назад
Происхождение и эволюция

Некоторые синапсиды и вымершие млекопитающие. Автор — М. Панова.
Птицы и млекопитающие представляют собой 2 вершины эволюционного древа позвоночных. Их развитие, однако, шло абсолютно независимыми путями, а высокий уровень организации был достигнут разными средствами. В некоторых отношениях млекопитающие даже уступают птицам. Так, их метаболизм менее интенсивен, а температура тела ниже; соответственно, ниже механическая мощность. Хуже, чем у птиц, развито и зрение млекопитающих. В значительной мере это обусловливается тем, что птицы произошли во 2-й половине мезозоя от наиболее прогрессивной группы рептилий — архозавров (Archosauria), а предшественники млекопитающих — синапсиды (Synapsida) — ответвились еще в среднем карбоне от близких к амфибиям предков. Именно этим можно объяснить наличие у млекопитающих не экономящих воду почек, железистой кожи и ряда других особенностей.
Уже первые синапсиды легко распознаются по характерному типу редукции покровных костей черепа и наличию с каждой его стороны единственного височного окна, расположенного ниже заглазничной кости. Окно это ограничивается единственной скуловой дугой, состоявшей, как и у млекопитающих, из отростка чешуйчатой кости и из скуловой кости. Также для синапсид был характерен целый ряд архаичных признаков: амфицельные позвонки, подвижные шейные и поясничные ребра, увеличенное количество пальцевых фаланг, полифиодонтная (многократно сменявшаяся) зубная система, дополнительные небные зубы, нижняя челюсть из семи костей, теменное отверстие, малые размеры головного мозга (относительно хорошо развитым был лишь мозжечок) и др.
Архаичные безусловные представители синапсид объединяются в группу сфенакообразных (Sphenacomorpha). Их выделение могло стать результатом приспособления к питанию крупной добычей. Это были первые настоящие хищники среди амниот. К концу карбона и особенно в начале перми группа претерпела бурный расцвет формообразования и создала богатую фауну из десятков родов. К этому времени существенно расширился адаптивный диапазон сфенакообразных, представленных как плотоядными (сфенакодоны (Sphenacodon), диметродоны (Dimetrodon) и др.), так и растительноядными (эдафозавры (Edaphosaurus) и др.) формами. Крупнейшие из этих животных превышали 3 м в длину. Обитали они преимущественно у водоемов, где находили пищу и укрытия.
Во многих отношениях сфенакообразные оставались достаточно примитивными: их конечности занимали расставленное или полувыпрямленное положение, височная впадина была небольшой, а скуловая дуга — широкой и массивной. Но сидящие в альвеолах зубы уже обнаруживали начальные этапы дифференцировки: на предчелюстных костях они напоминали резцы, за ними шли парные клыки, однако конические щечные зубы еще не были приспособлены к пережевыванию пищи. У многих сфенакообразных, представителей различных семейств, на спинной стороне имелась кожная складка, соединявшая необычайно высокие (до 1 м) остистые отростки туловищных позвонков, от плечевого пояса и по крестцовый отдел. При этом у части эдафозавров эти ости несли короткие поперечно ориентированные выступы. Высказывались разные предположения относительно назначения данного «паруса». Согласно одной из версий, складка имела богатое кровоснабжение и могла играть важную роль в поведенческой терморегуляции сфенакообразных как орган регулируемого поглощения тепла или, наоборот, теплоотдачи.
Для сфенакообразных было характерно отсутствие известной у многих низших тетрапод ушной вырезки на заднем краю крыши черепа, которая предположительно возникла на месте брызгальца, вмещала барабанную перепонку и занимала подходящее положение для передачи вибраций с перепонки через стремечко (рудимент подвеска (hyomandibulare)) на внутреннее ухо. По причинам, связанным с техникой обработки корма, челюстной сустав этих синапсид располагался очень низко, крупное стремя тянулось к нему, контактировало с квадратной и сочленовной костями. Возможно, оно передавало вибрации с грунта через нижнюю челюсть (сейсмический слух). На наружной стороне угловой кости сфенакообразные имели т. н. отогнутую пластинку, которая тонким и широким гребнем выступала от наружной стороны кости, располагаясь параллельно ее боковой поверхности. Как предполагают, под ней находилась воздушная полость, возникавшая как выпячивание канала брызгальца. Эта воздушная камера позволяла отогнутой пластинке колебаться, поддаваясь давлению звуковой волны, и могла служить резонатором. Таким образом, очевидно уже у сфенакообразных чувствительность к звуковым колебаниям в воздухе была приурочена к области челюстного сустава, тем самым наметились предпосылки к формированию здесь среднего уха.
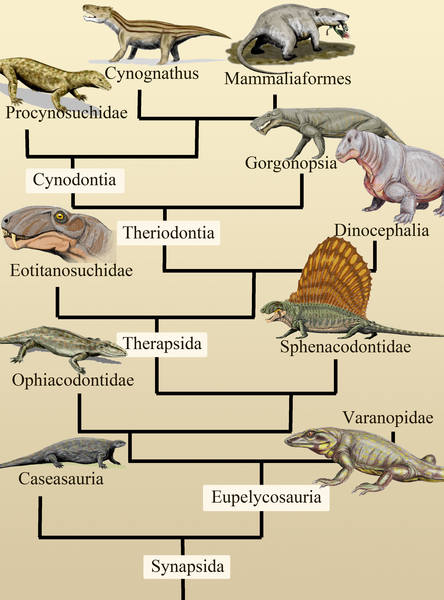
Обобщенное эволюционное древо синапсид.
В средней перми на смену сфенакообразным пришли их более прогрессивные потомки — динообразные (Dinomorpha). До среднего триаса они дали широкий веер форм, среди которых имелись как хищные, так и растительноядные. Наиболее мелкие из них были величиной с крысу, а крупнейшие достигали размеров слона.
У динообразных отогнутая пластинка угловой кости выражена значительно сильнее, чем у сфенакообразных, и ее значение как маркера области восприятия звуков из воздуха подтверждается выраженным в разной степени подразделением нижней челюсти на два отдела, непрочно связанных между собой. Это крупная зубная кость и расположенный позади нее комплекс небольших «постдентальных» костей — сочленовная, угловая, надугловая и еще две покровные кости. Квадратная кость непрочно связана с черепной коробкой, вместе со стремечком она входит в комплекс образований, предположительно передававших колебания внутреннему уху.
Вследствие податливости непрочно скрепленных между собой костных элементов в области челюстного сустава он не мог предоставлять нижней челюсти надежной опоры, необходимой для сильного сжатия добычи челюстями. В связи с этим динообразные использовали в основном горизонтальные, продольные движения и усилия нижней челюсти.
Так, растительноядные дицинодонтоиды (Dicynodontoidea), просуществовавшие с конца перми до конца триаса, были весьма разнообразны по размерам и строению тела, но имели довольно постоянные пропорции черепа из-за уникальной специализации ротового аппарата. Височная область черепа была у них удлинена, а скуловая дуга превращена в широкую горизонтальную полку, от обеих сторон которой вероятно начинались челюстные мышцы, ориентированные, судя по конфигурации черепа, почти горизонтально. От зубов чаще всего оставалась лишь пара гипертрофированных верхних клыков. Очевидно, впереди обе челюсти несли клювовидный роговой покров. Челюстной сустав отличался продольно растянутой суставной поверхностью на нижней челюсти. Ее рельеф свидетельствует о способности к широкому возвратно-поступательному скольжению, при котором острые лезвия роговых чехлов на челюстях могли резать растительный материал.
Другим примером адаптации к использованию продольных движений и усилий нижней челюсти из-за ослабления челюстного сустава могут служить горгонопиды (Gorgonopida) — доминирующие хищники средней и поздней перми, охотившиеся на крупную добычу. У них сильно выступали нижние клыки, а щечные зубы, как и у дицинодонтов, были слабы или отсутствовали. Зубная кость имела длинный венечный отросток (место крепления располагавшейся позади челюстной мышцы) и вместе с крупным клыком напоминала Г-образный багорик, что позволяло этим животным эффективно разрывать добычу резкими продольными рывками (или, возможно, сериями рывков, используя нижние клыки в качестве виброинструмента).
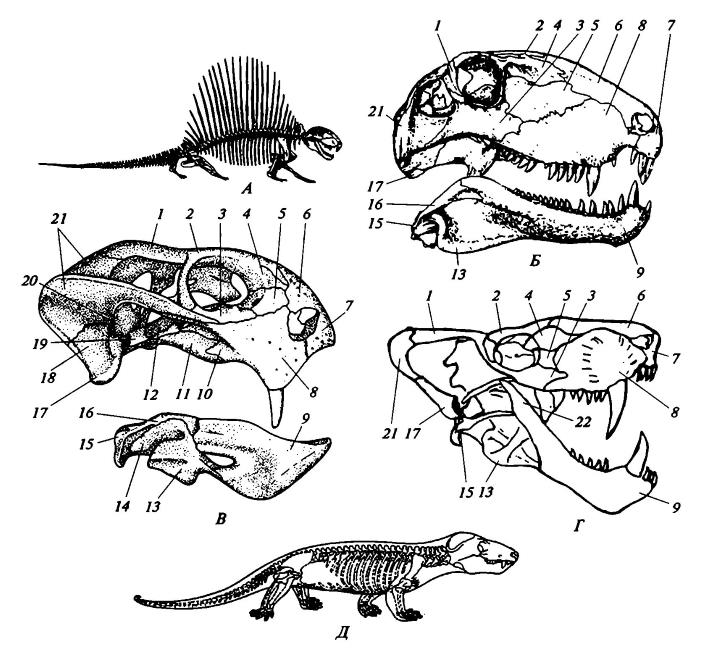
Морфологические особенности некоторых синапсид:
А — скелет Dimetrodon; Б — череп Dimetrodon; В — череп Dicynodon; Г — череп Sauroctonus; Д — скелет Cynognathus; 1 — заглазничная кость; 2 — лобная кость; 3 — скуловая кость; 4 — предлобная кость; 5 — слезная кость; 6 — носовая кость; 7 — предчелюстная кость; 8 — верхнечелюстная кость; 9 — зубная кость; 10 — поперечная кость; 11 — крыловидная кость; 12 — переднеклиновидная кость; 13 — отогнутая пластинка угловой кости; 14 — угловая кость; 15 — сочленовная кость; 16 — надугловая кость; 17 — квадратная кость; 18 — квадратноскуловая кость; 19 — верхнекрыловидная кость; 20 — переднеушная кость; 21 — чешуйчатая кость; 22 — венечный отросток зубной кости.
В середине перми от неспециализированных динообразных произошли терозавры (Therosauria), которые в итоге дали начало млекопитающим. При этом становление характерной для млекопитающих организации (маммализация) происходило параллельно в обеих их ветвях — у цинодонтов (Cynodontia) и тероцефалов (Therocephalia). Вторая ветвь оказалась тупиковой.
Цинодонты изначально представляли собой активных хищных или всеядных животных размером от крысы до собаки. В начале триаса они широко распространяются и полностью замещают горгонопсид. Среди цинодонтов появляются относительно специализированные крупные (величиной с медведя) формы, как растительноядные, так и плотоядные (например, циногнаты (Cynognathus)). К концу триаса разнообразие группы идет на убыль, но отдельные ее представители встречаются вплоть до раннего мела. Примечательно, что позднемеловая и раннепалеогеновая эволюция млекопитающих почти в точности воспроизвела эту картину адаптивной радиации: на основе небольших генерализованных всеядных животных возникли крупные специализированные растительноядные и вскоре вслед за этим питающиеся ими также крупные специализированные хищники.
Череп цинодонтов был, как правило, низким из-за малых размеров головного мозга, но очень широким, поскольку мощные скуловые дуги были сильно выгнуты в стороны, обрамляя заполненные мускулатурой обширные височные окна. Мускульные волокна тянулись на венечный отросток не только сзади и изнутри — от черепной коробки, но и снаружи — от скуловой дуги. В ходе эволюции совершилась переориентация последней мускульной порции — область отхождения ее волокон от черепа распространилась вперед с чешуйчатой кости на скуловую и даже на верхнечелюстную, что дало ей способность тянуть нижнюю челюсть вверх и вперед. Так возникла большая жевательная мышца (массетер), способная уравновесить направленную назад составляющую усилия архаичной части мускулатуры (височной мышцы) и вместе с ней тянуть нижнюю челюсть точно вверх, как это необходимо для обработки пищевых объектов щечными зубами (т. е. для жевания).
Развитие массетера компенсировало неуклонное ослабление первичного челюстного сустава через сочленовную и квадратную кости, не способных предоставить необходимую опору увеличивающейся зубной кости. В результате у высших цинодонтов зубная кость приобрела новую, независимую опору на осевой череп, а именно на чешуйчатую кость. Таким образом, образовался вторичный, или маммальный, челюстной сустав. Поначалу он был весьма примитивным. К примеру, у цинодонтов из семейства трителодонтовых (Tritheledontidae) зубная и чешуйчатая кости уже сочленялись, однако на первой еще не было сочленовой головки, а на второй отсутствовала выраженная сочленовная впадина. Некоторое время слабеющий первичный челюстной сустав продолжал функционировать наряду с усиливающимся вторичным. В конце концов у млекопитающих остался только вторичный сустав, а сильно уменьшившиеся квадратная и сочленовная кости превратились в элементы среднего уха (молоточек и наковальню соответственно).
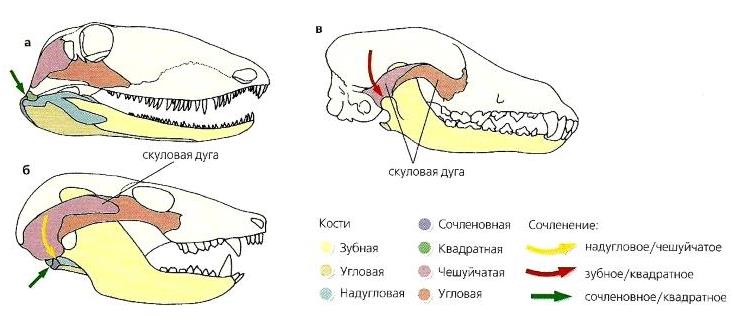
Эволюция шарнирного механизма соединения нижней челюсти с черепом. Нижняя челюсть раннепермского пеликозавра Ophiacodon (а) состоит из нескольких костей, в том числе сочленовной, угловой, надугловой и зубной. У среднетриасового цинодонта Probainognathus (б) сохраняется примитивное сочленение посредством сочленовной и квадратной (на черепе) костей, но в формировании сустава начинают участвовать новые кости — надугловая (на нижней челюсти) и чешуйчатая (на черепе), причем надугловая кость занимает это положение вследствие значительного разрастания зубной кости. Другим важным изменением стало развитие скуловой дуги, к которой прикрепляются мощные жевательные мышцы. У ныне живущих млекопитающих (показан волк (Canis lupus)) (в) имеется сочленение между зубной и чешуйчатой костями, причем зубная кость осталась единственной в нижней челюсти.
Формирование комплекса типичных для млекопитающих признаков происходило одновременно в нескольких филетических линиях цинодонтов (в большинстве своем окончившихся слепо), так сказать широким фронтом. Например, перестройка челюстной мускулатуры и формирование маммального челюстного сустава происходили независимо в 3 или 4 линиях. Также в нескольких линиях, причем не совпадающих с предыдущими, происходило становление дифференцированной зубной системы с острыми резцами, увеличенными клыками и бугорчатыми щечными зубами. Кроме того, цинодонты приобрели вторичное костное небо, которое укрепило челюстной аппарат и разграничило дыхательный и пищеварительный тракты. Это разделение позволило не прерывать дыхания при пережевывании пищи, а в дальнейшем — и при сосании молока детенышами. Для того и для другого развились мясистые губы и подвижный язык. На конце морды и по всей верхней губе сформировалось сенсорное поле и, возможно, сидящие на нем «протовибриссы». В носовой полости появилась верхняя обонятельная раковина, обеспечивающая обогрев и увлажнение вдыхаемого воздуха. Несколько расширились большие полушария головного мозга. Образовался двойной затылочный мыщелок.
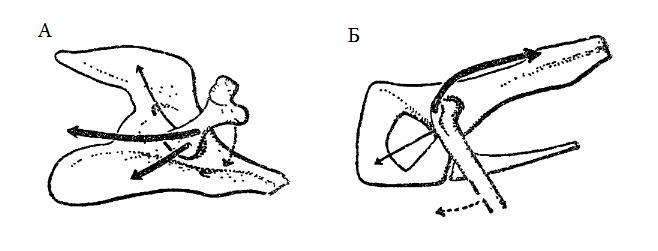
Схема, показывающая разницу в мускулатуре области тазобедренного сочленения у пеликозавра (А) и млекопитающего (Б). На А главные мышцы, двигающие бедренную кость вниз и назад (показаны жирными линиями), прикрепляются к хвосту и к вентральной части тазового пояса. На Б эти мышцы редуцированы и в значительной степени заменены мышцами, прикрепленными к большому вертелу и помещающимися на подвздошной кости.
У тероцефалов маммализация протекала не менее последовательно, но отличалась в деталях; к примеру, челюстного сочленения между зубной и чешуйчатой костью они так и не приобрели. В среднем триасе тероцефалы исчезли, не оставив потомков.
Сухой и жаркий климат триаса благоприятствовал эволюционному развитию и увеличению разнообразия тетрапод с завропсидными качествами (свойствами настоящих ящеров). Разнообразие и численность синапсид сократились, размеры их тела уменьшились. Они были вынуждены довольствоваться второстепенными нишами, что в частности выразилось в переходе к сумеречному и ночному образу жизни. Однако именно жизнь в таких условиях оказалась мощным стимулом к развитию более совершенных органов обоняния, слуха, осязания и параллельно — головного мозга. Как следствие, усложнилось поведение и общение между особями.
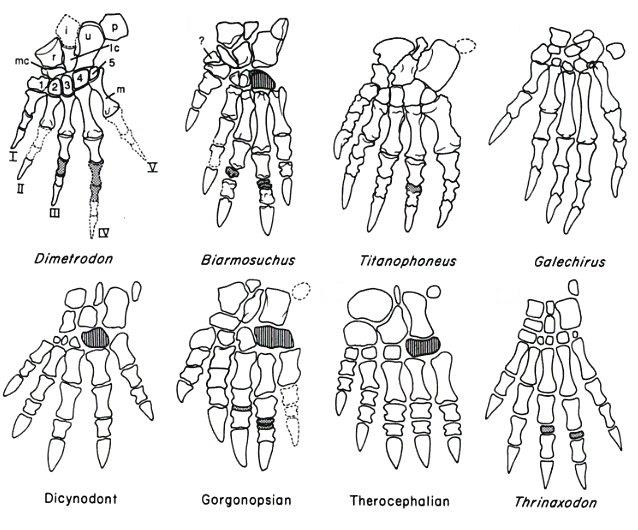
Строение кисти различных синапсид.
Наиболее высокоорганизованные цинодонты — представители семейств трителодонтовых (Trithelodontidae), дроматериевых (Dromatheriidae), бразилодонтовых (Brasilodontidae), териогерпетовых (Therioherpetidae) и некоторые другие — уже мало отличались от первых млекопитающих. Однако какая именно их линия привела к млекопитающим, пока неясно, поскольку предполагаемые предки демонстрируют различную степень маммализации. На их фоне млекопитающеобразные (Mammaliaformes) (отряды морганукодонтов (Morganucodonta) и докодонтов (Docodonta)) еще сохраняют некоторые архаичные морфологические черты, среди которых наиболее очевидная — рудименты постдентальных костей, еще не вошедших целиком в состав костного среднего уха.
Последовательность адаптивных преобразований, ведущих к образованию среднего уха млекопитающих, довольно ясна. Из отогнутой пластинки угловой кости возникли как барабанная перепонка, так и рамка для нее. Сочленовная кость на каких-то стадиях процесса тоже формировала заднюю часть рамки, а позже сохранила связь с центром барабанной перепонки, образовав путь для передачи вибраций через челюстной сустав на квадратную кость и далее по стремени к внутреннему уху. В окончательном виде у млекопитающих угловая кость получила название барабанной. Возникший из сочленовной кости молоточек, производная квадратной кости наковальня и стремечко очень мелки, что уменьшает обусловленное инерцией угасание передаваемых ими колебаний.
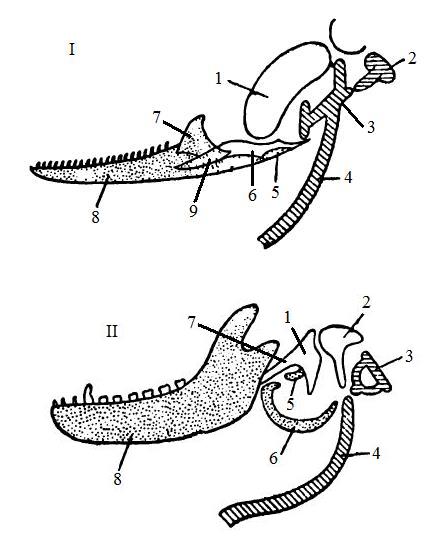
Сравнительная схема строения слуховых косточек и их происхождения у рептилий (I) и млекопитающих (II); пропорции не соблюдены.
I: 1 — квадратная кость (quadratum); 2 — столбик (columella); 3 — extracolumella; 4 — гиоидная дуга; 5 — goniale; 6 — сочленовная кость; 7 — венечный отросток (processus coronoideus); 8 — зубная кость (dentale); 9 — подугловая кость (subangulare).
II: 1 — молоточек (malleus) — проксимальная часть меккелева хряща; 2 — наковальня (incus), развившаяся из квадратной кости; 3 — стремечко (stapes), соответствующее столбику птиц и рептилий; 4 — гиоидная дуга (hyoideum); 5 — processus fulianus (goniale); 6 — барабанная кость (tympanicum), соответствующая угловой кости (angulare); 7 — часть меккелева хряща; 8 — зубная кость (dentale).
вперед
в оглавление