (Tachyglossus aculeatus).
Пищеварительная система
Слюнные железы маленькие (утконос) или сильно увеличенные (ехидны). Желудок простой, без пищеварительных желез, что является единственным случаем у современных млекопитающих. У утконоса его функция заключается в хранении пищи аналогично зобу птиц, а у ехидн стенки желудка выстланы ороговевающим эпителием и дополнительно служат для перетирания корма. Пищеварительный тракт делится на тонкую и толстую кишки, но отчетливо выраженные различия между ними отсутствуют. Имеется очень короткая слепая кишка. У самцов и самок есть клоака — карман, объединяющий выходы из пищеварительной, выделительной и половой систем. По сути, она представлена лишь задним отделом, примерно соответствующим проктодеуму рептилий. Передний отдел клоаки разделен на крупный верхний проход — копродеум, куда открывается прямая кишка, и нижнюю часть — уродеум, формирующую мочеполовой синус. Печень многодольчатая, с желчным пузырем.
Выделительная система
Почки по строению напоминают таковые у прочих млекопитающих и лишены выраженного сходства с дольчатыми почечными органами рептилий. В отличие от последних, отсутствует воротная система почек; напротив, как и у других млекопитающих, насыщенная кислородом кровь поступает в почку через почечную артерию и отводится затем по почечной вене. В то же время, характерно наличие т. н. карликовых нефронов без длинных петель Генле, обнаруженных также в почках многих рептилий. Мочеточники порознь открываются в передний конец мочеполового синуса, а напротив них по средней линии нижней поверхности синуса открывается мочевой пузырь. Благодаря такому их взаиморасположению моча поступает в пузырь, не смешиваясь с фекалиями. И у самцов и у самок сокращение мочевого пузыря выбрасывает мочу в синус, задний конец которого открывается в клоаку с ее нижней стороны. Подобно териям, однопроходные уреотелические, с мочевиной в качестве основного продукта азотистого обмена. Мочевинный азот составляет 82-90 % всего азота, выделяемого с мочой. Конечным продуктом метаболизма пуриновых оснований является мочевая кислота, по-видимому, в дальнейшем окисляемая уриказой печени до аллантоина, как и у большинства плацентарных. Так или иначе, в моче и крови мочевой кислоты содержится совсем немного.
Сердечно-сосудистая система
Сердечно-сосудистая система не демонстрирует существенных отличий от таковой у териевых млекопитающих. Подобно последним, имеется 2 круга кровообращения, а дуга аорты происходит из левого ответвления IV-й жаберной дуги. Присутствуют остатки абдоминальной вены, свойственной амфибиям и рептилиям. Сердце четырехкамерное, однако сохраняет некоторые примитивные признаки. Клапан левого желудочка трехстворчатый, но его движениями управляют не сухожильные хорды, как у териев, а удлиненные сосочковые мышцы стенок желудочка, вложенные в перепончатые створки клапана. Правый предсердно-желудочковый клапан также управляется посредством сосочковых мышц. 2 коронарные артерии проходят по наружной поверхности сердца вполне типичным для териев образом. Отток венозной крови осуществляется за счет большой сердечной вены, которая, как у териев и птиц, открывается непосредственно в правое предсердие. Однако у однопроходных венозная кровь помимо этого переносится в правое предсердие коронарной веной, которая тоже открывается прямо в предсердие. Коронарные вены не встречены ни у других современных млекопитающих, ни у птиц. Водитель сердечного ритма развит относительно слабо. Волокна синусового и атриовентрикулярного узлов кажутся отдельными объектами, поскольку окрашены в более бледный цвет и имеют более округлые ядра, чем ядра сократительных волокон. Оба узла богато иннервированы аксонами, спирально обвитыми вокруг их волокон.
Уровень гемоглобина в крови достаточно высокий. Подобно териям (за исключением представителей семейства верблюдовых (Camelidae)), эритроциты имеют двояковогнутую форму и лишены ядра. Однако их необычной особенностью является крайне низкий уровень содержания аденозинтрифосфата (АТФ) — основного источника для утилизации глюкозы в эритроцитах взрослых териев. Тем не мене, благодаря гликолитическим ферментам эритроциты однопроходных способны метаболизировать глюкозу в лактат со скоростью, сопоставимой с таковой у других млекопитающих. Тромбоциты сходны с этими клетками у прочих представителей класса. В то же время, лейкоциты примечательны наличием телец Деле в сегментоядерных нейтрофилах. Аналогичным образом, несегментированный характер ядер эозинофилов и базофилов необычен по сравнению с большинством млекопитающих. У утконоса имеются базофилы и эозинофилы, тогда как у ехидн они практически отсутствуют.
Дыхательная система
Подобно териям и в противоположность рептилиям, в легких хорошо развита разветвленная система бронхов и бронхиол. Легкие большие и несимметричны: правое состоит из 2 долей, левое — из 1. У основания грудной клетки расположена хороша развитая диафрагма. Она очень широкая, мускулистая на периферии и сухожильная в центре непосредственно под сердцем.
Эндокринная система
Гипофиз представляет собой округлое образование, расположенное на конце воронки в основании головного мозга. Как и у всех позвоночных, он состоит из 2 различных по происхождению и структуре долей: вздутого задне-нижнего продолжения воронки — нейрогипофиза, и удлиненного аденогипофиза. Подобно другим позвоночным, нейрогипофиз в основном сложен немиелинизированными волокнами, в окончаниях которых запасаются нонапептидные гормоны, образовавшиеся в гипоталамусе. Из этих окончаний гормоны выделяются в кровоток. Как и в нейрогипофизе плацентарных, они представлены окситоцином и вазопрессином. Окситоцин являет особый интерес, поскольку очень важен для млечного выкапмливания детенышей. В состав аденогипофиза входят pars distalis, pars tuberalis и pars intermedia, последняя из которых накладывается на переднюю поверхность нейрогипофиза и отделена от остального аденогипофиза гипофизарной щелью. По своему строению гипофиз очень сходен с таковым у рептилий и обладает значительным подобием у утконоса и ехидн.
Надпочечники относительно маленькие (ехидны) или достаточно крупные, сравнимые с таковыми у плацентарных (утконос). Они расположены вплотную к передне-внутренним поверхностям почек вблизи выхода мочеточников. Наружная поверхность надпочечников выпуклая, тогда как внутренняя уплощенная. Внутреннее их строение в значительной мере сходно с рептильным. В отличии от териев, в надпочечниках которых корковая ткань со всех сторон окружает хромаффинную, у однопроходных корковая ткань сосредоточена в передней части надпочечников, тогда как хромаффинная — в задней. Четкая граница между обоими типами ткани отсутствует. В состав коры входит 3 распознаваемых видов клеток, но они не разделяются на концентрические зоны, как это наблюдается в надпочечниках териев. По сути, эта часть железы именуется корковой только из-за того, что здесь синтезируются стероидные гормоны (кортикоиды), как в истинной коре надпочечников териев. Можно предположить, что ферментативные системы для биосинтеза кортикостероидов сходны с таковыми у териев. У утконоса уровень секреции кортикостероидов и их содержание в крови относительно выше, чем у ехидн. По-видимому, это свидетельствует о большей значимости надпочечников в регуляции обмена веществ утконоса. В хромаффинной части надпочечников образуется адреналин.
Подобно надпочечникам, щитовидная железа утконоса секретирует гормоны интенсивнее, чем у ехидн, что связывают с его более высоким уровнем метаболизма и темпами роста. Поджелудочная железа дискретна, подобно хищным, а не диффузная, как у грызунов и зайцеобразных. Островки Лангерганса распределены по всей ее экзокринной ткани и содержат обычные альфа-, бета- и индифферентные клетки. В бета-клетках поджелудочной железы образуется инсулин.
Энергообмен
Температура тела относительно низкая и составляет в среднем 30-32 °С (для сравнения: у сумчатых — около 35 °С, у плацентарных — порядка 37 °С). Тем не менее, у активных животных температура остается постоянной в разнообразных условиях, значительно опускаясь лишь во время отдыха. Однако жару и утконос, и ехидны переносят плохо. Сравнительно низкая температура тела может являться примитивным признаком, унаследованным от базальных млекопитающих. Согласно другой точке зрения, она не была характерна для однопроходных изначально, а является результатом их приспособления к суровым жизненным условиям.
По уровню энергообмена однопроходные занимают промежуточное положение между пойкилотермными (рептилиями) и настоящими гомойотермными (териями и птицами), все же больше отклоняясь в сторону последних. Интенсивность основного обмена у утконоса сопоставим с таковым у большинства териевых млекопитающих. У ехидн, подобно другим мирмекофагам, его уровень невысок. В отличие от плацентарных и подобно сумчатым, бурая жировая ткань у однопроходных отсутствует. Таким образом, для них характерен лишь сократительный термогенез. Как и у рептилий, L-аскорбиновая кислота синтезируется только в почках.