в оглавление
назад
Потребление воды (продолжение)
Многие животные аридных зон отличаются сравнительной медлительностью и неповоротливостью. Представим на мгновение поведенческие стереотипы двугорбого верблюда (Camelus bactrianus), хомяка (Cricetus cricetus) или сурка-байбака (Marmota bobak). Локомоции этих животных отличаются особой рациональностью. Лишние движения требуют дополнительной энергии и воды. Любопытно, что для обслуживания мышечных сокращений у пустынных животных используется метаболическая вода, которая к тому же не расходуется на охлаждение за счет испарения. Потерю воды на терморегуляцию также поисками прохладного места. Сюда относится и жизнь в норах. Эта поведенческая адаптация дает возможность использовать почвенную влагу и экономить воду за счет сокращения испарения с поверхности тела.
Итак, поддержание водного баланса тесно связано с питанием и терморегуляцией. Важно учитывать, что сохранение воды некоторым образом невыгодно для животного, поскольку может мешать нормальной терморегуляции или снижать поступление энергии. Различные механизмы сохранения по-разному важны для разных видов, что зависит от нормальных экологических условий их существования. На первый взгляд система регуляции водного баланса служит наглядным примером гомеостаза: животное обнаруживает отклонения от нормальных количеств (объема) и концентрации (осмоляльности) внеклеточной воды и принимает меры для исправления положения, поглощая воду или снижая ее потери посредством тех или иных механизмов. Однако ситуация не так проста из-за взаимодействия с другими системами. Например, животное может потреблять воду, когда оно не обезвожено.
Животные пьют воду непосредственно из водоемов или получают ее вместе с пищей, содержащей достаточное количество влаги, или, наконец, в областях с холодной зимой, — поедая снег. Последний отличается от питьевой воды тем, что почти не содержит минеральных примесей, поэтому снежная вода «безвкусна» и при длительном потреблении (например, в тундре, где продолжительность снежного периода достигает 240-260 и более дней в году) вызывает у растительноядных млекопитающих острое минеральное голодание. У одомашненного северного оленя (Rangifer tarandus), питающегося зимой кормами, бедными зольными элементами, и около 8 месяцев получающего воду из поедаемого снега, кости к весне становятся крайне хрупкими: недостаток минерального питания организм восполняет за счет мобилизации своих внутренних ресурсов, до предела расходуя то, что отложено в скелете. При ловле оленей арканом и других операциях упавшие животные нередко получают переломы ребер, костей ног и т. п., что осенью бывает крайне редко. Длительное употребление снежной воды не проходит бесследно и для человека, вызывая ухудшение состояния зубов и т. п. Замечено также, что во всех случаях, когда зимой есть доступ к воде ключей, наледей или незамерзающих порожистых участков рек, и растительноядные и хищные млекопитающие отдают ей предпочтение по сравнению со снегом.
С другой стороны, сильная минерализация воды, избыток растворенных в ней солей делает воду океанов, морей и многих внутренних водоемов засушливых областей совершенно непригодной для ряда млекопитающих, происходящих из областей с достаточным или избыточным увлажнением, где они всегда пользовались «хорошей питьевой водой». Например, человек может пить соленую океанскую воду в ограниченном количестве в продолжение всего 4-5 дней; в дальнейшем организм настолько перегружается солями, что почки не справляются с их выведением, вследствие чего развивается острое заболевание почек — нефрит. В то же время джейраны, обитающие на восточном побережье Каспия, ходят на водопой даже к Кара-Богаз-Голу, крайне насыщенному солями. Двугорбый верблюд (Camelus bactrianus), происходящий из аридных областей Центральной Азии, охотно пьет сильно солоноватую и горько-соленую воду пустынных озер, от которой отказываются даже долго не имевшие воды лошади и коровы. Породы скота, подвергшиеся длительному отбору, например, каракульские овцы из Каракумов, также успешно потребляют сильно минерализованную воду из колодцев сухих степей и пустынь. Однако в вопросах об оценках воды по ее пригодности для водопоя и нормах ее потребления различными животными еще много неясностей.
Исключительно большую роль открытые водопои играют в жизни рукокрылым — группы, особенно богатой видами и особями в субтропических районах и еще более — в тропиках Азии, Африки и Америки. Покидая вечером свои убежища, летучие мыши прежде всего направляются к водоемам (ключам, широким арыкам, прудам, озерам) и пьют налету, после чего начинают охоту за насекомыми. Охлаждение организма при напряженной мускульной работе во время полета осуществляется через испарение воды с поверхности легких, рта и, возможно, других участков тела летучих мышей (установлено, что мелкие птицы во время сезонных миграций теряют очень много воды при беспосадочных перелетах над морем и т. п.). Поэтому, питаясь насекомыми, подобно ушастым ежам (Hemiechinus), рукокрылые в отличие от последних не могут обходиться без водопоя. Все колонии и поселения летучих мышей, находящиеся в пустынях, степях и горах аридных континентальных районов, расположены поблизости от водоемов, пригодных для подчерпывания воды налету. Это обстоятельство очень четко сказывается на географическом распределении колоний летучих мышей в засушливых областях. Последнее без учета водопоев не может быть правильно понято. Примечательно, что регулярно пьют и охотящиеся на лету насекомоядные птицы (стрижи, ласточки, щурки, козодои), тогда как виды с другой биологией, но живущие в тех же районах, могут обходиться без водопоя.
Наоборот, мало нуждаются или совсем не нуждаются в водопоях многие сумчатые, ксенартры, насекомоядные, грызуны, хищники и копытные, живущие в пустынях и сухих степях всех континентов. В процессе эволюции у них выработались уникальные механизмы рационального использования влаги. Так, вомбатам (Vombatidae) достаточно всего около 22 мл воды на 1 кг массы тела в сутки. Многие мелкие зверьки, например ушастые ежи (Hemiechinus), суслики (Spermophilus), песчанки (Gerbillinae), тушканчики (Dipodidae), всю необходимую влагу получают из пищи. Весной ими поедается сочная зелень и насекомые, а когда наступит период летней жары и земля днем начнет раскаляться до 60-80 °С, они выкапывают луковицы, клубни, надкусывают мощные стержневые корни многолетних растений и слизывают выступающий сок. В сухой и морозный период зимы в песках Каракумов грызуны получают необходимую влагу, обгладывая сочную кору кустов и деревьев — кандымов, саксаула, песчаной акации и др. Эти растения имеют мощную корневую систему, достигающую грунтовой воды. Кроме того, грызуны поедают мелкую растительную труху, покрытую инеем, который выпадает по ночам. Однако не следует думать, что эти зверьки вовсе не умеют пить — когда весной на такырах скопляется снеговая вода, ее пьют суслики, песчанки, ушастые ежи и зайцы-толаи, обитающие поблизости, но это удел крайне малого числа особей.
Мелкие грызуны, содержавшиеся в клетках и получавшие в качестве корма овес, овсянку и свеклу, потребляли каждый за сутки в среднем следующее количество воды: полевая мышь (Apodemus agrarius) — 1,7 куб. см, домовая мышь (Mus musculus) — 1,5 куб. см, мышь-малютка (Micromys minutus) — 1,3 куб. см, азиатская лесная мышь (Apodemus speciosus) — 1,25 куб. см, рыжая лесная полевка (Myodes glareolus) — 1,0 куб. см, европейская лесная мышь (Apodemus sylvaticus) — 0,7 куб. см. Все перечисленные зверьки — обитатели районов с мезофитной растительностью и достаточно обильными осадками. Резко отличается от них степная пеструшка (Lagurus lagurus). Из 12 ее особей, участвовавших в опыте, воду пили только 4; их суточная норма потребления равнялась всего 0,1 куб. см, наличия сочного корма — свеклы было вполне достаточно, чтобы удовлетворить потребность пеструшек в воде. Это вполне объяснимо, поскольку в природе пеструшкам нередко в течение месяцев приходится питаться довольно сухими листьями степных злаков, полыней и т. п., совсем не имея питьевой воды.
Грызуны аридных зон обладают уникальной способностью удовлетворять свои потребности в воде за счет метаболической влаги семян пустынных растений и паров, содержащихся в атмосферном воздухе. Египетская песчанка (Gerbillus pyramidum), кенгуровые (Dipodomys) и мешотчатые (Perognathus) прыгуны способны питаться семенами, которые содержат менее 10 % воды. Щетинистых прыгунов Сан-Диего (Chaetodipus fallax) в течение 3 лет содержали на сухих зернах без питьевой воды. Метаболической влаги семян было достаточно для того, чтобы зверьки в полной мере обеспечивали свой обмен веществ водой. За это время самки принесли многочисленное потомство, которое успешно выкармливали собственным молоком.
Эксперименты показали, что кенгурового прыгуна Мерриама (Dipodomys merriami), обитающего в пустынях на юго-западе США и в Мексике, невозможно обезводить за счет лишения питьевой воды и кормления исключительно сухими злаковыми зернами. На протяжении длительного времени содержание воды в теле зверька не изменялось. Однако толерантность животных к дефициту воды не беспредельна. Даже у кенгурового прыгуна начинается дегидратация тела при относительной влажности воздуха ниже 8 %. При такой низкой влажности резко возрастает испарение воды с поверхности тела. Поскольку в реальной жизни относительная влажность атмосферного воздуха даже в пустынных районах выше этой критической отметки, угрозы обезвоживания организма обитателей пустынь не существует. Когда кенгурового прыгуна содержали на перловой крупе в климатической камере, в которую подается воздух с нулевой влажностью, он начинал терять вес за счет дегидратации. Но уже при содержании 2-3 мг воды в одном литре воздуха живая масса грызуна стабилизировалась. А при более высокой влажности воздуха живая масса зверька быстро нарастала за счет восстановления потерянной влаги.
Также можно вызвать обезвоживание организма кенгурового прыгуна путем увеличения метаболических затрат воды. В экспериментальных условиях такой результат получают при назначении животному высокобелковой диеты, например, за счет включения в рацион соевых бобов, жмыха, гороха. Образующаяся в процессе катаболизма азотсодержащих веществ мочевина повышает осмотическое давление и водородный баланс внутренней среды, т. е. изменяет одну из наиболее жестких констант гомеостаза. Для понижения осмотического давления и выведения мочевины из организма требуется вода. Поэтому белковая нагрузка приводит к дегидратации животного организма. Таким образом, становится понятным биологический смысл изменения поведения ряда животных в условиях дефицита питьевой воды. В целях сохранения жизни им остается одно — вообще отказаться от пищи. Поэтому в условиях высоких температур атмосферного воздуха и его низкой относительной влажности поведение животных меняется. Они ограничивают локомоции, сокращают потребление корма и становятся разборчивыми к его составу. В предельно экстремальных условиях некоторые пустынные обитатели прибегают к последнему маневру — они впадают в спячку и таким образом экономят влагу и сохраняют жизнеспособность.

Водный баланс кенгурового прыгуна (Dipodomys sp.). Этот зверек обитает в местах, где температура воздуха может подниматься до 40 °C в тени. Поэтому весь день он проводит в норе, где сохраняется температура около 30 °C и выше влажность. Кенгуровый прыгун активен только в сумерки и рано утром, когда выходит на поиски пищи. Он никогда не пьет и не потеет, а в его почках в 5 раз больше фильтрующих капсул, чем у человека. Благодаря высококонцентрированной моче организм теряет мало воды.
Хищные, например обычные в пустынях Азии перевязка (Vormela peregusna), корсак (Vulpes corsac), барханная кошка (Felis margarita), а в Африке — лисица фенек (Vulpes zerda), черноногая кошка (Felis nigripes) и др., получают достаточное количество влаги, поедая своих жертв — грызунов, насекомых, и т. п. При случае хищники тоже охотно пользуются вегетационной, или «растительной водой», поедая, например, плоды дикого арбуза, широко распространенного в пустынях Африки и Передней Азии, а на бахчах степей и пустынь — культивируемые арбузы и дыни. Волк (Canis lupus) и лисица (Vulpes vulpes) являются частыми посетителями бахчей и местами серьезные враги бахчеводства. Установлено, что в сухих подгорных районах Туркмении шакал (Canis aureus) сильнее объедает грозди винограда на плантациях, когда в арыках нет воды и наоборот. Однако более высокая численность выводковых нор волка, шакала, лисицы, полосатой гиены и ряда других видов поблизости от водопоев и явная спорадичность распределения в безводных участках пустынь указывают, что питьевая вода в жизни крупных хищников играет большую роль, чем в жизни мелких куниц, ежей и т. п.
Как было сказано, крупные фитофаги засушливых областей также приспособлены к дефициту влаги. Медвежий павиан (Papio ursinus), обитающий в пустыне Намиб на юге Африки, по-видимому, может обходиться без воды до 26 дней. Жираф (Giraffa camelopardalis) в пустыне не живет, но питье воды для него — поистине тяжкое испытание. Чтобы дотянуться до лужи, ему приходится широко расставлять ноги и осторожно опускать шею и голову вниз. Поскольку в это время жираф может стать легкой добычей львов и других хищников, пить воду он старается как можно реже — иногда раз в несколько недель. Всю необходимую организму влагу он получает с древесной листвой.

Жираф (Giraffa camelopardalis) на водопое.
Образцом неприхотливости в засушливых условиях пустыни служат курдючные и жирнохвостые овцы, зебувидный скот, и особенно верблюды. В условиях недостатка или полного отсутствия воды ее источником у них служит внутренний жир. Масса жира, накопленного в курдюках овец и горбах верблюдов, составляет до 20 % от их живой массы. При окислении этого жира выделяется достаточное количество воды, чтобы не только поддержать основной обмен, но и выполнять все необходимые движения, связанные с пищевым, половым поведением, защитными действиями. Расчеты показывают, что 50-килограммовая курдючная овца при себе имеет запас воды около 10 л, а 500-килограммовый верблюд — около 100 л. У бореальных животных имеет место значительный объем желудочно-кишечного тракта и большая относительная длина кишечника, у них откладывается больше внутреннего жира в сальниках и околопочечной капсуле.
Впрочем, значение метаболической воды, образующейся в организме верблюдов при использовании жировых отложений, значительно меньше того, какое ему приписывали ранее. Летом этим выносливым копытным при каждом водопое требуются десятки литров воды — например, один самец массой 352 кг употребил 104 л за один прием. Однако весной верблюды могут проводить на пастбищах до 2-2,5 месяцев, не приближаясь к водным источникам и получая всю необходимую влагу вместе с поедаемыми растениями. При температуре воздуха 40-50 °С эти копытные, потребляя только сухие корма, остаюся активным и обходится без питьевой воды в течение 2-3 недель. Вопреки прежним представлениям, выпитая ими вода не резервируется в желудке, а чрезвычайно быстро всасывается. Благодаря особенностям их кровяной плазмы даже острый дефицит воды в организме не вызывает сгущения крови, т. е. не нарушает нормальной циркуляции последней. Состав молока лактирующей верблюдицы также остается постоянным и не зависит от количества выпитой воды.
При критически высоких температурах воздуха и отсутствии питьевой воды верблюды повышают теплоотдачу тела за счет иррадиации и излучения. При этом они не тратят воду на терморегуляцию через испарение. Параллельно с этим происходит снижение теплопродукции как следствие снижения уровня обмена веществ. Установлено, что верблюды способны на четверть понизить основной обмен. Однако остается неясным, каков механизм самоконтроля у верблюда. Похожие возможности манипулирования напряжением обмена веществ демонстрируют йоги. Но следует подчеркнуть, что 25 %-ное понижение обмена у человека не совместимо с жизнью.
Не менее примечательна способность верблюдов к терморегуляции. Выделение пота у них начинается лишь при 41 °C, когда дальнейшее повышение температуры тела непосредственно угрожает их жизни (человек потеет уже при температуре тела 37 °C). Наконец, во время сна и отдыха температура тела верблюдов существенно снижается (иногда на 10 °C), что также сокращает расход воды. Вследствие «плавающего» характера интенсивности обмена веществ верблюды формально не могут быть называны гомойотермными, т. е. животными с постоянной температурой тела.
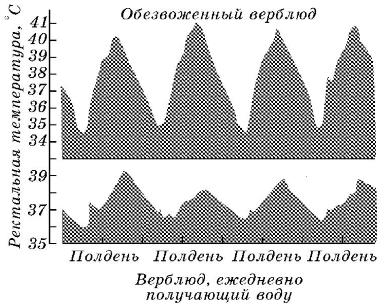
Суточные колебания температуры тела двугорбого верблюда (Camelus bactrianus).
Интересно и то, что верблюды не прибегают к поведенческим адаптациям. Они не тратят энергию и воду на поиски тени или укрытия, не смещают свою активность на ночное время. Верблюды экономят на другом. Их локомоции отличаются медлительностью и плавностью, что обеспечивает меньшие затраты энергии и воды. При первой возможности животные ложатся на живот или на бок, чем дополнительно сокращают затраты воды на обмен веществ и мышечные сокращения. Режим дыхания у верблюдов почти не изменяется. Количество образующейся и выделяемой мочи у них существенно меньше, чем у других видов жвачных животных. К тому же эти копытные обладают уникальной способностью концентрировать мочу. При дефекации верблюды выделяют горячий кал с очень низким содержанием воды. Последнее обстоятельство позволяет животным решить сразу три задачи: завершить процесс пищеварения, избавиться от избыточного тепла и сохранить воду.
Другую тактику в сходных условиях демонстрируют аборигенные породы овец. На фоне верблюдов они существенно отличаются тем, что прежде всего вовлекают в процесс адаптации этологический ресурс. Овцы пустынь активные действия переносят на то время суток, когда температура воздуха минимальна. Так, пищевая активность у них наиболее высока ранним утром. Еще до наступления полуденной жары животные уходят в тень под навес. Если тени в ближайшем окружении нет, курдючные овцы стараются отыскать подходящий участок (свободный от камней, в идеале сырой), на который ложатся животом. Такое поведение овец имеет большой смысл. В области живота у пустынных овец шерстный покров развит слабо или вовсе отсутствует. Поэтому контакт живота с относительно прохладной землей способствует избавлению от лишней тепловой энергии. Овцы, подобно верблюдам, экономят воду на мочеиспускании и дефекации. У курдючных, каракульских и прочих аборигенных овец пустыни моча очень концентрированная, а кал просто напоминает камни — настолько в нем мало воды. Особенностью овец является и то, что потребление холодной воды у них вызывает снижение ректальной температуры на 1-2 °С.
Часто встречается утверждение, что копытные пустынь и степей, обладающие способностью к быстрому бегу, регулярно пользуются водопоями, иногда расположенными даже на расстоянии десятков километров от их пастбищ. В действительности для большинства видов особой необходимости в этом нет, так как они находят зеленые растения, содержащие достаточное количество воды. Весной кормовые травы песчаных пустынь Средней Азии (в основном эфемеры и эфемероиды) содержат 75-80 % влаги, а некоторые до 90 % и даже выше, т. е. влажность их измеряется отношениями 1 : 3, 1 : 4 и даже 1 : 9. При выпадении дождей или росы это отношение у смоченного травостоя повышается до 1 : 14 и даже 1 : 19. Многие массовые кормовые растения содержат очень много воды и без наружного их смачивания. В пустынях Средней и Центральной Азии таковы солянки, в пустынях Африки — молочаи и уже упоминавшийся дикий арбуз, в пустынях Америки — кактусы, которые содержат до 91-94 % и более воды. Каракульская овца в пустыне при летней пастьбе на участках с высохшим травостоем выпивает в сутки 10-12 л (редко до 15), а весной, питаясь сочной свежей травой — только 1-2,5 л воды.
Лишь по мере высыхания весеннего зеленого ковра в пустынях копытные начинают кочевать в поисках менее выгоревших пастбищ и приближаются к водопоям. Кормящие самки джейрана (Gazella subgutturosa) с ягнятами собираются близ рек, озер на равнинах или у ключей в предгорьях. Лактирующие животные и подрастающие молодые более нуждаются в воде, чем взрослые самцы. Последние даже в жаркие месяцы довольствуются растительной влагой: ощипывают сочные листья каперса, верхушки верблюжьей колючки и других долго не высыхающих растений с мощными корнями. Местами жарким летом, как, например, по течению р. Сарысу в 1948 году, встречается масса самок джейрана, откочевавших с молодыми ближе к воде, тогда как самцы остаются в безводных местах пустыни Бетпак-Дала на многие десятки километров южнее. Антилопы и зебры Африки в случае пересыхания водоемов также совершают кочевки и настоящие миграции или выкапывают глубокие ямы в песке, добираясь до водоносного слоя. Ямы-колодцы на обсохших озерах Казахстана роют волки.
Несомненно, физиологическая роль «растительной» и обычной питьевой воды не равноценна. Вода сочных растительных кормов, связанная с коллоидами, лучше усваивается организмом, положительно влияет на интенсивность роста тканей, в том числе и на формирование половых клеток. У свиней скармливание сочных корнеплодов сильно повышает плодовитость (увеличение числа поросят в каждом помете). Необходимо учитывать, что постоянное потребление млекопитающими вегетационной воды — явление, очень широко распространенное. Таким образом, предлагаемые некоторыми авторами термины «снегопой» и «травопой» заслуживают признания наряду с привычным «водопой», так как с точки зрения экологии млекопитающих каждому из них соответствует особое содержание.
Поедая очень сочный и водянистый корм на весенних пастбищах в пустыне или на летних в лесной зоне, животные получают не только вполне достаточное, а порой даже чрезмерное количество воды. Избыточное количество влаги получают также олени (Cervus), лоси (Alces alces), косули (Capreolus) и серны (Rupicapra rupicapra), пасущиеся на сочных горных лугах (например, Кавказа, Алтая, Саян), где выпадает много летних осадков. Замечено, что у животных при такой пище расстраивается пищеварение и они жадно поедают мелкоструктурные глины и суглинки, отыскивая их в обнажениях склонов, в конечных моренах и т. п. Такие места нередко называют солонцами, хотя в поглощаемом млекопитающими материале почти полностью отсутствуют важные биогенные соли. У животных, поедающих мелкоструктурную глину, пищевая кашица становится более густой, и дефекация приходит в норму (поедание глины домашними копытными также чаще отмечают при выпасе на свежей траве и в дождливую погоду).
Животноводы знают, что излишнее потребление воды домашними животными приводит к значительному скоплению жидкости в тканях, вызывает их вялость и отечность, у молодых животных ухудшает пищеварение. Рабочие лошади при чрезмерно водянистом корме слабеют и потеют, овцы подвергаются тяжелым общим заболеваниям и т. д. С этими фактами следует сопоставить установленное наблюдениями улучшение биологической ценности пастбищ диких животных после подсыхания части растений и созревания их семян. Избыток воды устраняется, а семена, содержащие много ценных жиров, белков, углеводов, служат для накопления энергетических резервов — подкожного и полостного жира, гликогена и т. п., необходимых как при спячке, так и для активной жизни в сезоны с резко ухудшенными кормовыми условиями.
Итак, при внимательном изучении проблемы обнаруживается, что животные аридной зоны все-таки обладают способностью к непосредственному резервированию воды в собственном организме. Роль депо воды у животных, адаптированных к постоянному дефициту влаги, выполняет скелетная мускулатура, соединительная ткань и паренхиматозные органы. Сезонные колебания концентрации воды в мясе туркменских овец лежат в пределах от 75 % до 82 %. Максимального уровня этот показатель достигает в мае после выпаса животных на сочных кормах. Осенью, т. е. после засушливого летнего периода с дефицитом воды, концентрация влаги в скелетной мускулатуре овец минимальна. Для сравнения отметим, что у животных умеренного климата с высоким уровнем атмосферных осадков содержание воды в скелетной мускулатуре не подвержено сезонным колебаниям и остается постоянным (75-76 %) круглогодично. У архаромериносовых овец этот показатель существенно ниже, чем у овец засушливой зоны, — 55-60 %.
вперед
в оглавление