в оглавление
назад
Потребление воды
Вода совершенно необходима для нормального функционирования любого живого организма. Она является одним из важнейших строительных материалов при образовании и росте клеток, также она растворяет различные вещества и играет важную роль в процессах терморегуляции.
В среднем животный организм содержит около 70 % воды. Порядка 70 % ее находится внутри клеток в составе клеточной протоплазмы. Эту внутриклеточную воду именуют структурируемой, она обладает высокой биологической активностью и обеспечивает устойчивость организма к воздействию агрессивных факторов окружающей среды. Оставшиеся 30 % воды организма приходится на внеклеточную жидкость, из которой межклеточная жидкость составляет 20 %, вода плазмы крови — 8%, вода лимфы — 2%. В крови объемная доля воды составляет примерно 90 %, в веществе мозга — 85 %, в тканях почек — 80 %, в мышцах — 75 %, в печени — 70 %, в костях — 30 %, в жировых тканях — 25 %. С возрастом относительное количество воды в тканях уменьшается.
Недостаток воды большинство млекопитающих переносит гораздо труднее, чем недостаток пищи. Эмоция жажды создает условия для первоочередного удовлетворения потребности в воде. Приоритетность питьевого поведения при жажде биологически оправдано. Во-первых, у подавляющего большинства животных не существует органов депонирования воды, т. е. вода не запасается животными впрок. Во-вторых, пагубные последствия дегидратации организма столь велики, что на ее фоне удовлетворение других потребностей выглядит полной бессмыслицей. Лабораторные мыши, лишенные одной воды, умирают в 10 раз скорее, чем лишенные всякой пищи, но получающие воду. Это в полной мере относится и к человеку: без пищи летальный исход наступает, при сравнительно благоприятной температуре воздуха, через 25-30 дней, а без воды человек погибает уже спустя 10 дней. Потребность взрослого человека в воде равна 35 г в сутки на 1 кг веса тела; у ребенка она выше в 3-4 раза.
Если воды, добытой во внешней среде, не хватает, используется влага, образующаяся при обмене веществ, — т. н. метаболическая вода. Расщепление различных питательных веществ сопровождается образованием разного количества воды. При использовании за сутки 1 кг жира образуется 1,1 л воды, 1 кг крахмала — 0,5 л, 1 кг протеинов — 0,4 л. Ткани в очень малой степени способны утрачивать входящую в их состав жидкость: при длительном голодании, потеряв весь жир и половину белков, тело утрачивает только 10-15 % воды. Потери более 20 % воды в большинстве случаев сопровождаются необратимыми изменениями в организме и приводят к смерти. При этом терпимость к дегидратации организма, как и ее интенсивность, у разных видов и экологических типов животных неодинакова. Например, организм верблюдов (Camelus) выдерживает потерю до 40 % влаги, а потери воды через кожу у мешотчатых прыгунов (Heteromyidae) в 2-3 раза ниже, чем у находящихся с ними в одинаковых условиях содержания лабораторных крыс и мышей.

Прайд львов (Panthera leo) и саванный слон (Loxodonta africana) на водопое.
Проблемы с обеспечением организма водой прежде всего касаются животных, обитающих в жарком засушливом климате. В зоне умеренного климата отсутствие питьевой воды не имеет таких тяжелых последствий для животных, как в низких широтах. Сочная растительность, в достатке имеющаяся в районах с умеренным климатом, обеспечивает животных влагой и в отсутствие питьевой воды. Однако появление жажды у животных не обязательно вызвано высокими температурами и дефицитом питьевой воды. Высокая осмоляльность внутренней среды может развиться в случае избыточного поступления в организм животного солей (прежде всего хлорида натрия), выделения из организма большого объема воды, как это бывает у высокопродуктивных лактирующих коров с молоком или в процессе родов с околоплодными водами.
Частота приема воды зависит от многих факторов, среди которых можно назвать состав рациона, режим питания, температуру окружающей среды, физическую нагрузку, лактацию у самок. На лабораторных крысах показано, что при кормлении животных сухими кормами частота подходов к поилке составляет 7-10 за сутки. При кормлении крыс влажными мешанками частота подходов к поилке сокращается в несколько раз. Собаки при одноразовом кормлении влажным кормом и при средней физической нагрузке зимой пьют 3-4 раза, а летом — до 10 раз.
Крупный рогатый скот при содержании в летнее время на пастбище пьет воду 3-4 раза. В условиях зимнего стойлового содержания кратность потребления воды у лактирующих коров возрастает в 2-3 раза. Однако количество потребляемой за один подход воды меньше, чем при содержании коров в летнее время на пастбище. Общее количество выпиваемой за сутки воды у лактирующих коров зависит главным образом от суточного удоя и влажности корма. Специалисты отмечают наличие межпородных различий в потреблении воды. Браманский, зебувидный скот, буйволы пьют реже и при этом выпивают меньшее количество воды по сравнению с представителями европейских пород крупного рогатого скота. Межпородные различия характерны и для мелкого рогатого скота. Овцы и козы аридной зоны (например, курдючные овцы) более экономны в потреблении воды по сравнению с романовскими овцами.

Пьющий воду такин (Budorcas taxicolor).
Организм усваивает воду следующим образом. Попав в рот, она затем проходит через пищевод, желудок и достигает кишечника. Если концентрация солей в жидкости кишечника ниже, чем в крови, вода посредством осмоса проникает в кровь. Но если эта концентрация выше (например, после приема соленой пищи или воды), вода может мигрировать в обратном направлении и в сосудах наступает временное обезвоживание. Выход воды из кровеносных сосудов приводит к изменению концентрации солей во внеклеточном пространстве, что вызывает некоторое перераспределение воды между ним и клетками; в результате клетки частично обезвоживаются и сморщиваются. Эти изменения обнаруживаются специальными клетками в гипоталамической области мозга, называемыми осморецепторами. Их стимуляция оказывает два главных действия: усиленный поиск воды для питья и активацию различных механизмов ее сохранения.
Осморецепторы контролируют выделение антидиуретического гормона вазопрессина из гипофиза, расположенного непосредственно под гипоталамусом. Присутствие вазопрессина в кровотоке ведет к понижению количества и повышению концентрации мочи, выделяемой почками. Повреждение гипофиза или связанных с ним областей гипоталамуса вызывает несахарный диабет, к симптомам которого относится чрезмерное мочеотделение и вследствие этого усиленная жажда. Таким образом, в основе адаптационного функционирования мочевыделительной системы лежит нейрогуморальная регуляция с ярко выраженной гормональной составляющей.
Животные обезвоживаются не только вследствие дегидратации клеток, но и за счет уменьшения объема внеклеточной жидкости. Геморрагия и другие формы кровопотерь не меняют осмотического баланса, но утраченная жидкость должна быть возмещена. Для обнаружения такой потери у животных имеются разные механизмы. В ответ на снижение объема крови, протекающей через почки, вырабатывается гормон ренин. Он поступает в кровь и там стимулирует синтез другого гормона — ангиотензина, который оказывает два основных воздействия: во-первых, поддерживает нормальное кровяное давление — а следовательно, регулирует кровообращение, — во-вторых, служит мощным возбудителем жажды (животные, получившие ничтожные количества ангиотензина, прерывают любую свою деятельность и начинают искать воду).

Западные серые кенгуру (Macropus fuliginosus) пьют из временной лужи.
Процессы пищеварения пустынных животных подчинены задаче экономии воды. Так, у овец специфика проявляется уже на стадии ротового пищеварения. У этих животных прием сухого корма не сопровождается усилением секреции слюнных желез смешанного типа, как это имеет место у всех других животных и человека. В формировании пищевого кома принимает участие только серозная слюна, выделяемая околоушными железами. Подчелюстная и подъязычная железы при приеме корма не активизируются. Интересно, что попадающий в ротовую полость песок также не вызывает усиления саливации, т. е. у пустынных овец нет того, что называют «отмывной слюной». У ягнят экспериментальное введение в ротовую полость мелкого песка вызывает классическую рефлекторную реакцию отторжения за счет усиленной саливации. Отмывающий эффект слюны у ягнят пропадает по мере их взросления, а у взрослых животных отсутствует полностью.
У других хорошо адаптированных к жизни в пустыне животных такой особенности ротового пищеварения не обнаружено. К примеру, пустынные песчанки (Gerbillinae) после перевода с влажного корма на сухой демонстрируют классическую ответную реакцию застенных слюнных желез в форме усиления саливации. За счет усиления саливации они очищают ротовую полость от песка. Здесь будет интересным заметить, что у песчанок имеется своя особенность приема корма. Прежде чем взять корм в рот, песчанки тщательно отряхивают его от песка. Это поведенческая деталь имеет своей целью экономию слюны и, следовательно, воды. Суммарный объем выделяемой слюны при приеме корма у всех пустынных животных меньше, чем, скажем, у собаки.
Общим свойством всех животных аридной зоны является и то, что у них количество выделяемого желудочного, поджелудочного и кишечного сока больше по сравнению с животными умеренных зон. Следовательно, пониженная саливация у них компенсируется секреторной активностью желудка, кишечника, а также застенными пищеварительными железами — поджелудочной железой и печенью. Поэтому химус тонкого отдела кишечника у обитателей пустынь имеет жидкую консистенцию и представляется гомеостатичной средой даже в условиях экстремальных температур и дефицита воды.
Показательно, что в метаболизме пустынных животных вода используется с большой эффективностью. У каракульских овец объем воды, которая всасывается через слизистую оболочку желудочно-кишечного тракта, в 2-3 раза превышает объем выпитой воды. Опыты с радиоактивной водой (помеченной тяжелым водородом) показали, что одна и та же молекула воды появляется в просвете желудочно-кишечного тракта до 10 раз, прежде чем она попадает в состав мочи или кала и выводится из организма. У верблюда даже после 14-дневного лишения воды содержание воды в составе химуса тонкого отдела кишечника не изменилось и составляло около 97 %. Кишечный гомеостаз поддерживался и за счет того, что поступление химуса в тонкий отдел кишечника из сычуга при дефиците воды сильно ограничивался.
У пустынных животных специфично и пищеварение в толстом отделе кишечника. Прежде всего, здесь происходит интенсивная абсорбция воды из химуса. У пустынных животных кал содержит намного меньше воды по сравнению с животными умеренного климата. Если у лабораторной крысы содержание воды в кале составляет 70 %, то у мешотчатого прыгуна — всего 45 %. Тот факт, что пустынные животные при депривации воды погибают зачастую не от дегидратации, а от закупорки кишечника чрезмерно твердым калом, подчеркивает приоритетность экономии воды в метаболизме и важную роль толстого отдела кишечника в рационализации использования воды.
Еще одной специфической особенностью физиологии желудочно-кишечного тракта животных аридной зоны является его необычайно высокая экскреторная активность. Дефицит воды неизбежно приводит к повышению осмоляльности внутренней среды. Однако неизменность осмотического давления протоплазмы и межклеточной жидкости представляет одно из обязательных условий жизнедеятельности клетки. Повышение осмоляльности крови, лимфы и других биологических жидкостей несовместимо с жизнью высших животных. Именно по этой причине осмотическое давление крови выступает как жесткая константа гомеостаза. Каким же образом при дефиците воды удается удержать осмос в рамках физиологической нормы?
В обычных (неэкстремальных) условиях лишние соли и мочевина выводятся из животного организма через почки в составе мочи. У животных аридной зоны образуется небольшое количество мочи. Поэтому избыток солей и катаболитов у них удаляется другим путем, а именно через органы пищеварения. У большой песчанки (Rhombomys opimus) — удобного объекта для моделирования водного голодания — интенсивность диуреза чрезвычайно низкая. В отсутствие воды количество мочи у нее дополнительно сокращается. Но на этом фоне происходит 100 %-ное увеличение зольности кала, т. е. функцию экскреции солей берет на себя желудочно-кишечный тракт и прежде всего толстый отдел кишечника. Кроме солей через стенку пищеварительного канала удаляется из организма и мочевина.
В целом, у животных аридной зоны зафиксирована высокая толерантность к мочевине. У пустынных грызунов концентрация мочевины в крови при дефиците воды возрастает 10-кратно без вреда для организма. При этом мочевина в больших количествах выделяется с калом. Мочевина экскретируется в составе различных пищеварительных соков (слюна, желудочный, кишечный, панкреатический сок, желчь) и непосредственно через слизистую оболочку по всей длине желудочно-кишечного тракта. При водном голодании мочевину в повышенных концентрациях находят даже в составе желудочного химуса.
Таким образом, система пищеварения у животных аридной зоны принимает активное участие в экономии воды и поддержании водно-солевого гомеостаза.
Системы мочеобразования и выделения обитателей аридных зон обладают целым рядом специфических морфологических черт. Так, по сравнению с животными увлажненных биотопов у их почек лучше развит мозговой слой. Это происходит за счет увеличения прямоканальцевой части нефрона и положительно коррелирует со способностью концентрировать мочу. Например, у льва (Panthera leo) толщина мозгового слоя почек составляет 34 мм, тогда как у домашней свиньи она достигает всего 6,5 мм.
Соотношение толщины коркового и мозгового вещества в почках различных млекопитающих.
Помимо структурных особенностей у животных засушливых областей найдены функциональные особенности мочевыделительной системы. У мешотчатых прыгунов мочевой пузырь обладает выраженной способностью реабсорбировать воду из состава вторичной мочи. Концентрация вазопрессина у прыгунов повышена. Концентрация данного гормона в их моче составляет 50 ед/мл, тогда как у лабораторных крыс — всего 5-7 ед/мл. В ткани гипофиза мешотчатых прыгунов содержание вазопрессина равно 0,9 ед/мг, у лабораторных крыс — в три раза меньше (0,3 ед/мг). При недостатке воды различия между этими животными сохраняются, хотя секреторная активность нейрогипофиза усиливается как у одних, так и у других. За сутки у мешотчатых прыгунов образуется всего несколько капель мочи при очень высокой осмоляльности, которая достигается за счет высокой концентрации мочевины (23 %) и хлорида натрия (9 %). Для наглядности укажем, что у лабораторных крыс количество образующейся за сутки мочи на три порядка выше при более низкой концентрации мочевины и хлорида натрия. Отметим, что осмоляльность мочи у собаки и человека в 1,5-2 раза ниже, чем у лабораторной крысы.
Одна из самых высоких способностей почек к реабсорбции воды обнаружена у спинификсовой прыгающей мыши (Notomys alexis). Осмотическое давление мочи этого животного в 3 раза выше по сравнению с мочой мешотчатого прыгуна и достигает 9370 мм/осм/л. В целом потеря живой массы при депривации воды у аридных животных относительно низкая. Если верблюд за рабочий день, получая лишь сено низкого качества, теряет 2-3 % от живой массы, то лошадь и осел в тех же условиях за счет обезвоживания потеряют 6-8 % живой массы.

Как известно, испарение воды с поверхности кожи и слизистых в условиях высоких температур воздуха предотвращает перегрев организма. Однако в большинстве районов нашей планеты с жарким климатом возникает дефицит воды, поэтому у животных аридных зон в процессе эволюции выработался очень эффективный механизм экономии воды, и прежде всего за счет испарения. Масштабные исследования показали, что кожно-легочные потери воды у обитателей аридных зон приблизительно на 1/3 ниже, чем у домашних животных.
В основе экономии на испарении лежат морфофункциональные особенности кожи и органов дыхания пустынных животных. У последних кожа сухая и лишена потовых желез. Развитый кожный эпидермис создает дополнительную защиту и противодействует испарению воды. Такой же эффект создает и небольшая жировая прослойка вдоль хребта, которая имеется у большинства животных сухих областей. Нельзя недооценивать и роль органов дыхания в экономии воды. Внешне это проявляется в том, что влажность выдыхаемого воздуха у пустынных животных регулируется в зависимости от температуры окружающей среды и доступности воды.
Экономия на испарении воды через кожу обнаружена у всех пустынных грызунов — крыс, песчанок, сусликов, мышей. Например, при температуре воздуха 30 °С у лабораторной крысы и большой песчанки (Rhombomys opimus) при равной живой массе и приблизительно одинаковой площади поверхности тела отмечается 2-кратная разница в количестве испарившейся воды за единицу времени. Причем у крысы повышение температуры воздуха от уровня температурного преферендума до +30 °С сопровождается резким возрастанием испарения воды через кожу и легкие. При температуре 30 °С разница у этих двух животных в суммарной потере воды приближается к 100 %. Песчанка расходует на терморегуляцию гораздо меньше воды. При повышении температуры с 10 до 30 °С затраты воды у нее увеличиваются не более, чем на 10 %.
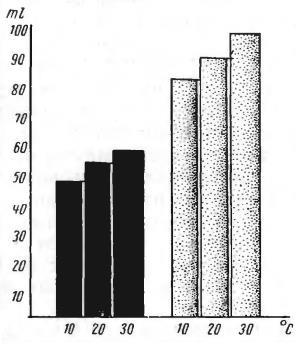
Кожно-легочные потери воды у большой песчанки (Rhombomys opimus) (слева) и серой крысы (Rattus norvegicus) (справа) при температуре окружающей среды 10, 20 и 30 °С.
Кожно-легочная экономия воды имеет место и у других видов животных, приспособившихся к жизни в засушливых условиях. При ограничении водопоя овцы понижают потери воды через органы дыхания на 20-25 %. У курдючных овец после 4-дневного водного голодания при температуре воздуха 31-33 °С относительная влажность выдыхаемого воздуха понизилась с 78 % до 62 %. У каракульских овец в похожих условиях наблюдений отмечали понижение влажности выдыхаемого воздуха на четверть. Такой же, но еще более эффективный механизм водной экономии демонстрируют и верблюды. В условиях доступности питьевой воды относительная влажность выдыхаемого этими животными воздуха составляет 75 % на фоне 90 %-ной относительной влажности атмосферного воздуха. Но при водном голодании выделение воды из организма верблюдов резко сокращается, о чем свидетельствует относительная влажность выдыхаемого воздуха (менее 30 %).
Каковы же физиологические механизмы сокращения потерь воды через органы дыхания? Их, по меньшей мере, два. Во-первых, экономию обеспечивает снижение легочной вентиляции в условиях дефицита воды. Пустынные животные способны понижать основной обмен при экстремальном повышении температуры среды. Естественно, что следствием снижения основного метаболизма является снижение потребления клетками и тканями кислорода. Угнетение дыхания при повышении температуры воздуха приводит к сокращению объема выделяемых с выдыхаемым воздухом паров воды. Во-вторых, экономия воды достигается и за счет специфических изменений морфологии и функций органов дыхания. У животных аридной зоны верхние дыхательные пути имеют большую длину и площадь слизистой оболочки. Такие животные, как верблюды (Camelus), жираф (Giraffa camelopardalis), джейран (Gazella subgutturosa), сайгак (Saiga tatarica), курдючная овца, аборигенные козы, имеют длинную шею (следовательно, и длинную трахею) и горбатый нос.

Нос сайгака (Saiga tatarica) как приспособление для сбережения воды.
При вдохе по мере движения горячего воздуха по верхним дыхательным путям происходит его охлаждение за счет испарения воды со слизистых оболочек. Однако образующийся конденсат остается в пределах верхних дыхательных путей. Механизм удержания конденсата имеет биохимическую основу. Вода связывается слизью, которой всегда забиты носовые ходы пустынных животных. Мукополисахариды — структурные элементы слизистых образований — в силу своих особых физикохимических свойств служат как акцепторами, так и донорами молекул воды. К регуляции этих процессов имеют отношение ферменты паракринной секреции. Так, известна причастность фермента гиалуронидазы к абсорбции слизью водяных паров. Экспериментально доказано, что активность фермента возрастает у пустынных животных в условиях водного голодания и высоких температур среды.
Помимо верхних дыхательных путей в процессе удержания воды участвуют и легкие. В условиях водного ограничения животных в легочном эпителии увеличивается число макрофагов, которые к тому же изменяются функционально. У верблюда после 2-3-недельной водной депривации в легких обнаруживаются скопления макрофагов, часть которых из фагоцитов превращаются в пиноциты. Гистохимические исследования свидетельствуют о том, что этот процесс трансформации макрофагов сопровождается потерей большей части липидов. Взамен жировых капель клетки приобретают специфические вакуоли, которые позволяют им пиноцитировать воду. Клетки с такими свойствами появляются не только в составе стенки, но и в просвете легочных альвеол верблюда и песчанки. Интересно и то, что эпителий, который выстилает бронхиолы легких пустынных животных, — цилиндрический. Плоский эпителий появляется лишь в самих альвеолах.
Венозная кровь, охлажденная в результате испарения воды в новых полостях, принимает участие в терморегуляции головного мозга. Этот процесс хорошо изучен у африканских газелей. Мозг получает кровь по сонным артериям. Но у газелей и, вероятно, некоторых других обитателей аридной зоны внутренние сонные артерии у основания черепа разделяются на сотни мелких артерий и проходят через венозный синус, по которому протекает охлажденная венозная кровь от верхних дыхательных путей. Данная магистраль охлаждает артериальную кровь на 2-3 °С. Из синуса мелкие артерии уходят в сторону мозга. Об эффективности механизма охлаждения говорит следующий факт. У газели Томсона (Gazella thomsoni) во время бега со скоростью 40 км/ч обмен веществ возрастает в 40 раз. Температура тела у животного при этом повышается с 39 С до 44 °С. Однако к головному мозгу артериальная кровь поступает с температурой менее 41 °С.
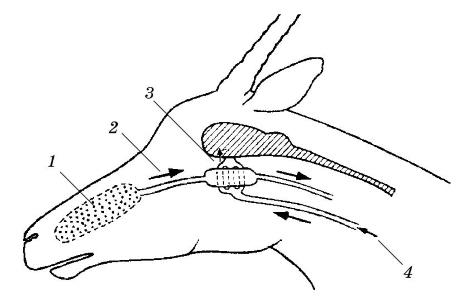
Схема охлаждения головного мозга: 1 — испарение; 2 — холодная венозная кровь; 3 — охлажденная артериальная кровь; 4 — теплая артериальная кровь.
Как следует из сказанного, морфофункциональные особенности органов дыхания позволяют пустынным животным экономить воду и одновременно сохранять высокую жизнеспособность организма даже в условиях экстремально высоких температур среды обитания.
вперед
в оглавление