Распространение
Современные представители группы распространены в Австралии, на Тасмании и Новой Гвинее. Ископаемые формы обитали также в Южной Америке и, вероятно, Антарктиде. Можно предположить, что однопроходные в указанных регионах некогда достигали значительного разнообразия, но их процветание закончилось с приходом сумчатых млекопитающих в самом конце мела или в начале кайнозоя.
Образ жизни
Для небольшой группы специализированных животных, однопроходные занимают удивительно широкий спектр обитаемых сред. Они населяют леса различного типа, поросшие кустарником степи, каменистые и песчаные участки, встречаются на равнинах и в горах, порой поднимаясь выше 4000 м над ур. моря. Ведут полуводный (утконос) или наземный (ехидны) образ жизни. Кормятся мелкими водными животными (утконос) или почвенными беспозвоночными (ехидны). Уровень пищевой конкуренции с другими животными незначителен. Активны в вечерних сумерках, ночью и на рассвете. Светлое время суток проводят в естественных укрытиях или собственных норах. Помимо полуденного зноя, однопроходные избегают и ночного холода, поэтому в прохладный сезон зачастую питаются днем. Ехидны при недостатке корма впадают в спячку, во время которой температура их тела лишь ненамного превышает температуру окружающей среды.
За исключением самок в период выведения потомства, однопроходные ведут одиночный образ жизни. Взрослые особи занимают индивидуальные участки, которые могут частично перекрываться с несколькими другими. Территориальность выражена слабо. За исключением сезона спаривания, животные демонстрируют очень мало интереса друг к другу. Естественных врагов у взрослых особей на сегодняшний день относительно немного. Являются очень робкими и скрытными существами, наблюдать за которыми нелегко. Содержание в неволе сопряжено с рядом трудностей, а добиться размножения в искусственных условиях крайне сложно.
Происхождение
Филогенетическое положение однопроходных проблематично. Их палеонтологическая летопись слабо способствует установлению родственных связей с другими группами. О примитивности однопроходных говорят признаки, унаследованные ими от зверообразных предков, общие с рептилиями и в то же время отсутствующие у териев: наличие септомаксилл, предлобных и заднелобных костей в черепе, присутствие межключицы, метакоракоидов и прокоракоидов в плечевом поясе, отсутствие эпифизов на большей части позвонков, наличие шейных ребер, некоторые примитивные особенности строения сердечно-сосудистой системы, откладывание яиц, наличие яйцевого зуба.
Вместе с тем, у современных однопроходных имеются многие черты, присутствующие также у териев и в массе своей являющиеся синапоморфиями млекопитающих: наличие (утконос) позднее исчезающих сложных щечных зубов, состав нижней челюсти из единственной зубной кости, причленяющейся непосредственно к чешуйчатой, 3 косточки в среднем ухе (молоточек, наковальня и стремя, развившиеся независимо от териев), 2 затылочных мыщелка, 7 шейных позвонков, эпифизы на костях конечностей, по 3 фаланги во II-м-V-м пальцах всех конечностей, волосяной покров, млечные железы (правда, имеющие примитивное строение), наличие брюшной сумки (впрочем, она тоже могла развиться независимо), способность к поддержанию постоянной температуры тела (хотя и на относительно низком уровне) и др.
Наконец, имеется ряд признаков, которые могут считаться новоприобритениями однопроходных: сильно вытянутый лицевой отдел черепа, редукция зубов с замещением их роговыми пластинками, слабо выраженная шея, приземистое туловище, короткие и мощные расставленные в стороны конечности, вывернутая наружу стопа, большое количество механо- и электрорецепторов в коже рыла, частично завитая ушная улитка, незначительное развитие ушной раковины, шпора на пятке стопы и связанная с ней железа. Кое-какие из них также характерны для некоторых териев, не связанных с однопроходными близким родством, но напоминающими их в экологическом отношении. Таким образом, однопроходные демонстрируют мозаику примитивных и специализированных черт, что вообще характерно для мезозойских групп млекопитающих.
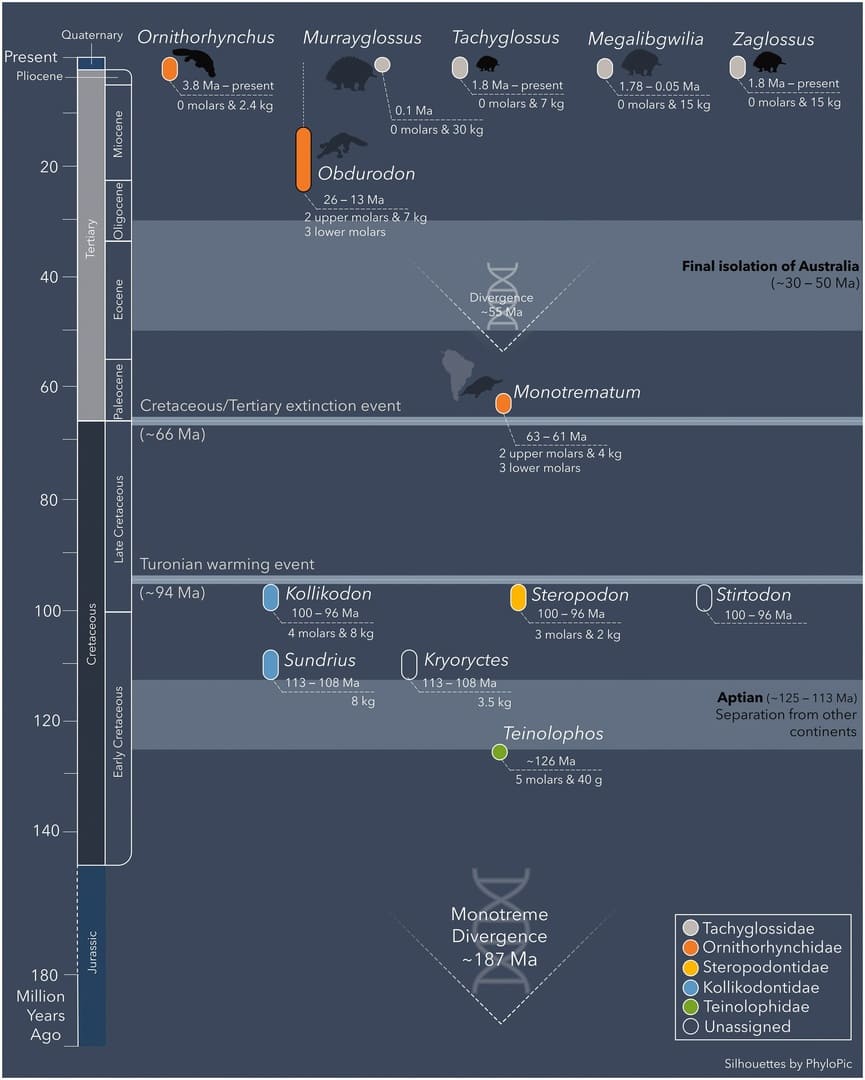
Стратиграфическое положение родов однопроходных (Monotremata).
Выделение эволюционной линии однопроходных произошло очень давно, вероятно, еще в первой половине мезозоя, так как меловые сумчатые и плацентарные уже утратили многие архаичные признаки, сохранившиеся у современных однопроходных. Эти плезиоморфные черты у однопроходных затемняются автапоморфными особенностями высокой специализации, развившимися в связи с их своеобразным образом жизни. Необычный для млекопитающих рацион, состоящий главным образом из мягкотелых беспозвоночных, привел к редукции зубной системы. У многих представителей группы наличествуют приспособления для рытья — широкие, длинные когти, верхний отросток малой берцовой кости, удлиненное назад твердое небо и др. У ехидн, поедающих муравьев и термитов, присутствует длинный червеобразный язык, очень тонкая нижняя челюсть без восходящей ветви, длинный тонкий клювовидный передний отдел морды с маленьким отверстием на конце. Значительно специализирован головной мозг ехидн — с довольно большими, снабженными извилинами полушариями; у утконосов головной мозг архаичный.
Когда в начале XIX в. впервые было обнаружено сходство однопроходных с рептилиями, среди ученых возникло буквально смятение; некоторые даже высказывали мнение, что они могут представлять собой недостающее звено между млекопитающими и рептилиями. На сегодняшний день филогенетическое положение однопроходных все еще продолжает оставаться неопределенным. Найденные окаменелости этих животных относительно редки, а до миоцена их известна всего лишь горстка. Примитивная биология и архаичное строение скелета позволяют предполагать дивергенцию предков однопроходных от самых ранних триконодонтов или даже млекопитающеобразных еще до конца триаса.
Согласно другой гипотезе, базирующейся на находке в нижнем мелу Австралии однопроходных (Teinolophos, Steropodon) с щечными зубами, строение которых представляется возможным вывести из примитивного трибосфенического типа, эта группа рассматривается в качестве близкородственной териям. Предполагают, что однопроходные (шире — австралосфениды (Australosphenida)) могли обособиться от общего с териями ствола на уровне дриолестид (Dryolestida) или эвпантотериев (Eupantotheria), но какие-либо связующие звенья между ними не обнаружены. Свидетельства наличия у однопроходных общих предков с более поздними териевыми или нетериевыми группами также отсутствуют. Согласно результатам молекулярно-генетических исследований, однопроходные отклонились от линии териев 217-161 млн. лет назад, еще до их разделения на сумчатых и плацентарных. Следуя еще одной точке зрения, однопроходные могли отделиться от дотрибосфенических млекопитающих независимо от австралосфенид. Наконец, существует экстремальное предположение о происхождении однопроходных от терапсид независимо от других млекопитающих.

Однопроходные позднемеловой фауны Лайтнинг-Ридж из Нового Южного Уэльса, Австралия; по часовой стрелке снизу слева: опалий (Opalios splendens), стиртодон (Stirtodon elizabethae), колликодон (Kollikodon ritchiei), стероподон (Steropodon galmani), парвопал (Parvopalus clytiei), дхарражарра (Dharragarra aurora). Автор — П. Схоутен
Однопроходные известны лишь с материков, некогда составлявших Восточную Гондвану. Немногие мезозойские находки предположительно указывают на то, что они возникли от изолированных в Австралийской области предков в ранне- или среднеюрское время. Гипотетическое базальное однопроходное могло иметь следующий облик. Это было мелкое насекомоядное существо, ведущее наземный образ жизни. Ноздри находились почти на самом конце конического рыльца и были окружены голым, влажным ринариумом. Глаза и ушные раковины были относительно небольшими. Достаточно длинные конечности занимали более парасагиттальное положение, чем у современных представителей группы. Лапы несли хорошо развитые когти и были приспособлены для рытья. Хвост достаточно длинный, круглый в сечении и сужающийся к концу. Тело покрывал хорошо развитый мех, однако вибриссы отсутствовали. В целом животное могло напоминать безиглого ежа или тенрека.
Впоследствии однопроходные через Антарктиду распространились в Южную Америку, но, вероятно, ввиду ухудшения климата, а также конкуренции и хищничества со стороны более прогрессивных сумчатых млекопитающих не смогли задержаться ни на одном из этих континентов длительное время. Тот факт, что однопроходные сохранились в Австралийской области, может быть объяснен отчасти их высокоспециализированным образом жизни, отчасти же — продолжительной изоляцией этого региона от прочих материков. Остатки древнейших австралийских сумчатых имеют лишь раннеэоценовый возраст, а ко времени возникновения серьезной конкуренции с этой группой местные однопроходные уже достаточно надежно закрепились в своих экологических нишах. Согласно молекулярным данным, эволюционные линии ехидн и утконосов разошлись в период от 48 до 19 млн. лет назад.
В качестве наиболее вероятной сестринской группы для однопроходных рассматриваются геносфериды (Henosferida) из среднеюрских отложений Западной Гондваны. У обеих групп отмечено продвинутое претрибосфеническое строение нижних коренных при вероятном отсутствии протокона на верхних зубах и плезиоморфное сохранение зазубных костей и "ложноуглового" отростка нижней челюсти. Также общими для этих групп признаками являются зубная формула с 3 коренными зубами и положение меккелевой борозды, которая проходит ниже нижнечелюстного отверстия. В ходе дальнейшей эволюции у однопроходных сформировалось "маммальное" среднее ухо с 3 слуховыми косточками — сходное с таковым у териевых млекопитающих, но появившееся независимо от них. На последнее, в частности, указывает негомологичность у териев и однопроходных таких тесно связанных со средним ухом структур, как депрессоры нижней челюсти.
Разнообразие ископаемых однопроходных (Monotremata). Автор — artbyjrc.
Систематика
Для однопроходных установлен следующий состав:
семейство †Тейнолофовые (Teinolophidae Flannery, Rich, Vickers-Rich, Ziegler, Veatch et Helgen, 2022);
? семейство †Колликодонтовые (Kollikodontidae Flannery, Archer, Rich et Jones, 1995);
семейство †Стероподонтовые (Steropodontidae Archer, Flannery, Ritchie et Jones, 1995);
надсемейство Орниторинхоиды (Ornithorhynchoidea Flannery, McCurry, Rich, Vickers-Rich, Smith et Helgen, 2024);
семейство †Опаловые (Opalionidae Flannery, McCurry, Rich, Vickers-Rich, Smith et Helgen, 2024);
семейство Утконосовые (Ornithorhynchidae Gray, 1825);
семейство Ехидновые (Tachyglossidae Gill, 1872).