в оглавление
назад
Происхождение и эволюция (продолжение)
Прогрессивная эволюция млекопитающих была связана с приобретением таких важных приспособлений, как истинная теплокровность и высокий уровень обмена веществ. Этому способствовали изменения в дыхательной и кровеносной системах, что выразилось в разделении сердца на четыре камеры и в сохранении одной дуги аорты, обусловивших несмешиваемость артериальной и венозной крови, в усложнении кожных покровов, играющих важнейшую роль в терморегуляции, в наличии вторичного костного неба, обеспечившего возможность дыхания во время еды, в изменении эффективности кормовой стратегии, обеспечившей ускоренное переваривание пищи. Совершенная кормовая стратегия оказалась возможной вследствие образования высокофункционального челюстного сустава, усложнения челюстной мускулатуры и дальнейшей дифференцировки зубов. Адаптивные возможности млекопитающих резко возросли после появления у них способности к живорождению и молочному выкармливанию детенышей. Наконец, решающим для эволюционного успеха группы стало последовательное усложнение органов чувств, центральной нервной системы (коры больших полушарий) и гормональной системы. В совокупности все это привело не только к совершенствованию организма как целостной системы, но и обеспечило сложное поведение. Следствием было развитие связей индивидов и образование сложных динамических группировок. Подобная "социализация" отношений в популяциях млекопитающих создала новые возможности в борьбе за существование и положение в биоценозах.
Самые ранние млекопитающие предположительно появились в среднем триасе, а их достоверные остатки известны из позднего триаса. Большинство мезозойских представителей класса описано по изолированным зубам и фрагментам черепа; лишь для некоторых известно строение посткраниального скелета. Они представляли собой мелких или средних по размерам (от землеройки до крысы, только немногие достигали величины барсука) коротконогих и длиннохвостых животных, покрытых короткой шерстью. У первых млекопитающих имелись сильно вытянутая морда, зачаточные ушные раковины и небольшие глаза. Для черепа характерны довольно крупная (минимум в 3-4 раза больше, чем у цинодонтов) мозговая камера и незамкнутая сзади глазница. Основные черепные признаки маммальной организации — в первую очередь, наличие костного среднего уха, усложненной зубной системы, а также ряд специфических особенностей строения боковой стенки черепа — развивались постепенно, причем в разных филетических линиях — в различной последовательности. Например, в разных подклассах неодинаково шло становление окклюзии, или постоянства смыкания зубных коронок.
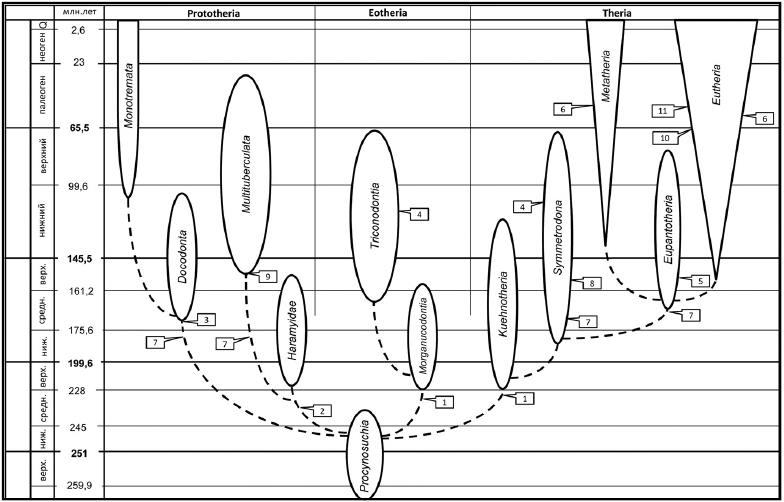
Ранняя радиация класса млекопитающих и время появления их основных признаков: 1 — тритуберкулярные (трехбугорчатые) щечные зубы; 2 — многобугорчатые коронки щечных зубов; 3 — "докодонтовый" тип коронки щечных зубов; 4 — нарушение симметродонтии: верхние щечные зубы расширяются трансверзально, нижние — вытягиваются вдоль оси челюсти; 5 — формирование трибосфенических щечных зубов; 6 — возникновение квадритуберкулярных верхних щечных зубов за счет разрастаний гипокона; 7 — редукция постдентальных костей и фомирование сустава нижней челюсти млекопитающих по схеме dentale-squamosum, увеличение массивности сочленовного отростка; 8 — появление углового отростка нижней челюсти; 9 — возникновение диастемы, гипертрофированное развитие резцов; 10 — формирование совершенной плацентарности: набор морфологических и физиологических признаков длительного вынашивания эмбриона и рождения крупных детенышей; 11 — резкое увеличение размеров головного мозга.
У синапсид зубы сменялись до 6 раз; неоднократная их замена зафиксирована и у базального млекопитающеобразного синоконодона (Sinoconodon rigneyi). В дальнейшем устанавливается однократная замена зубов. Ограничение числа зубных генераций обеспечило окклюзию, что косвенно свидетельствует о ее значении для эффективной обработки пищи. Вероятно, именно с формированием специализированного постоянного зубного аппарата, открывающего млекопитающим широкие возможности для пищевых адаптаций, связано прекращение их роста по достижении половой зрелости, чего не наблюдается у рыб, амфибий, рептилий, синапсид и уже упомянутого синоконодона. В самом деле, древнейшие млекопитающие питались беспозвоночными, но вскоре начали появляться плотоядные, всеядные и растительноядные формы.
В позднем триасе существовало по меньшей мере 2 отряда, уже приобретших набор характерных маммальных признаков, — гарамийиды (Haramiyida) и кюнеотерииды (Kuehneotheriida). К концу юры млекопитающие были уже достаточно разнообразны таксономически. На протяжении юрского периода возникло не менее 8 отрядов; на самом деле их, вероятно, было больше, поскольку первые однопроходные (Monotremata), скорее всего, также имеют юрский возраст. Гарамийиды не перешли рубеж триаса и юры; ряд возникших в средней юре отрядов существовал на протяжении большей части мела, т. е. около 100 млн. лет (эвтриконодонты (Eutriconodonta), спалакотериоиды (Spalacotherioidea), дриолестиды (Dryolestida)). Наибольшими же "долгожителями" среди всех млекопитающих являются многобугорчатые (Multituberculata): наиболее ранние их представители (из семейства плагиаулациевых (Plagiaulacidae)) известны из средней юры, а наиболее поздние (из семейств эвкосмодонтовых (Eucosmodontidae) и тениолабовых (Taeniolabididae)) — из среднего палеогена, так что общая протяженность существования этого отряда составила около 120 млн. лет. Для сравнения можно указать, что возраст самых старых современных отрядов едва ли намного превышает 70 млн. лет.
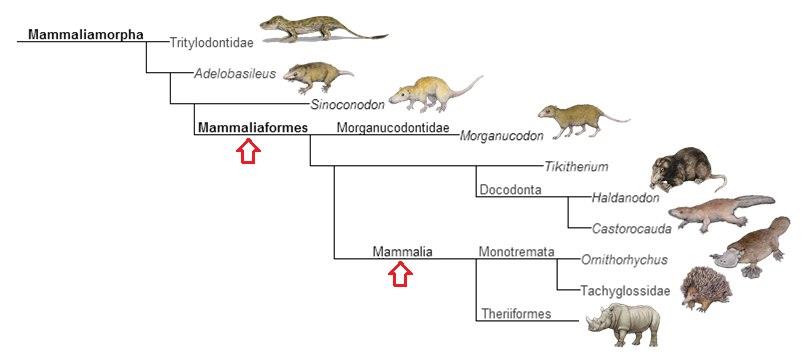
Родственные связи между млекопитающеобразными и собственно млекопитающими.
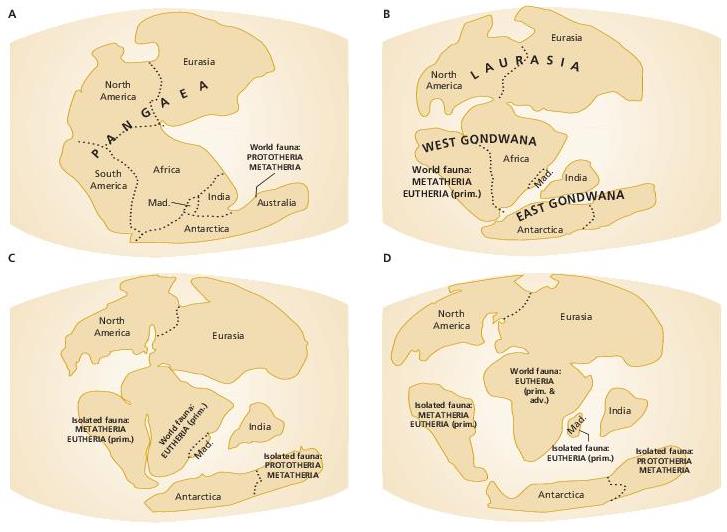
К позднетриасово-среднеюрской радиации млекопитающих относится выделение нескольких крупных филогенетических линий подклассового ранга. Наиболее значимым событием чаще всего считается базальная радиация общего ствола на две основные клады — австралосфенид (Australosphenida) (включает подкласс прототериев, или первозверей (Prototheria), с отрядом Monotremata, а также несколько небольших вымерших групп) и териеобразных (Theriimorpha) (объединяет подавляющее большинство остальных млекопитающих). В ряде палеогеографических реконструкций обособление австралосфенид связывается со средне-позднетриасовым разделением Пангеи на Гондвану (южный континент) и Лавразию (северный континент), что позволило дать кладе именно такое название. В градистских системах, как классических, так и новейших, к прототериям наряду с однопроходными иногда относят и других низших млекопитающих — многобугорчатых (Multituberculata) и трехбугорчатых (Triconodonta). В кладистических системах эти две группы считаются возникшими независимо от прототериев кладами аллотериев (Allotheria) (с отрядами Haramiyoidea и Multituberculata) и триконотериев (Triconotheria) (с единственным отрядом Triconodonta); в данной работе они рассматриваются в качестве подклассов в составе клады Theriimorpha.
Существует точка зрения, согласно которой австралосфениды выделились из среды синапсид самостоятельно, безотносительно к териеобразным. Результаты тонкого сравнительного анализа морфологии черепа (прежде всего, височной области и среднего уха) указывают на их большее сходство с динообразными, нежели с цинодонтами. В таком случае австралосфенид можно рассматривать как порождение удаленного края того обширного фронта эволюции синапсид, на котором разворачивался процесс маммализации. Если эта точка зрения верна, млекопитающие являются полифилетической группой, представленной 2 независимыми эволюционными линиями.
Ключевая для млекопитающих группа голотериев (Holotheria) включает всех их представителей с полностью развитыми признаками маммальной организации. Примитивные представители этой группы (отряды кюнеотериид (Kuehneotheriida), амфитериид (Amphitheriida), перамурид (Peramurida) и дриолестид (Dryolestidа)) существовали долго — начиная с позднего триаса и до позднего мела. Ранее всех их объединяли в группу эвпантотериев (Eupantotheria), но ныне она признана парафилетической и в кладистических классификациях отсутствует. Эволюция (и соответствующая ей кладистическая макросистема) голотериев связана с последовательным формированием трибосфенической зубной коронки. К ключевым событиям среднеюрской и раннемеловой радиации голотериев относится последовательное обособление клад трехнотериев (Trechnotheria), кладотериев (Cladotheria), цатериев (Zatheria) и, наконец, трибосфенид (Tribosphenida) с полностью развитыми, как видно из названия, признаками трибосфенических моляров. Ранние голотерии (в классических системах их обычно объединяют в инфракласс пантотериев (Pantotheria)) известны из Европы, Азии, Северной Америки, так что первые (среднеюрские-раннемеловые) этапы их становления, вероятно, связаны с «северной сушей» — Лавразией.
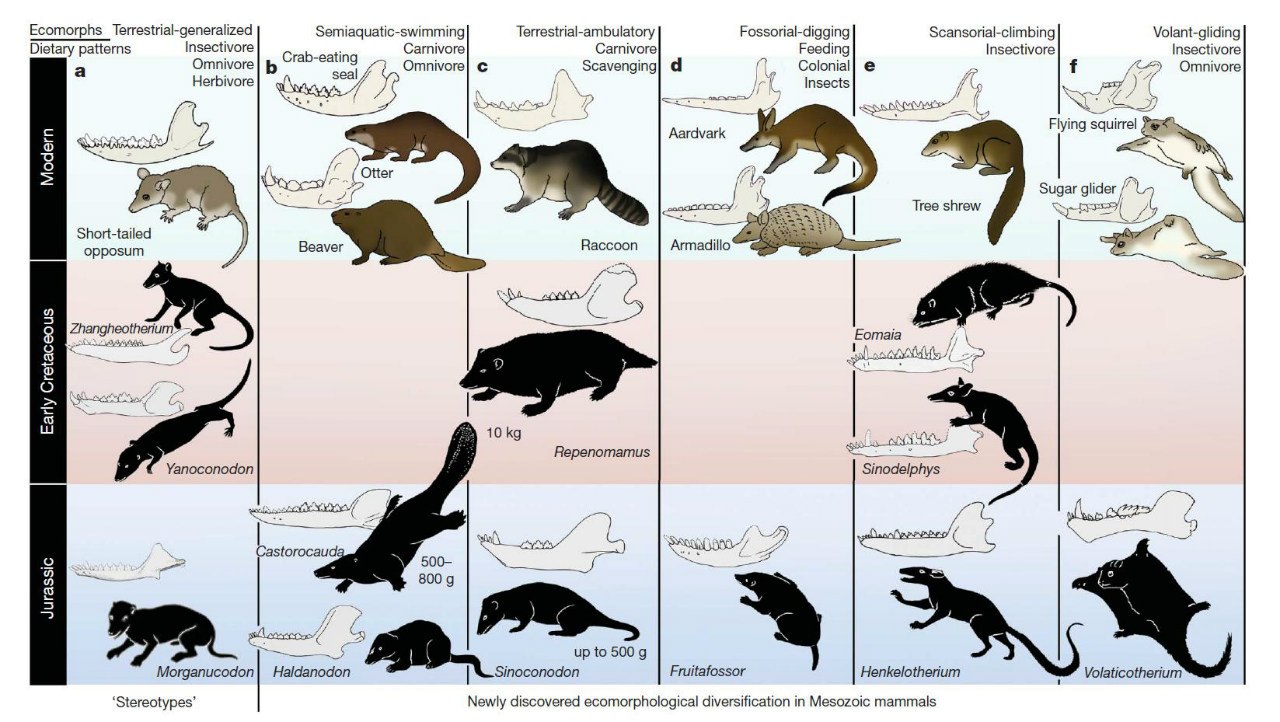
Экологические аналоги некоторых современных млекопитающих из юры и раннего мела.
В кладу Tribosphenida зачисляют метатериев (Metatheria) и эвтериев (Eutheria), представляющих собой соответственно сумчатых и плацентарных в широком смысле этих понятий. Базальная радиация трибосфенид начинается формированием нескольких групп, в классических системах относимых к т. н. метатериево-эвтериевой граде. Среди них наиболее известны меловые дельтатероиды (Deltatheroida), которых одно время в качестве предковых сближали то с Metatheria, то с Eutheria (здесь они рассматриваются в качестве базальных метатериев). Практически все самые ранние ископаемые находки трибосфенид датируются ранним мелом, но на самом деле эта клада, возможно, значительно древнее: указанием на это могут служить среднеюрские остатки из Китая, предположительно отнесенные к эвтериям, — юрамайя (Juramaia sinensis). Если это верно, то и обособление, и начальная дивергенция трибосфенид произошли, возможно, уже в юре.
Наиболее древние метатерии достоверно известны из раннего мела Китая (синодельф (Sinodelphys szalayi)); их первые (и гораздо более обильные) остатки из Северной Америки немногим моложе. Эти факты не совсем вписываются в наиболее популярную на сегодняшний день схему, согласно которой сумчатые начали формироваться на территории, соответствующей нынешнему Новому Свету, откуда проникли в Австралию (через Антарктиду) и в Старый Свет (известны начиная со среднего палеогена по ранний неоген Европы, Азии и Северной Африки). По-видимому, не позднее среднего мела метатерии разделились на 2 основные ветви — америдельфий (Ameridelphia) и австралодельфий (Australidelphia): первые известны из обеих Америк, Старого Света и Антарктиды; вторые — из Антарктиды, Австралии и южной части Южной Америки. Всего за историю сумчатых возникло 8-9 отрядов, из которых до современности не дошли только 2.
В ходе десятков миллионов лет самостоятельной эволюции метатерии и эвтерии достигли различных, но во многом принципиально сходных результатов. Например, решая задачу приспособления к поеданию растительной пищи, каждая ветвь приобрела ферментные системы в переднем и заднем отделах кишечника, используя для этого совершенно разные микробные сообщества. Обе ветви сходным образом решили проблему стирания зубов твердыми растительными тканями. Некоторые представители этих групп даже приобрели чрезвычайно сильное внешнее сходство. Однако в свете имеющихся данных приходится признать, что эволюция сумчатых по сравнению с основной массой плацентарных протекает гораздо медленнее, а сами они в среднем отличаются более низким уровнем приспособляемости. В связи с данными обстоятельствами в большинстве областей земного шара сумчатые не смогли успешно конкурировать с плацентарными и исчезли.
Южная Америка послужила убежищем для многих сумчатых на протяжении большей части кайнозоя, когда этот континент был изолирован от остального мира. Здесь возникли разнообразные группы сумчатых, главным образом хищных, которые почти полностью вымерли, когда восстановился Панамский перешеек и множество более высокоразвитых плацентарных заселило эту часть света. Единственный континент, где сумчатые достигли настоящего расцвета — это Австралия. Как показывают геологические данные, она отделилась от остальной суши в меловом периоде и с тех пор остается изолированной. По-видимому, к моменту отделения ни одно плацентарное млекопитающее не успело проникнуть в Австралию, и лишь немногим удалось добраться до нее впоследствии. Здесь сумчатые практически не имели соперников; они свободно радиировали и достигли такого разнообразия, что заполнили почти все экологические ниши, которые плацентарные млекопитающие занимают в других областях.
За все время существования эвтериев сформировалось около 40 таксонов отрядного ранга; из них к современной (голоценовой) фауне относится порядка 30 отрядов. Появление большинства из них датируется ранним палеогеном (собственно говоря, этим во многом и задается граница между мезозоем и кайнозоем); время существования современных отрядов надежно прослеживается до середины-начала палеогена. Среди последних древнее всех (по времени появления в палеонтологической летописи) хоботные (Proboscidea), некоторые насекомоядные (в широком понимании), грызуны (Rodentia), приматы (Primates), хищные (Carnivora) и китообразные (Cetacea). Лишь немногие отряды эвтериев (каждый представлен единственным семейством) существовали сравнительно недолго: арктостилопиды (Arctostylopida) и ксенунгуляты (Xenungulata) ограничены ранним, а миксодонтии (Mixodontia) — средним палеогеном; представители отряда бибималагазий (Bibymalagasia) известны только из антропогеновых отложений Мадагаскара. В наше время эвтерии составляют ядро фауны млекопитающих и представлены большим разнообразием жизненных форм, занявших практически все ландшафты Земли. Несмотря на бурную эволюцию большинства эвтериев, некоторые современные их представители (например, тенреки (Tenrecidae) и щелезубы (Solenodontidae)) в сравнении с меловыми плацентарными изменились мало.
В оценках базальной радиации эвтериев единодушие отсутствует. В ставшей уже классической кладистической схеме она задается дихотомией ксенартр (Xenarthra) и эпитериев (Epitheria), которая наиболее соответствует большинству палеонтологических фактов и может быть датирована, вероятно, средним мелом. Этой схеме отчасти соответствует гипотеза, которая обосновывается молекулярно-генетическими данными, она также допускает раннее обособление ксенартр. Последние включают отряды броненосцев (Cingulata) и неполнозубых (Pilosa). К неполнозубым относятся ленивцы (Folivora) и муравьеды (Vermilingua). Ксенартры высокоспециализированны, но весьма разнообразны по экологическим адаптациям.

Во взглядах на эпитериев между морфологической и молекулярно-генетической схемами имеются существенные расхождения. Первая предполагает их разделение на унгвикулят (Unguiculata) и ферунгулят (Ferungulata), вторая — на афротериев (Afrotheria) и бореоэвтериев (Boreoeutheria). Примечательно, что в обоих случаях время разделения оценивается в 105-110 млн. лет, что соответствует середине мелового периода. Авторы молекулярно-генетической схемы связывают свою филогенетическую реконструкцию с изоляцией южного суперконтинент Гондваны от северной Лавразии и ее распадом на восточный афро-аравийский и западный южноамериканский блоки: предполагается, что афротерии сформировались на территории Афро-Аравии в изоляции от прочих эвтериев. Согласно данному подходу, афротерии включают современные отряды афросорицид (Afrosoricida), прыгунчиковых (Macroscelidea), трубкозубых (Tubulidentata), даманов (Hyracoidea), сирен (Sirenia) и хоботных (Proboscidea), из вымерших сюда входят птолемейиды (Ptolemaiida) и эмбритоподы (Embrithopoda).
В свою очередь, бореоэвтерии объединяют плацентарных, выделившихся и прошедших первоначальную радиацию на материках Северного полушария — лавразитериев (Laurasiatheria) и эвархонтоглиров (Euarchontoglires), разделившихся около 95 млн. лет назад. Первые включают эвлипотифлей (Eulipotyphla), ферэ (Ferae) и цетунгулят (Cetungulata), вторые — грызунообразных (Glires) и эвархонтов (Euarchonta). О месте некоторых отрядов в границах данной системы ведутся споры. Так, рукокрылые (Chiroptera) на основании наличия сходных морфологических признаков объединяются с тупайеобразными (Scadentia), шерстокрылами (Dermoptera) и приматами (Primates) в кладу архонтов (Archonta). Методами же молекулярной биологии рукокрылые определяются в Laurasiatheria, в рамках которых вместе с непарнопалыми (Perissodactyla), хищными (Carnivora) и панголинами (Pholidota) объединяются в группировку пегасоферэ (Pegasoferae). Прыгунчики (Macroscelidea) в морфологическом варианте систематики объединяются с грызунами (Rodentia) и зайцеобразными (Lagomorpha) в кладу анагалид (Anagalida), а по молекулярно-генетическим данным их относят к Afrotheria.
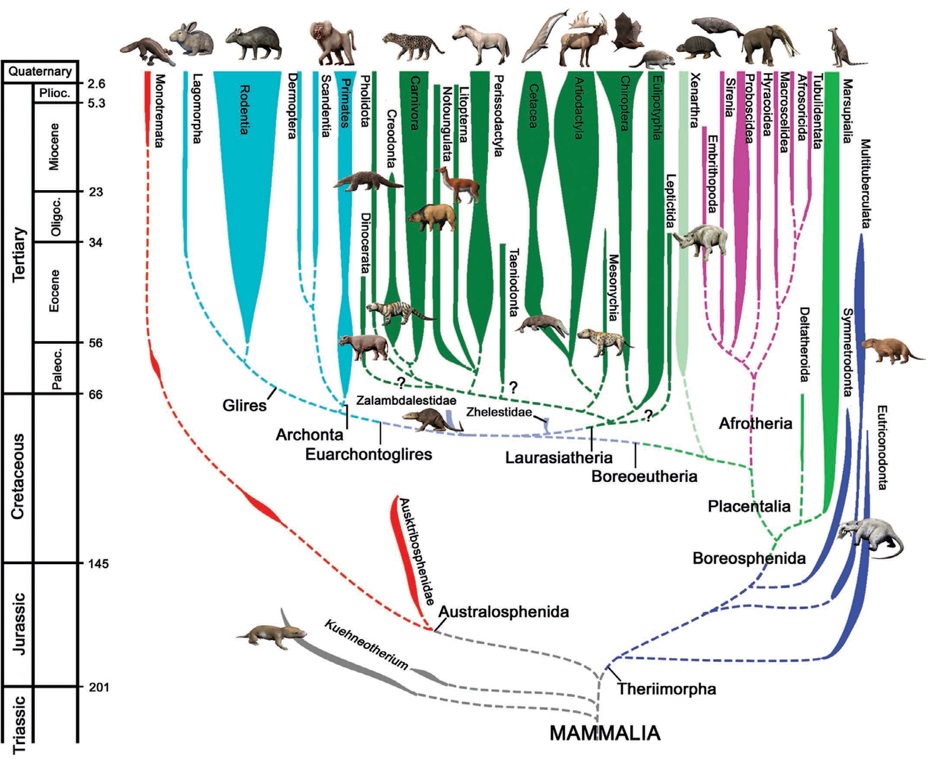
Существенно иная датировка и схема формирования основных групп эвтериевых млекопитающих предлагается в филогенетической схеме, основанной главным образом на палеогеографических реконструкциях. В ней ранняя радиация эвтериев датируется средней юрой и связывается с отделением древней Азии от блока Европа-Африка-Северная Америка. В основание восточной (азиатской) группы положены дельтатероиды (Deltatheroida): в нее входят хищные, рукокрылые, зайцеобразные и некоторые другие. В основание западной (евро-афро-американской) группы лежат дриолестиды (Dryolestida): в нее объединены приматы, грызуны, все унгуляты и ряд других. Впрочем, эта схема также весьма спорна.
Следуя канве кладистических реконструкций, важнейшим событием в истории плацентарных млекопитающих следует считать возникновение во второй половине мелового периода (возможно, раньше) группы Epitheria. Самые первые из них, лептиктиды (Leptictida), просуществовали около 90 млн. лет и вымерли во второй половине палеогена; вероятно, их архаичные меловые формы дали начало прочим эпитериям. Последние составляют подавляющее большинство современной фауны млекопитающих; они — единственные представители этого класса в Евразии, Африке и, за малым исключением, Северной Америке; кроме того, из млекопитающих только они в полной мере освоили водную и воздушную среду.
История формирования ключевых групп эпитериев далека от ясности, поскольку, как видно из вышеизложенных схем, весьма противоречиво трактуются отношения между основными кладами этой группы. Вероятно, причина состоит в том, что начальная дивергенция эпитериев носила взрывной (одномоментный по геологическим меркам) характер, а их морфологическую основу составляла довольно однородная в морфологическом отношении группа (типа примитивных насекомоядных в широком смысле), и потому последовательность событий не улавливается путем сравнительного анализа доступных признаков. Так что остается лишь констатировать, что к концу мезозоя некоторые, а в начале кайнозоя почти все основные филетические ветви (отряды и надотрядные группировки) эпитериев уже присутствуют в палеонтологической летописи. На этом основании можно с большой долей вероятности предполагать, что время их возникновения и начальной радиации — как минимум поздний, а то и средний мел.

К концу мезозоя млекопитающие были достаточно разнообразными, но все еще оставались мелкими. Некоторые представители класса из позднего мела - раннего палеоцена Северной Америки: слева — эодельфис (Eodelphis), вверху — птилоды (Ptilodus), справа — гобиаконодоны (Gobiaconodon).
Кладистическими средствами на морфолого-палеонтологических материалах в ранней эволюции эпитериев, по-видимому, достаточно обоснованно показано обособление эвлипотифлей (Euipotyphla) прежде расхождения других основных клад. Среди последних вполне надежно обосновывается единство происхождения каждой из четырех основных групп — анагалид (Anagalida), архонтов (Archonta), ферэ (Ferae) и унгулят, или копытных (Ungulata).
История становления насекомоядных в широком понимании (Insectivora) достаточно неопределенна, поскольку имеются значительные расхождения в оценке состава каждой из относящихся к этой кладе групп. По-видимому, они развивались 2 независимыми линиями, возникшими в среднем и позднем мелу. Первая линия соответствует насекомоядным в узком смысле, или эвлипотифлям (Eulipotyphla); сюда относятся землеройкообразные (Soricomorpha) и ежообразные (Erinaceomorpha). Вторая линия представлена тенрековыми (Tenrecoidea), объединяющими златокротов (Chrysochloridae) и собственно тенреков (Tenrecidae). Согласно молекулярно-генетической схеме тенреки переименованы в афросорицид (Afrosoricida) и отнесены к Afrotheria.
Анагалиды (Anagalida), известные уже с раннего палеогена, представлены несколькими отрядами, из которых до современности дожили 3 — прыгунчики (Macroscelidea) (по молекулярно-генетическим данным принадлежащие к афротериям), зайцеобразные (Lagomorpha) и грызуны (Rodentia). Грызуны очень широко радиировали: известно не менее 60 их семейств, из которых более 30 — современные. Это наивысший показатель среди нынешних млекопитающих, свидетельствующий о подлинном расцвете данной группы. Что касается зайцеобразных, то они никогда не составляли конкуренции грызунам в отношении разнообразия (только 2-3 семейства) и, судя по динамике родового состава, уже прошли пик своего эволюционного процветания: из 60 с лишним родов зайцеобразных в современной фауне осталось немногим более 10.
Приматы (Primates) — одна из наиболее рано радиировавших ветвей архонтов. Они возникли, вероятно, на территории Северной Америки, были достаточно многочисленны и вполне узнаваемы уже в начале палеогена. Приблизительно в это время произошло и их разделение на ключевые группы, в том числе отделение от основного ствола ранних шерстокрылов (Dermoptera). Впрочем, вопрос об "основном стволе" приматов остается открытым, поскольку неясна временная последовательность заселения эндемичными группами отдельных изолированных массивов суши — Южной Америки (широконосые обезьяны (Platyrhini)) и Мадагаскара (несколько надсемейств мокроносых обезьян (Strepsirrhini)). Всего за историю приматов возникло около 30 семейств, из которых лишь немногим более 10 достигли современности. Наиболее же разнообразным таксономически этот отряд был в середине палеогена, откуда известно не менее 15 семейств (в том числе уже несколько современных).

Разнообразие райнекайнозойских эвтериев (все изображенные животные известны из первой половины эоцена Европы и Северной Америки): 1 — Hassianycteris; 2 — Eurotamandua; 3 — Paramys; 4 — Oxyaena; 5 — Eobasileus; 6 — Eurolemur; 7 — Leptictidium; 8 — Propalaeotherium; 9 — Mezonyx; 10 — Phenacodus. Автор — С. Крускоп.
Рукокрылые (Chiroptera), в некоторых систематиках также относящиеся к архонтам, приобрели свои основные черты строения, связанные с полетом, уже в среднем палеогене, став в то время одними из наиболее специализированных эпитериев. Таким образом, они существуют без особых перестроек организации не менее 50 млн. лет и в настоящее время являются вполне процветающей группой.
Группа ферэ (Ferae) довольно разнородна по составу и характеру адаптаций. В нее входят, с одной стороны, креодонты (Creodonta) в широком смысле вместе с хищными (Carnivora), а с другой — цимолесты (Cimolesta) и близкие к ним панголины (Pholidota). Первые два отряда специализировались как плотоядные; вторая ветвь представлена преимущественно растительноядными (в виде исключения — насекомоядными) формами. В современной фауне из числа Ferae лишь Carnivora являются вполне процветающей группой.
Среди унгулят наибольшее значение (из-за широкого разнообразия) имеет обширная предположительно монофилетическая группа цетунгулят, или китокопытных (Cetungulata), в основе которое лежат палеогеновые кондиляртры (Condylarthra) в широком смысле. Последние сыграли воистину ключевую роль в становлении этой филемы копытных, из них легко выводится практически весь веер их филетических линий.
Наиболее широкую радиацию цетунгуляты дали в Южной Америке, где возникли 6 эндемичных отрядов, объединяемых в полифилитическую группу южноамериканских копытных, или меридиунгулят (Meridiungulata), — литоптерны (Litopterna), нотоунгуляты (Notoungulata), астрапотерии (Astrapotheria), пиротерии (Pyrotheria) и ксенунгуляты (Xenungulata). Наиболее значительного разнообразия достигали нотоунгуляты: в составе этого отряда выделяется несколько подотрядов и более десятка семейств, одно из которых даже смогло проникнуть в Северную Америку. Наряду с литоптернами нотоунгуляты дожили до позднего плейстоцена.
Постепенное увеличение размеров тела и завитие рогообразных выростов у бронтотериев (Brontotheriidae), снизу вверх: Eotitanops (ранний эоцен, 55 млн. лет назад); Manteoceras (начало среднего эоцена, около 50 млн. лет назад); Protitanotherium (конец среднего эоцена, 45 млн. лет назад); Megacerops (поздний эоцен, 35 млн. лет назад).
Из доживших до современности групп в филему Cetungulata включаются отряды парнопалых (Artiodactyla), китообразных (Cetacea), непарнопалых (Perissodactyla) и группировка пенунгулят (Paenungulata). В настоящее время имеются веские основания полагать, что первые два отряда входят в единую филетическую линию, которая получила название эпарктоцион (Eparctocyona), или китопарнопалых (Cetartiodactyla). Раннепалеогеновые находки ближайших еще четвероногих предков китов, по-видимому, подтверждают молекулярно-генетические данные и указывают на то, что их наземных предков следует искать среди не генерализованных мезонихий (Mesonychia), как полагали ранее, а среди каких-то наиболее архаичных раннепалеогеновых "парнопалых". Эти же данные весомо свидетельствуют в пользу монофилии Cetacea. Первыми среди китообразных были среднепалеогеновые археоцеты (Archeoceti), у которых еще сохранился развитый пояс задних конечностей и не было связанных с эхолокацией анатомических черт строения черепа.
В первой половине кайнозоя одной из наиболее разнообразных групп копытных являлись непарнопалые (Perissodactyla). Возникновение отряда датируется ранним палеогеном, в конце палеогена в нем насчитывалось не менее 12 семейств, но к середине неогена многие из них уже вымерли. В современной фауне сохранилось всего 3 семейства непарнопалых; отряд находится в состоянии эволюционного затухания. С непарнопалыми ныне сближают и вымерший отряд десмостилий (Desmostylia), в прошлом обычно зачисляемый в Afrotheria.
Весьма детально прослежено историческое развитие Paenungulata, включающих в современной фауне отряд даманов (Hyracoidea), хоботных (Proboscidea) и сирен (Sirenia). Это одна из немногих групп, в прочтении филогении которых почти нет разногласий. Они претерпели в первой половине кайнозоя поистине взрывную эволюцию. К базальной радиации этой группы относится разделение даманов (Hyracoidea), эмбритоподов (Embritopoda) и тетитериев (Tethytheria). Среди последних особенно примечательна история Proboscidea, в ходе которой возникло не менее 10 семейств. Их эволюция шла настолько быстро и сопровождалась столь глубокой специализацией, что большинство этих семейств включает лишь по 2-3 рода. Только занимающие вершину эволюционного древа хоботных семейства гомфотериевых (Gomphotheriidae) и слоновых (Elephantidae) радиировали более дробно: в каждом из них насчитывается не менее 10 родов.
Согласно молекулярно-генетическим данным, цетунгуляты — сборная группа: пенунгуляты с данной точки зрения должны быть отнесены к афротериям, а китопарнопалые и непарнопалые — к лавразитериям. В границах последних китопарнопалые (Cetartiodactyla) и пегасоферэ (Pegasoferae) объединены в группировку скротифер (Scrotifera).
вперед
в оглавление