в оглавление
назад
Репродуктивная система
Органы размножения млекопитающих устроены сложнее, чем у других амниот. К ним относят половые железы, или гонады (gonads), половые протоки и различные дополнительные органы.
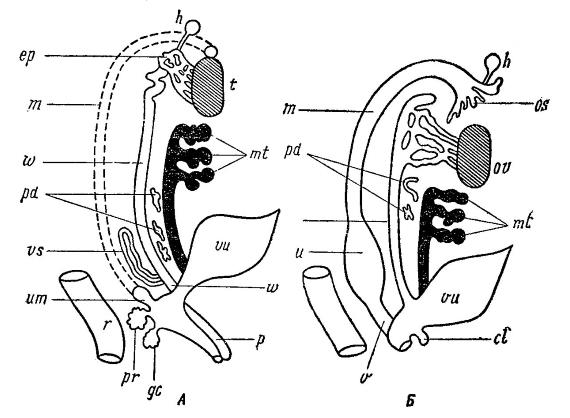
Схема мочеполового аппарата самца (А) и самки (Б) млекопитающих: h — стебельчатая гидатида; m — мюллеров канал (ductus Mulleri); w — семяпровод (ductus deferens); os — воронка яйцевода; ер — придаток семенника (epididymis); t — семенник (testis); ov — яичник (ovarium); mt — почка (metanephros); pd — остаток первичной почки; u — матка (uterus); v — влагалище (vagina); vs — пузыревидная железа (glandula vesicularis); um — мужская матка (uterus masculinus); vu — мочевой пузырь (vesica urinaria); pr — предстательная железа (gl. prostata); gc — куперова железа (gl. couperi); cl — клитор (clitoris); p — копулятивный орган (penis); r — прямая кишка (rectum).
Половые органы самцов
Самцам половыми железами служат парные семенники (testis, или testiculus) — компактные эллипсоидные тела, в которых развиваются мужские половые клетки; в то же время они играют роль желез внутренней секреции. Семенники подвержены сезонным изменениям, но обычно их структура в периоды размножения и покоя различается не так заметно, как у яичников самок.
Семенники закладываются в брюшной полости, где у т. н. крипторхов, подобно рептилиям, остаются пожизненно. Таковы неопущенные абдоминальные семенники однопроходных (Monotremata), части ксенартр (Xenarthra) и большинства афротериев (Afrotheria) — тенреков (Tenrecidae), златокротов (Chrysochloridae), прыгунчиков (Macroscelidea), даманов (Hyracoidea), хоботных (Proboscidea) и сирен (Sirenia). Опускаются из своего первоначального положения, но остаются в брюшной полости или под кожей в паховой области аскротальные семенники некоторых ксенартр (Xenarthra), трубкозубых (Tubulidentata), большинства насекомоядных (Eulipotyphla), части грызунов (Rodentia), рукокрылых (Chiroptera), а также тапиров (Tapiridae), носорогов (Rhinocerotidae), бегемотов (Hippopotamidae), китообразных (Cetacea), панголинов (Pholidota) и ушастых тюленей (Otariidae). У многих млекопитающих семенники в период спаривания опускаются в мошонку (scrotum), а затем втягиваются обратно. Такого рода семенники называются семискротальными, они характерны для некоторых насекомоядных (Eulipotyphla) и летучих мышей (Microchiroptera), а также для зайцеобразных (Lagomorpha) и грызунов (Rodentia). У аплодонтии (Aplodontia rufa) во временную мошонку опускаются абдоминальные семенники. У остальных млекопитающих семенники опускаются в мошонку в раннем возрасте и остаются здесь пожизненно. Такие семенники именуются скротальными и свойственны части грызунов (Rodentia), крыланам (Pteropodidae), приматам (Primates), лошадям (Equidae), большинству хищных (Carnivora) и парнопалым (Artiodactyla). По-видимому, отдельное состояние представляет собой постоянная мошонка сумчатых (Metatheria), расположенная у них впереди пениса.
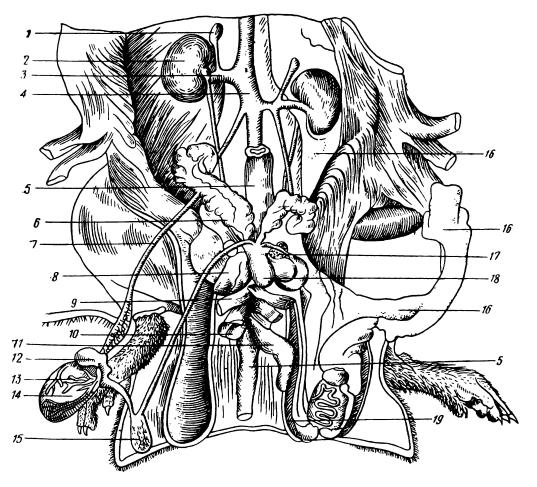
Мочеполовые органы самца серой крысы (Rattus norvegicus): 1 — надпочечная железа; 2 — правая почка; 3 — почечная артерия; 4 — дорсальная аорта; 5 — прямая кишка (rectum); 6 — семенной мешок (vesicula seminalis); 7 — arteria spermatica interna; 8 — простата (gl. рrostate); 9 — musculus rectus abdominis; 10 — museums bulboсavernosus; 11 — половой член (penis); 12 — caput epididymis; 13 — артерии семенника; 14 — семенник; 15 — cauda epididymis; 16 — жир; 17 — железка выносящего протока семени; 18 — мочевой пузырь; 19 — вскрытая мошонка (scrotum). На правой стороне жир удален.
Мошонка представляет собой кожный мешок с парой карманов — влагалищных отростков (processus vaginales), которые выпячиваются наружу на брюшной стороне тела непосредственно перед тазом. Изнутри они выстланы эпителием и образуют особый отдел брюшной полости. Каждый семенник опускается в соответствующий карман мошонки; за ним, естественно, следуют семяпровод и кровеносные сосуды; с дном кармана его связывает брыжейка, называемая направляющей связкой (gubernaculums). В семискротальных семенниках карман мошонки сохраняет проход в брюшную полость — паховый канал (canalis inguinalis), и между сезонами размножения находящаяся в стенке этого кармана мышца (musculus cremaster) может втягивать из него семенник внутрь тела. В скротальных семенниках паховый канал зарастает, и карман мошонки отгораживается от тазовой полости складкой ткани. Прорыв этой складки — слабого места в стенке тазовой полости — может приводить к явлению, называемому у человека паховой грыжей. Чаще всего мошонка помещается позади копулятивного органа, и лишь у сумчатых, зайцеобразных и некоторых насекомоядных располагается кпереди от него. Кроме того, мошонка зайцеобразных примечательна тем, что каждый семенник располагается как бы отдельно, а не в общем мешке и может свободно перемещаться в брюшную полость через широко открытые паховые кольца независимо от сезона размножения.
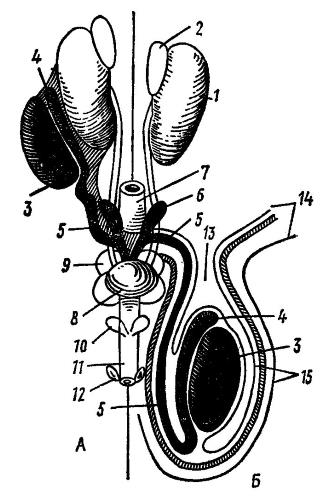
Схема половых органов самцов млекопитающих: А — семенники расположены в брюшной полости; Б — семенники в мошонке; 1 — почка; 2 — надпочечник; 3 — семенник; 4 — придаток семенника; 5 — семяпровод; 6 — семенной пузырек; 7 — прямая кишка; 8 — мочевой пузырь; 9 — предстательная железа; 10 — бульбоуретральные железы; 11 — половой член; 12 — препуциальные железы; 13 — паховый канал; 14 — стенка живота; 15 — стенка мошонки.
Эксперименты и изучение аномалий вскрыли вероятные причины опускания семенников и образования мошонки. Наиболее распространенным является предположение о поддержании комфортной температуры для сперматогенеза и хранения спермы до момента оплодотворения. Та температура, что поддерживается внутри тела млекопитающих, обычно для этого слишком высока, а в выступающих наружу карманах мошонки она на несколько градусов ниже. Однако не исключено, что избегание высоких температур было вторичным, а мошонка развилась вследствие гандикапа или конкуренции спермы. При этом семенники стали очень крупными и появилась необходимость размещать их вне брюшной полости. В таком положении появились адаптации к сперматогенезу при температуре, немного более низкой, чем температура тела. Эти адаптации оказались необратимыми, и при возврате к абдоминальному положению семенников требуется их охлаждение. Существует также предположение, что мошонка появилась в связи с активным передвижением. Дело в том, что при галопировании и прыжках сгибание и разгибание поясничного отдела приводит к периодическому изменению давления в брюшной полости, нарушению оттока крови из брюшных вен и изменению давления внутри семенников, что может негативно сказываться на сперматогенезе. Действительно, многие негалопирующие млекопитающие не имеют мошонки (семенники при этом могут быть опущенными или неопущенными).
Филогенетические исследования дают несколько противоречивые результаты относительно того, сколько раз развивались опускание семенников и мошонка у млекопитающих. По одной из гипотез, мошонка развилась только один раз, и предковым для млекопитающих было наличие мошонки, а её утрата у некоторых современных групп является прогрессивным признаком. Согласно другому предположению, предковым состоянием было отсутствие мошонки и расположенные в полости тела семенники, а сама мошонка появлялась, по меньшей мере, два раза: у сумчатых (Metatheria) и у бореоэвтериев (Boreoeutheria). Этот вариант более правдоподобен, и мошонка у ряда бореоэвтериев утрачена вторично. У афротериев, наоборот, логичнее предположить первичное отсутствие мошонки.
У млекопитающих с аскротальными семенниками для поддержания более низкой их температуры служат лозовидное венозное сплетение и изолирующие жировые подушки. Лозовидное сплетение имеется и у видов с развитой мошонкой. Утрата мошонки у некоторых морских зверей (например, настоящих тюленей и китообразных) сопровождается изменениями в циркуляции крови в области репродуктивных органов. Слоны, даманы, сирены и прочие млекопитающие с абдоминальными семенниками ничего специально не охлаждают, лозовидного сплетения у них нет, а сперматогенез нормально осуществляется при температуре тела. Однако температура тела у этих видов обычно несколько ниже и находится примерно в диапазоне комфортной температуры образования и хранения спермы у других млекопитающих.
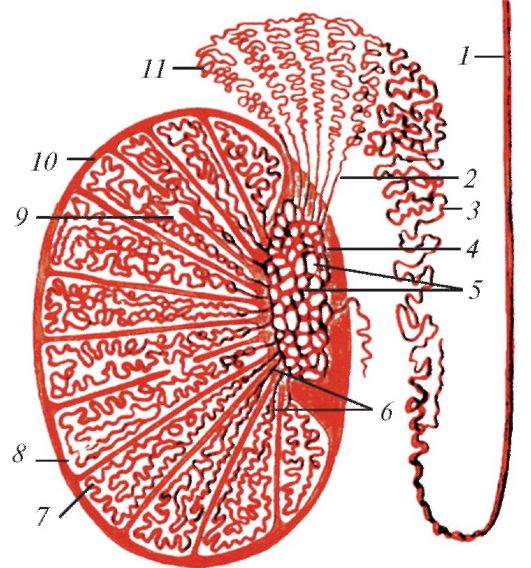
Схема строения семенника и его придатка у человека: 1 — семявыносящий проток; 2 — выносящие канальцы семенника; 3 — проток придатка; 4 — средостение семенника; 5 — сеть семенника; 6 — прямые семенные канальцы; 7 — извитые канальцы; 8 — долька семенника; 9 — сообщения между семенными канальцами соседних долек; 10 — белочная оболочка; 11 — долька придатка семенника.
Семенник пребывает в состоянии тургора. Наружную его часть образует гладкая соединительнотканная белочная оболочка (tunica albuginea), от которой внутрь врастают тяжи, разделяющие семенник на дольки. В дольках находятся извитые семенные канальцы (tubuli seminiferi), сформированные из первичных половых тяжей. Канальцы выстланы эпителием, который в каждый период размножения вырабатывает многочисленные сперматозоиды. Эпителий состоит из нескольких слоев и содержит как половые, так и относительно малочисленные клетки Сертоли, выполняющие вспомогательную и опорную функции. В основании эпителия находятся слабо дифференцированные первичные половые клетки — сперматогонии. Из них после ряда делений формируются зрелые клетки, выходящие на поверхность эпителия в качестве сперматозоидов. В одно и то же время различные извитые канальцы содержат половые клетки на разных стадиях развития. В прослойках между семенными канальцами помещаются крупные интерстициальные клетки Лейдига, вырабатывающие мужской половой гормон тестостерон. С семенниками у некоторых млекопитающих срастаются рудименты начальных отделов мюллеровых каналов (яйцеводов) — гидатиды Морганьи (hydatides Morgagni).
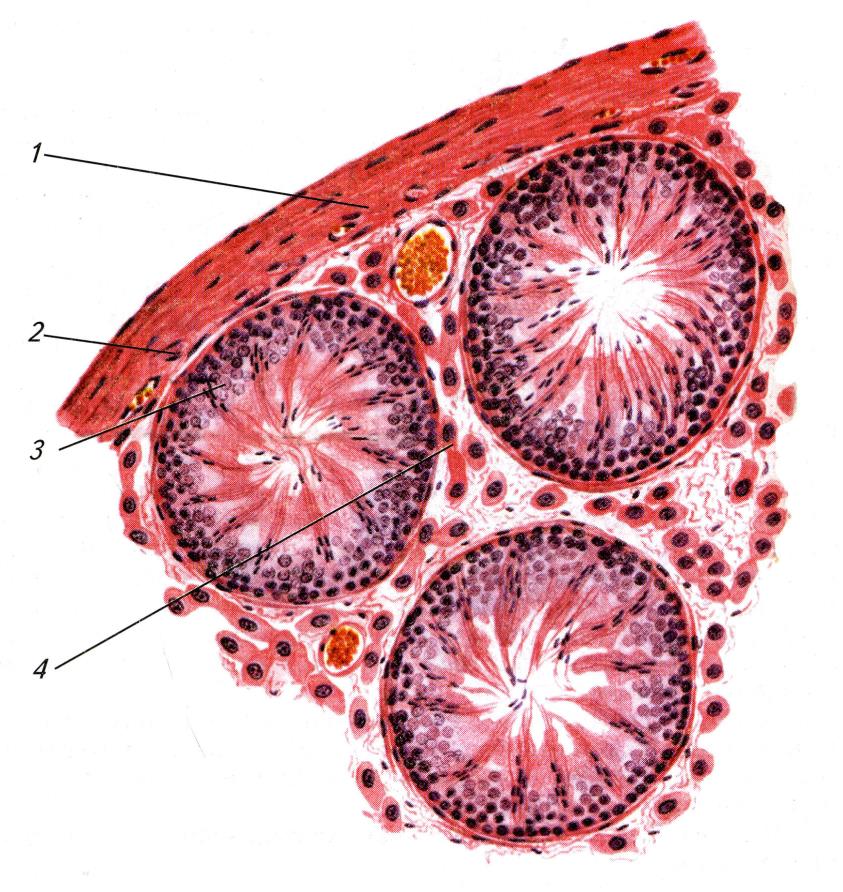
Разрез через семенник серой крысы (Rattus norvegicus): 1 — белочная оболочка; 2 — сосудистая оболочка; 3 — извитые семенные канальцы; 4 — интерстиций.
Концы извитых семенных канальцев соединены друг с другом сетью семенника (rete testis), тоже состоящей из мелких каналов. Сеть семенника переходит в семявыносящие каналы (ductuli efferentes), собирающиеся в большой общий проток, основной отдел которого выходит уже за пределы собственно семенника. Здесь он многократно сворачивается, формируя прилежащее к семеннику и вытянутое по его продольной оси зернистое образование — семенной придаток (epididymis), являющийся остатком переднего отдела туловищной почки, или мезонефроса. Каналы придатков семенников в свою очередь переходят в парные семяпроводы (ductus deferens, или ductus spermaticus). Последние представляют собой производные вольфовых каналов (ducti Wolfii), отводивших от мезонефрических почек мочу (в отличие от самцов, у взрослых самок млекопитающих не сохраняется даже их следов). В области мочевого пузыря семяпроводы с обеих сторон впадают в непарный мочеполовой канал (urethra).
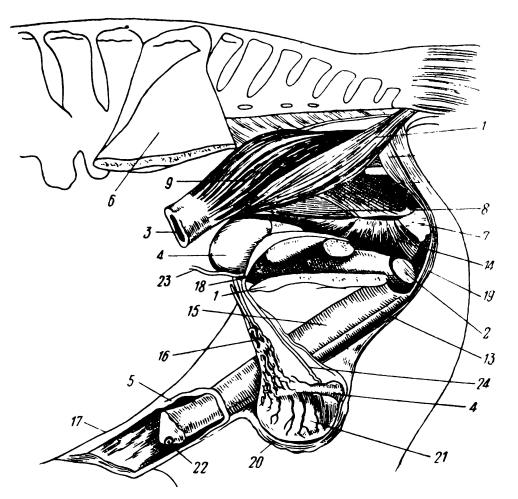
Половые органы самца лошади (Equus ferus): 1 — таз в продольном разрезе; 2 — бульбоуретральные железы; 3 — прямая кишка; 4 — epididymis; 5 — головка (glans penis); 6 — подвздошная кость; 7 — седалищная кость; 8 — порции мускула obturator internus; 9 — мускульные стенки прямой кишки; 10 — m. levator ani; 11 — m. sphincter ani externus; 12 — опускатель анального отверстия; 13 — musculue urenthralis; 14 — musculus obturator internus; 15 — половой член (penis); 26 — plexus pampiniformis; 17 — препуций (praeputium); 18 — простата (prostate); 19 — musculus retractor penis; 20 — мошонка (scrotum); 21 — семенник; 22 — наружное отверстие мочеполового канала; 23 — мочеточник (ureter); 24 — vas deferens.
К мужским половым органам у многих млекопитающих прилегают придаточные железы, главная роль которых — образование жидкой части спермы, или эйякулята. Хорошо развитая предстательная железа, или простата (glandula prostate), лежит у основания полового члена и шейки мочевого пузыря, окружая начальную часть мочеполового канала, куда и открывается. Именно водянистый секрет долек простаты в основном формирует жидкую среду спермы; кроме того, он стимулирует подвижность сперматозоидов. Пузыревидные железы (glandulae vesiculares), иногда не совсем удачно называемые семенными пузырьками (vesiculae seminales), представляют собой парные компактные тела с ребристой поверхностью, которые развиваются на конечной части семяпроводов и отворяются в начало мочеполового канала. Помимо разжижения семени, их густой и клейкий секрет служит для предотвращения обратного вытекания спермы из женских половых путей. У некоторых млекопитающих (части насекомоядных, летучих мышей и грызунов) выделения пузыревидных желез имеют студенистый характер и застывают в половых путях самки, закупоривая их после совокупления (формируют «пробку»). Ниже по бокам мочеполового канала расположена пара луковичных бульбоуретральных, или куперовых, желез (glandulae bulbourethrales). Таким образом, на выходе сперма представляет собой взвесь сперматозоидов в жидком секрете пузыревидных, предстательной в куперовых желез.
У самцов ряда групп (грызунов, копытных, хищных, приматов) в области предстательной железы между двумя семяпроводами сохраняется небольшой тонкостенный пузырек, называемый мужской маткой (uterus masculinus). Этот рудимент образован слитыми конечными отделами мюллеровых каналов и по своему положению соответствует матке (точнее, влагалищу) самок. Мужская матка открывается в основание мочеполового канала и может накапливать половые продукты.
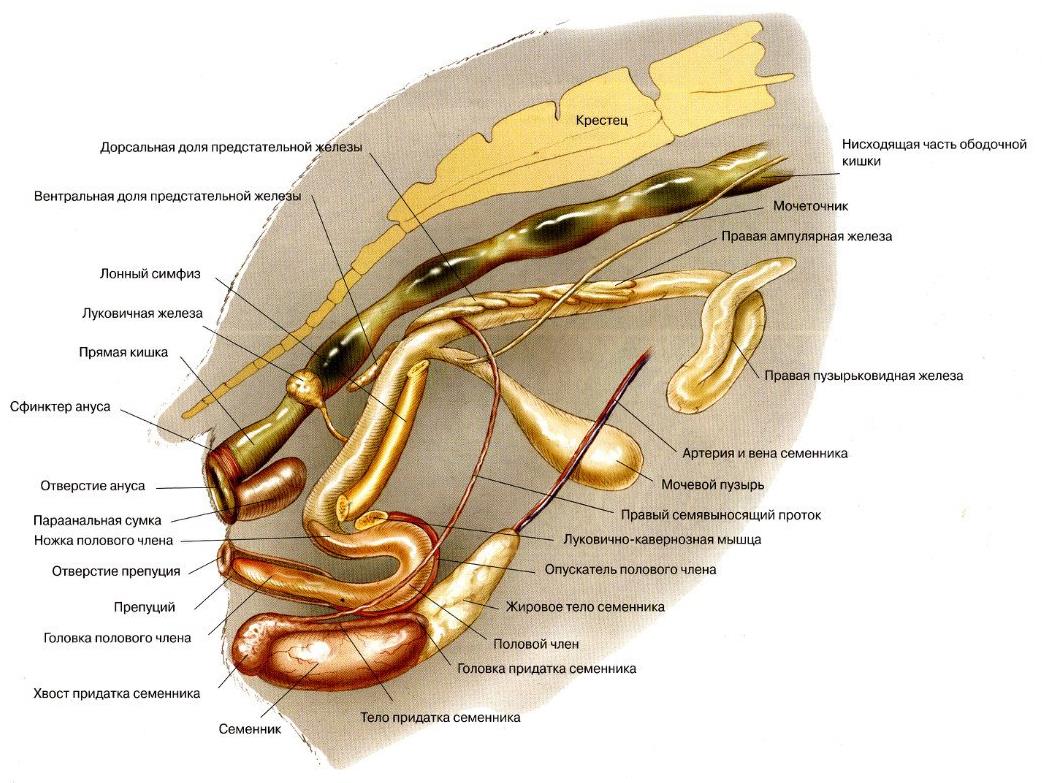
Половые органы самца морской свинки (Cavia porcellus).
Конечная часть мочеполового канала проходит через нижнюю часть полового члена, или пениса (penis), и открывается на его конце. Через это отверстие из организма выводится моча, однако основной функцией данного органа является внесение семени в половые пути самки. Внутри половой член поддерживается пещеристыми телами — прочностенными стержнями из губчатой соединительной ткани, пронизанными густой сетью кровеносных сосудов. Сверху и по бокам лежит пара пещеристых тел полового члена (corpora cavernosa penis), которые своими основаниями крепятся к седалищным костям. Снизу находится пещеристое тело мочеполового канала (corpus cavernosum urethrae), окружающее его подобно трубке. Пещеристые тела разделены продольной перегородкой из волокнистой соединительной ткани. На конце пещеристое тело мочеполового канала переходит в закругленную, иногда коническую мясистую головку (glans penis). Когда половой член находится в спокойном состоянии, головку облекает кожная складка — крайняя плоть, или препуций (preputium), имеющая вид двухслойной трубки. С крайней плотью бывает связана пара небольших плотных препуциальных желез. Они выделяют сильно пахнущий секрет, способствующий встрече особей противоположного пола и созданию состояния полового возбуждения.
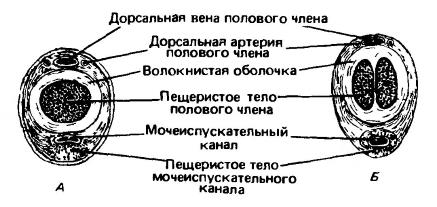
Поперечный разрез полового члена макака-резуса (Macaca mulatta): А — в дистальной части; Б — в проксимальной части.
Вследствие полового возбуждения внутренние полости пещеристых тел и головки под большим давлением наполняются кровью, благодаря чему половой член становится упругим и увеличивается в объеме. Данное состояние — эрекция — является необходимым условием для нормального осуществления полового акта.
В некоторых случаях для дополнительного укрепления полового члена образуется непарная удлиненная кость — бакулюм (baculum). Он обнаруживается у насекомоядных, хищных, части грызунов, некоторых рукокрылых и большинства приматов. Столь широкое распространение данного элемента указывает на его древнее происхождение. Бакулюм формируется из соединительнотканного тяжа, лежащего между пещеристыми телами над мочеиспускательным каналом. Из всех костей он наиболее разнообразен по форме. Так, бакулюм может иметь вид пластинки или стержня различной формы — прямой, кривой или дважды изогнутой; круглой, треугольной, квадратной или плоской в сечении; простой, заостренной, ложковидной, или продырявленной; длинной или короткой. Порой бакулюм достигает больших размеров и значительного веса (например, у моржа (Odobenus rosmarus) он имеет длину до 51,5 см и отвесную ширину у основания до 6,5 см); иногда она почти полностью редуцируется. Основание полового члена обвито двумя парными лентовидными мышцами.

Бакулюмы некоторых современных видов млекопитающих. Сверху — бакулюм моржа (Odobenus rosmarus).
У высших млекопитающих строение полового члена крайне разнообразно. У оленей (Cervidae), полорогих (Bovidae), жирафов (Giraffidae) и некоторых других он несет на конце тонкий отросток, пронизанный мочеполовым каналом. У грызунов (например, у тушканчиков (Dipodidae)) головка бывает покрыта твердыми роговыми шипиками и большими острыми шипами, усиливающими в момент копуляции возбуждение самки. У свиней (Suidae), бегемотов (Hippopotamidae), верблюдов (Camelidae), китообразных и некоторых сумчатых головка рудиментарна или полностью отсутствует. Сам пенис свиней по форме напоминает штопор.
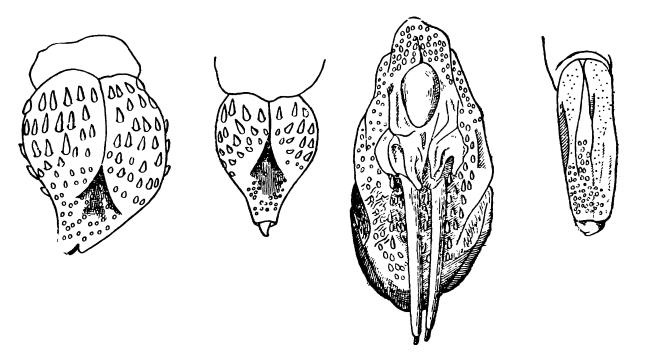
Строение головки полового члена некоторых тушканчиков, слева направо: большой тушканчик (Allactaga jaculus), тушканчик Северцова (Allactaga severtzovi), восточный тушканчик (Jaculus orientalis) (имеются большие шипы), египетский тушканчик (Jaculus jaculus).
У большинства сумчатых половой член раздвоен, соответственно двойному влагалищу самки. В таких случаях мочеполовое отверстие продолжается уретральными желобками на внутренней поверхности каждой половины головки. Кроме того, у ряда сумчатых (в частности, у некоторых хищных форм) снизу на члене развивается дополнительное эректильное тело. Примечательно, что половой член плацентарных также закладывается как парный орган, но в дальнейшем оба его зачатка срастаются.
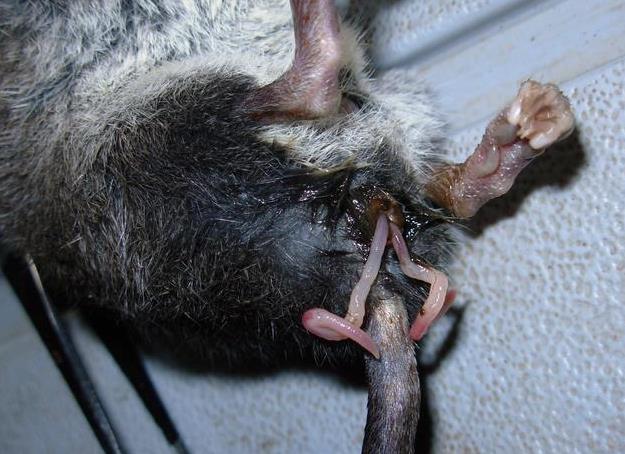
Эрегированный половой член серобрюхого ценолеста (Caenolestes caniventer).
Совершенно особое строение имеет половой член однопроходных, который в некоторых отношениях еще находится на переходной от рептилий стадии. Он расположен внутри клоаки и в спокойном состоянии втянут своим основанием в карманообразный выступ ее вентральной стенки непосредственно под мочеполовым синусом (sinus urogenitalis). По своей структуре этот член напоминает таковой у крокодилов и страусов, однако его боковые стенки замкнулись и образовали цельный канал. Данный канал не имеет прямого сообщения с мочеполовым синусом, но вследствие замыкания отверстия, ведущего из синуса в клоаку, такое сообщение временно устанавливается, и сперма при совокуплении течет не в клоаку, а через половой член. Таким образом, он пропускает только семя, моча выходит в клоаку мимо него. Форма члена также весьма своеобразна: у ехидн (Tachyglossidae) его головка разделяется на 2 лопасти, у утконосов (Ornithorhynchidae) — на 4. Во время полового акта используется только одна половина головки, в результате чего происходит односторонняя эйякуляция.
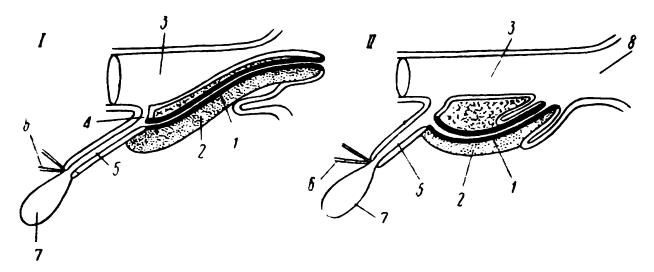
Схема полового члена однопроходных в состоянии эрекции (I) и в спокойном втянутом положении (II): 1 — пещеристая ткань; 2 — фиброзная ткань; 3 — клоака; 4 — мочевой канал; 5 — мочеполовой канал; 6 — мочеточники; 7 — мочевой пузырь; 8 — отверстие клоаки.
Отличия в строении половых органов часто обусловливают биологическую изоляцию между представителями различных таксонов (т. е. невозможность их совокупления). В то же время, они настолько характерны, что служат прекрасными признаками видовой и родовой дифференцировки.
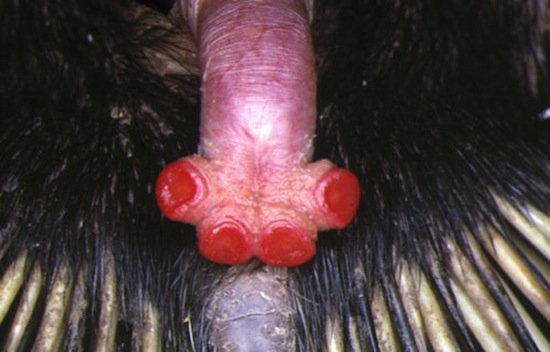
Эрегированный половой член австралийской ехидны (Tachyglossus aculeatus).
Половые органы самок
Половые железы самок представлены парными яичниками (ovaria) — неправильными телами с крупнозернистой поверхностью, подвешенными на широкой брыжейке к спинной стенке брюшной полости невдалеке позади почек. Размер яичников млекопитающих заметно меньше, чем у остальных позвоночных. Снаружи они покрыты зачатковым эпителием, который врастает в виде тяжей внутрь соединительнотканной основы — стромы, с расположенными в ней крупными интерстициальными клетками. К зачатковому слою примыкает фолликулярная зона, где в особых клеточных комплексах — фолликулах (folliculi ovarici) — развиваются половые клетки. Здесь же находятся крупные интерстициальные клетки, продуцирующие половой гормон эстроген. Глубже лежит сосудистая зона с кровеносными сосудами. Примечательно, что у утконоса (Ornithorhynchus anatinus), как и у большинства птиц, созревает только левый яичник.
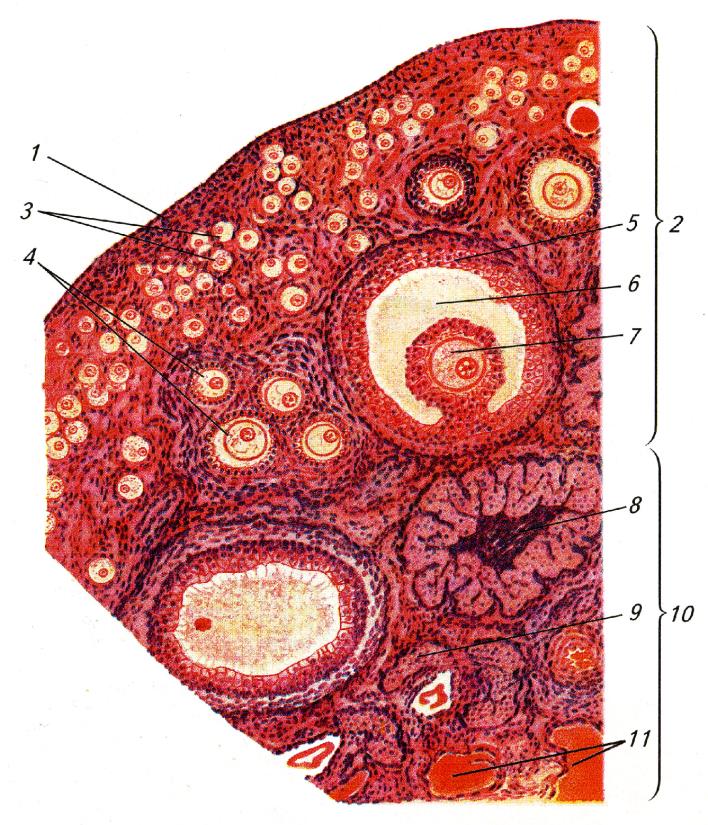
Разрез через яичник млекопитающего: 1 — зачатковый эпителий; 2 — корковое вещество; 3 — первичные фолликулы; 4 — растущие фолликулы; 5 — пузырчатый фолликул (графов пузырек); 6 — полость пузырчатого фолликула, заполненная жидкостью; 7 — яйцеклетка; 8 — желтое тело; 9 — атретические тела; 10 — мозговое вещество; 11 — соединительная ткань и кровеносные сосуды.
Остальные отделы женской половой системы, служащие в основном для выделения созревших яйцеклеток и вынашивания зародышей, представляют собой производные парных яйцеводов (oviducti), соответствующих мюллеровым каналам (ducti Mulleri). В своей начальной части это собственно яйцеводы, или фаллопиевы трубы (tubae uterinae) — тонкие извитые органы, выстланные мерцательным эпителием. Каждая труба начинается бахромчатой воронкой, вплотную приближенной к яичнику и окруженной вместе с ним складкой брюшины, образующей практически замкнутую сумку.
Фаллопиевы трубы переходят в матку (uterus) — толстостенное гладкомышечное образование, служащее для вынашивания зародыша. Ее богатый кровеносными сосудами эпителий — эндометрий (endometrium) — может быть относительно тонким в периоды покоя, но сильно утолщается в сезоны репродкутивной активности; ритмикой его роста управляют гормоны, вырабатываемые яичниками. Плацента (placenta), через которую питательные вещества передаются от матери к зародышу, образуются путем соединения эпителия матки с наружными зародышевыми оболочками.
Строение матки отличается значительным разнообразием. У однопроходных два раздельных яйцевода расширяются и переходят в такие же разделенные матки, которые независимо друг от друга и без образования влагалищ открываются в мочеполовой синус, далее переходящий в клоаку. Матка сумчатых также парная. У плацентарных наблюдаются все переходы от парной матки к непарной, с различной степенью слияния правого и левого маточных отделов. В двойной матке (uterus duplex) дистальные концы отделов сильно сближены, но открываются во влагалище самостоятельными отверстиями (некоторые рукокрылые, большинство грызунов, трубкозубые, даманы, хоботные). При двураздельной матке (uterus bipartitus) отделы срастаются дистальными концами и имеют одно общее выводное отверстие, но перегородка между ними сохраняется на всем остальном протяжении (часть рукокрылых, грызунов, хищных и свиные). В одиночной двурогой матке (uterus bicornis) отделы на значительном протяжении образуют единую полость, и лишь в передних верхних углах разделяются на два рога (насекомоядные, некоторые рукокрылые, большинство хищных и копытных, китообразные, мокроносые приматы). Наконец, в одиночной простой матке (uterus simplex) ее отделы полностью сливаются в мешкообразный орган, и парными остаются только верхние отделы яйцеводов — фаллопиевы трубы (ксенартры, некоторые рукокрылые, сухоносые приматы).
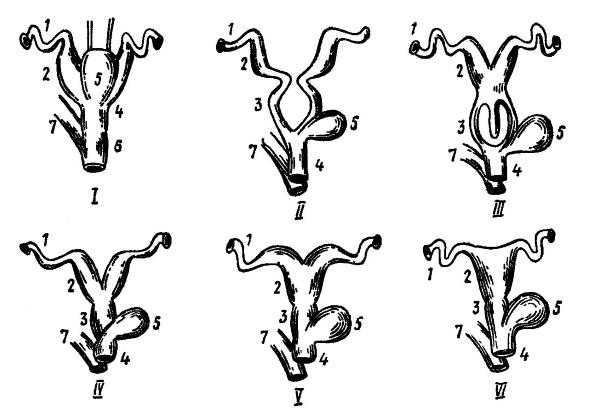
Схема эволюции яйцеводов у млекопитающих: I — однопроходное; II — низшее сумчатое (опоссум); III — высшее сумчатое (кенгуру); IV — плацентарное с двойной маткой; V — плацентарное с двурогой маткой; VI — плацентарное с простой маткой; 1 — фаллопиева труба; 2 — матка; 3 — влагалище; 4 — преддверие влагалища; 5 — мочевой пузырь; 6 — клоака; 7 — прямая кишка.
Конечная часть половых путей, служащая для выведения плода и принятия семени самца при копуляции, носит название влагалища (vagina). У сумчатых нижние участки первичных яйцеводов преобразуются в парные влагалища. Одними концами они открываются в мочеполовой синус, а другими соединяются во влагалищный синус, куда выходят отверстия шеек маток. Иногда влагалищный синус вытягивается, достигает своим нижним концом мочеполового синуса и прирастает к нему, образуя непарный слепой мешок. После первых родов этот мешок прорывается и образуется третий, срединный проход из матки наружу — псевдовлагалищный родовой канал. Обычные влагалища в подобных случаях для рождения детенышей не используются (например, у вомбатов, некоторых кенгуру).
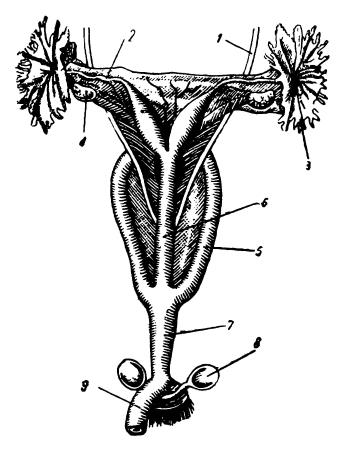
Половые органы рожавшей самки рыже-серого валлаби (Macropus rufogriseus): 1 — мочеточник; 2 — яйцевод; 3 — отверстие яйцевода; 4 — яичник; 5 — влагалища (по одному с каждой стороны); 6 — псевдовлагалищный родовой канал; 7 — мочеполовой синус; 8 — заднепроходные железы; 9 — прямая кишка.
У плацентарных влагалище всегда одно (в эмбриогенезе оно закладывается как парное, но затем его участки сливаются). Вместе с относительно коротким мочеиспускательным каналом оно открывается в гораздо более широкий, чем у самцов, мочеполовой синус — преддверие влагалища (vestibulum), или же отдельно, половым отверстием на поверхности тела. Первый вариант характерен для зайцеобразных, хищных и многих других зверей, второй встречается, например, у мышевидных грызунов.
Преддверие может быть относительно неглубоким (как у приматов) или достигать значительной глубины и выглядеть продолжением собственно влагалища (как у типичных хищных). В стенке преддверия расположен небольшой, выступающий наружу вырост — клитор (clitoris), гомологичный половому члену самцов. Подобно последнему, клитор состоит из пещеристых тел, головки и сильно варьирует по своей структуре. Иногда он содержит косточку, похожую на бакулюм самцов, но меньшую по размерам — баубеллюм (baubellum); она наблюдается в тех же систематических группах. У некоторых насекомоядных, грызунов и мокроносых обезьян в клиторе имеется канал, и тогда мочевой и половой пути оказываются разделенными. С преддверием влагалища связаны парные бартолиновы железы (glandulae vestibulares majores), гомологи бульбоуретральных желез самцов, выделяющие увлажняющий секрет.
Наружу преддверие открывается щелевидным мочеполовым отверстием — вульвой (vulva). С боков вульва ограничена кожными складками — большими половыми губами (labia majorа pudendi); у приматов также имеются более глубоко лежащие малые половые губы (labia minora pudendi). Расположенные здесь кожные железы выделяют пахучий секрет. У сумчатых, грызунов, копытных и приматов вход во влагалище суживается кольцевой перепонкой — девственной плевой (hymen vaginae), разрываемой при первом совокуплении. У молодых самок кротов половое отверстие отсутствует, а у взрослых бывает открыто только в период размножения (вероятно, это должно воспрепятствовать попаданию в половые ходы частиц грунта). После акта совокупления у грызунов, насекомоядных и летучих мышей образуется слизистая пробка, замыкающая вход во влагалище.
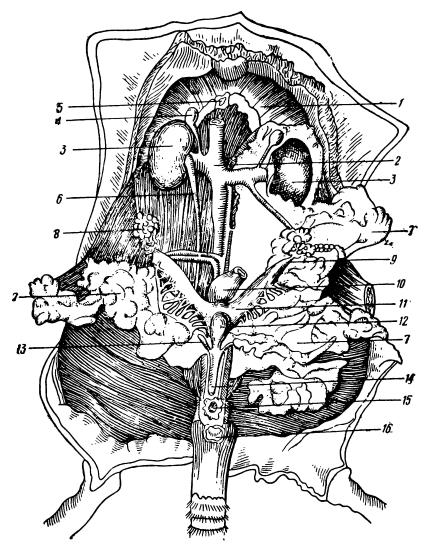
Мочеполовые органы самки серой крысы (Rattus norvegicus): 1 — диафрагма; 2 — дорсальная аорта; 3 — почки (левая прикрыта слоем жира); 4 — надпочечная железа; б — перерванная нижняя полая вена; 6 — мочеточник; 7 — жир; 8 — яичник; 9 — яйцевод; 10 — прямая кишка; 11 — матка; 12 — мочевой пузырь; 13 — мочеточник; 14 — нижний канал мочеточника; 15 — влагалище (vagina); 16 — анальное отверстие (на правой стороне большая часть жира удалена).
На ранних стадиях развития эмбрионов строение наружных половых органов у самцов и самок сходное. Однако в дальнейшем пещеристые тела увеличиваются в диаметре и в длину так, что уретра и половые протоки сливаются в единый канал, расположенный между ними — формируется половой член. У представителей большинства групп полость тела в области промежности выпячивается наружу и формирует мошонку, куда затем мигрируют семенники. У самок по бокам от мочеполового синуса развиваются складки — половые губы, а пещеристые тела образуют клитор. В остальном строение внешних половых органов остается сходным с эмбриональным. Яичники остаются в полости тела. Центральная часть мюллерова канала расширяется, образуя матку. При этом канал делится на собственно яйцевод, матку и влагалище.
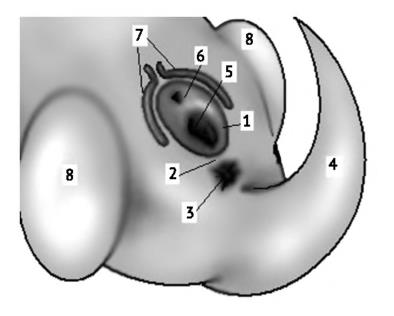
Закладка внешних половых органов млекопитающего: 1 — мочеполовой синус; 2 — промежность; 3 — анус; 4 — хвост; 5 — половое отверстие (протоки гонад); 6 — уретра; 7 — пещеристые тела; 8 — почки задних конечностей.