в оглавление
назад
Выделительная система
Как и у прочих амниот, процесс выделения из крови конечных продуктов метаболизма — азотистых продуктов белкового обмена, солей и воды — у млекопитающих осуществляется главным образом парными метанефрическими почками (metanephros). Номинально это тазовые почки, но расположены они кпереди от таза в поясничной области. Только у молодых однопроходных и сумчатых некоторое время функционируют свойственные анамниям мезонефрические, или туловищные, почки (mesonephros), у высших же млекопитающих они существуют только в эмбриональный период. От лентовидных туловищных почек у взрослых животных остаются лишь незначительные части (paradidymis), расположенные книзу от гонад. Помимо почек, в выделительных процессах у млекопитающих участвуют кишечная трубка, дыхательные органы и кожа.
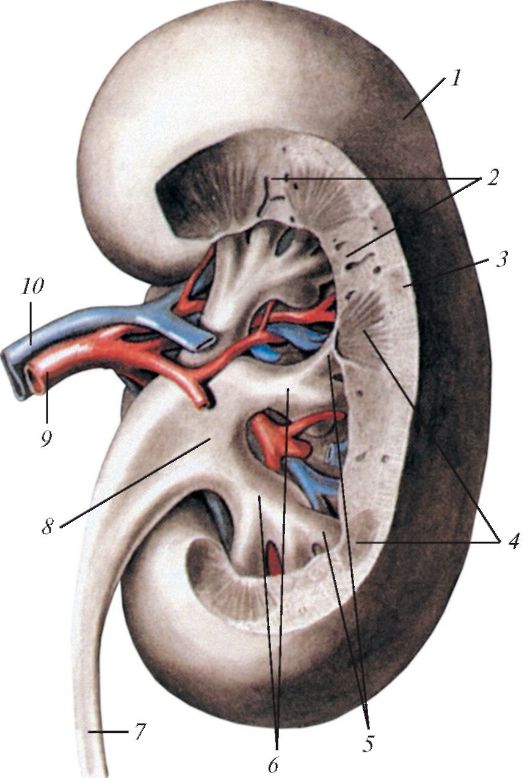
Продольный (фронтальный) разрез почки человека, вид сзади: 1 — капсула почки; 2 — почечные столбы; 3 — корковое вещество; 4 — мозговое вещество (пирамиды); 5 — малые почечные чашки; 6 — большие почечные чашки; 7 — мочеточник; 8 — почечная лоханка; 9 — почечная артерия; 10 — почечная вена.
Почки (renes) у большинства млекопитающих представляют собой компактные тела бобовидной формы с вырезкой на внутренней стороне, плотно прилегающие к спинной стенке брюшной полости по бокам от позвоночника. Поверхность их чаще гладкая, иногда бугорчатая (жвачные (Ruminantia), кошки (Felidae)). У медведей (Ursidae), ластоногих (Pinnipedia), китообразных (Cetacea), сирен (Sirenia), хоботных (Proboscidea) и некоторых других почки поделены перехватами на множество обособленных долек (reniculi), каждая из которых прикреплена к отдельной ветви мочеточника; у некоторых видов китов их число доходит до нескольких тысяч. Адаптивное значение такого разделения пока неясно. Спереди к почкам примыкают железы внутренней секреции — надпочечники (glandulae suprarenales).
Относительные размеры почек находятся в обратной зависимости от величины тела. Они велики у мелких форм и наоборот. Такая зависимость вполне понятна, если учесть, что у близких филогенетически и экологически видов интенсивность обмена веществ будет тем выше, чем меньше размеры тела. Хорошим, хотя и косвенным, подтверждением сказанному служит ясная корреляция между показателями относительной массы почки и сердца — органов, в одинаковой мере ответственных за уровень метаболизма.
На срезе почки млекопитающих обычно четко различимы зернистый наружный корковый (cortex renis) и исчерченный внутренний мозговой (medulla renis) слои. Обе эти области сообщаются с почечными артериями и венами. Корковый слой состоит в основном из почечных телец и извитых отделов канальцев. Мозговой слой построен главным образом из длинных прямых собирательных трубочек и петель Генле; именно они придают этому слою радиально исчерченный вид. Почечное, или мальпигиево, тельце (corpusculum renis) осуществляет первичный фильтрационный процесс. Оно состоит из двух частей — клубочка (glomerulum) и охватывающей его боуменовой капсулы (capsula glomeruli). Клубочек представляет собой маленькое компактное скопление артериальных капилляров, свернутых сложными петлями. Полусферическая боуменова капсула состоит из двух слоев. Внутренний слой, плотно прилегающий к стенкам сосудов клубочка, построен из сложно-звездчатых клеток — подоцитов, между отростками которых имеются узкие щели, так что местами от полости между слоями капсулы кровеносные сосуды отделяет лишь базальная мембрана.
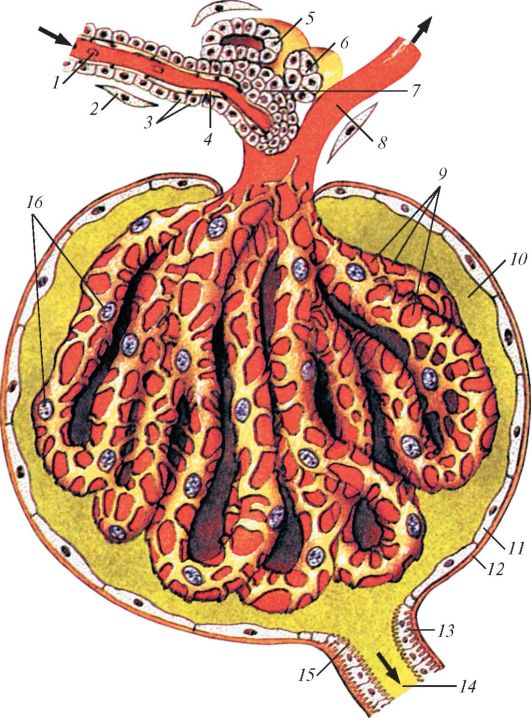
Схема микроскопического строения почечного тельца млекопитающего: 1 — приносящая клубочковая артериола; 2 — адвентициальная клетка; 3 — парагломерулярные клетки; 4 — эндотелиальная клетка; 5 — стенка дистального отдела нефрона; 6 — плотное пятно дистального отдела; 7 — клетки парагломерулярного комплекса; 8 — выносящая клубочковая артериола; 9 — клубочковые кровеносные капилляры; 10 — просвет капсулы клубочка; 11 — клетка наружной части капсулы клубочка; 12 — базальная мембрана наружной части капсулы клубочка; 13 — базальная исчерченность; 14 — проксимальная часть канальца нефрона; 15 — щеточная каемка; 16 — подоциты.
Из полости между слоями боуменовой капсулы выходит извитой каналец (tubulus contortus). Начинаясь извитым участком (проксимальным извитым канальцем), он переходит в петлю Генле. Эта петля представляет собой узкий и в основном прямолинейный тонкостенный канал, сложенный U-образно; он направлен в мозговой слой почки. Восходящая ветвь петли Генле в своей верхней половине расширена и более толстостенна. Она возвращается в корковый слой почки и здесь переходит в дистальный извитой каналец. Вместе со многими другими канальцами он впадает в прямую собирательную трубочку, пронизывающую мозговой слой параллельно петлям Генле. Проксимальный извитой каналец, петля Генле и дистальный извитой каналец густо оплетены сетью венозных капилляров. Для млекопитающих характерно веерообразное расположение собирательных трубочек (в отличие от гребенчатого у рептилий и птиц), которые сходятся в группы — пирамиды, или дольки почек (pyramida renalis).
Своими вершинами, т. н. почечными сосочками (papilla renalis), пирамиды обращены в медиальную, вогнутую сторону почки, где собирательные трубочки открываются в воронкообразный резервуар — почечную лоханку (pelvis renalis). Ближе к пирамидам лоханка нередко подразделяется на мелкие углубления — почечные чашечки (calices). За почечными воротами (hilus renalis) лоханка суживается и переходит в мочеточник (ureter). У огромного большинства млекопитающих парные мочеточники впадают в накопительный резервуар — мочевой пузырь (vesica urinaria). Он представляет собой грушевидный мешок с сильно растяжимыми, но крепкими стенками, содержащими толстый слой гладкой мускулатуры. Когда мочевой пузырь пуст, его внутренняя выстилка выглядит как многослойный эпителий; когда пузырь наполнен и этот эпителий растянут, он утоньшается до одного или двух слоев плоских клеток. Мочевой пузырь служит гигиеническим приспособлением. Выводной проток пузыря открывается в шейке вблизи отверстий мочеточников. У самцов он впадает в мочеполовой канал (urethra), проходящий внутри копулятивного органа и открывающийся на его конце. Короткий мочеиспускательный канал самки открывается в преддверие влагалища, откуда моча выводится наружу; в ряде случаев (например, у самок мышевидных грызунов) канал открывается отдельным мочевым отверстием. У однопроходных мочеточники открываются на сосочке в мочеполовой синус в верхнем отделе клоаки, откуда моча поступает в лежащее напротив отверстие мочевого пузыря.
Почечное тельце и его извитой каналец в совокупности образуют структурно-функциональную единицу почки — нефрон. У нефронов две главные функции: стабилизация среды организма и удаление отходов. Хотя у млекопитающих количество нефронов меньше, чем у птиц такого же размера, оно все же велико. Так, у мыши в обеих почках их насчитывается около 20 тыс., у кролика — 300 тыс., у человека — 2 млн. Для сравнения укажем, что в почках у курицы примерно 200 тыс., у лягушки 2 тыс., а у тритона — всего 500 нефронов.
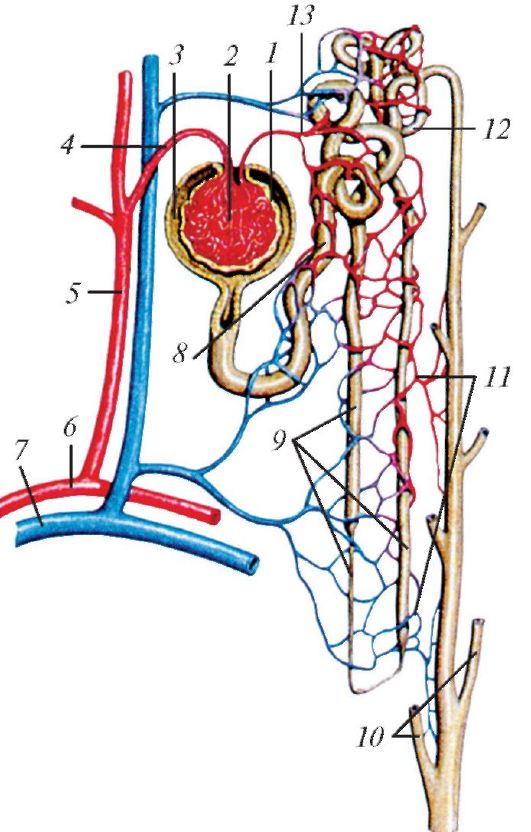
Схема строения и кровоснабжения нефрона млекопитающего: 1 — почечное тельце; 2 — клубочек почечного тельца (капилляры); 3 — капсула клубочка (боуменова капсула); 4 — приносящая клубочковая артериола; 5 — междольковая артерия; 6 — дуговая артерия; 7 — дуговая вена; 8 — проксимальная часть канальца нефрона; 9 — петля нефрона; 10 — сосочковые протоки; 11 — околоканальцевые капилляры; 12 — дистальная часть канальца нефрона; 13 — выносящая клубочковая артериола.
Результат функционирования нефрона — образование мочи, извлекаемой из связанных с ним кровеносных сосудов и предназначенной для выделения (а, следовательно, и регулировании химического состава и водного баланса крови). При этом в нефроне протекают два различных процесса: один связан с почечным тельцем, а другой — с канальцами. По строению тельце похоже на относительно простое фильтрующее приспособление: сквозь мембраны, отделяющие клубочек от полости боуменовой капсулы, под давлением проходит отфильтрованная плазма крови. Особую роль для эффективности фильтрации играет создаваемый энергией сердца перепад давления в артериолах клубочка, дополнительно поддерживаемый сужением выходного сосуда. Определенная доля этого перепада тратится на преодоление коллоидного осмотического давления белков. Анализ отобранной из боуменовой капсулы жидкости показывает присутствие в ней многих характерных для крови веществ, причем в таких же количественных соотношениях. Форменные элементы и крупные молекулы типа большинства белков сквозь этот фильтр не проникают, но остальные соединения, включая не только отходы, но и такие полезные для организма вещества, как сахара, соли, витамины, аминокислоты, — проходят через него легко. Кроме того, очень велико количество просачивающейся воды. Образовавшийся фильтрат условно называют первичной мочой. Подсчитано, что, если бы у человека вся первичная моча удалялась из организма, он выделял бы за сутки порядка 180 л жидкости.
Таким образом, если бы в почках шла только клубочковая фильтрация, организм вскоре совсем лишился бы воды. Кроме того, этот процесс не мог бы существенно менять количественные соотношения компонентов в жидкости тела: конечные продукты обмена и вредные или находящиеся в избытке соединения оставались бы в организме в прежних концентрациях, а ценные питательные вещества терялись бы. Эту проблему разрешает работа канальцев. Их выстилает эпителий, меняющий свой характер от одного отдела канальцев к другому. Основная функция его клеток — реабсорбция, т. е. обратное всасывание большей части фильтрата. Прежде всего, всасывается много воды; кроме того, по мере прохождения жидкости из нее извлекаются ценные вещества. Очистка первичной мочи от полезных примесей производится двумя путями — диффузией через стенки канальцев, подчиняющейся обычным физическим законам (разность концентрации в канальцах и в окружающей ткани почки) и путем активной реабсорбции с участием ферментов, связанных с мембранами клеток стенок канальцев.
В проксимальном извитом канальце идет реабсорбция глюкозы, витаминов, аминокислот, гидрокарбонатов, а также ионов хлора, калия и частично натрия. Параллельно изъятию полезных веществ, здесь происходит дальнейшее выделение в мочу отходов. Оставшаяся жидкость — в основном разбавленный раствор солей и мочевины — направляется по петле Генле в мозговой слой почки, в область пирамиды, где поддерживается высокое осмотическое давление тканевой жидкости (вне канальцев) в силу высокой концентрации в ней солей и мочевины. Благодаря хорошей проницаемости стенок нисходящей части петли Генле для воды она интенсивно выходит из канальцевой жидкости вплоть до достижения осмотического равновесия с окружающей жидкостью, отчего концентрация мочи повышается у разных млекопитающих в 5-16 раз. Вместе с тем, условий для реабсорбции ионов и других веществ в нисходящей части петли почти нет. Наоборот, стенки восходящей части петли Генле легко проницаемы для солей, но не для воды и лишь слабо — для мочевины. Соли пассивно покидают мочу по мере движения по канальцу в область менее концентрированной тканевой жидкости, а в его верхней, толстостенной части хлорид натрия активно выводится наружу с затратой значительной энергии. Это наиболее энергоемкое действие в почечной деятельности; возможно, именно им объясняется редукция воротной системы, благодаря которой почка получает только артериальную, богатую кислородом кровь. Активное удаление ионов натрия и хлора из мочи как раз и запускает процесс их накопления в тканевой жидкости и, в конечном счете, способствует эффективной реабсорбции воды, приводящей к выделению концентрированной мочи.
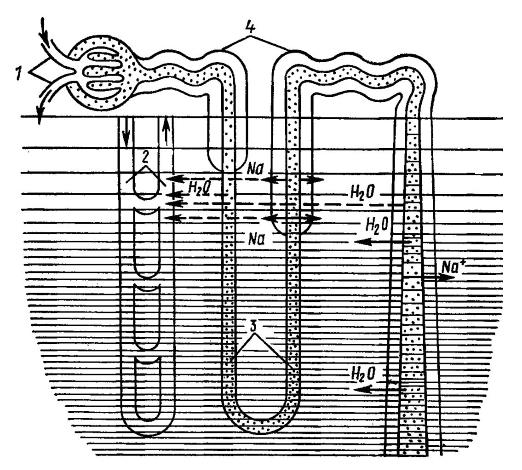
Противоточная система в петле Генле: 1 — приносящий и выносящий сосуды клубочка; 2 — кровеносные сосуды ткани почки; 3 — петля Генле; 4 — извитые отделы почечного канальца. Штриховкой показана концентрация ионов натрия в тканевой жидкости почки, точками — то же, в первичной моче (фильтрате клубочка). Сплошные стрелки — активная реабсорбция ионов натрия, пунктирные — пассивная реабсорбция. Вода (показана стрелками) пассивно переходит в межтканевую жидкость почки.
После петли Генле вернувшаяся в область коркового слоя моча переходит в дистальный извитой каналец. Он играет ключевую роль в регуляции концентрации ионов в организме за счет изменения количеств секретируемых в мочу ионов калия и реабсорбируемых хлорида натрия. В результате жидкость здесь имеет заметную концентрацию мочевины, но не соли. Проходя по собирательной трубочке сквозь пирамиду с ее гипертонической средой, моча опять интенсивно отдает воду и приобретает высокую концентрацию мочевины. Нижняя часть трубочки высокопроницаема для мочевины, которая частично покидает мочу и пассивно, по градиенту концентрации переходит в тканевую жидкость; здесь ее накопление дополнительно повышает осмотическое давление. Вода, попадающая из собирательной трубочки в эту жидкость и непрерывно разбавляющая ее, удаляется кровью, которая прошла через клубочек и по особым прямым сосудам сопровождает петлю Генле. Кроме того, эпителий собирательной трубочки выделяет в фильтрат ионы аммония и водорода, регулируя тем самым водородный показатель организма; в результате азот находится в моче млекопитающих не только в виде мочевины, но и в виде ионов аммония.
Всасывние воды в почечных канальцах опосредованно белками аквапоринами, образующими поры в мембранах их эпителия. Уменьшая или увеличивая число аквапоринов, можно изменять осмолярность мочи и ее количество. Осмолярность мочи также можно регулировать, изменяя осмолярность тканевой жидкости и тем самым изменяя параметры реабсорбции ионов. Эти механизмы дают возможность получать концентрированную мочу в условиях недостатка влаги и разбавленную — при избытке воды. Вообще, количество мочи и ее осмолярность находятся под контролем ряда гормонов, в частности, вазопрессина, ренин-ангиотензин-альдостероновой системы и предсердного натрийуретического пептида.
Таким образом, прошедшая весь цикл моча сильно отличается от исходного фильтрата. Прежде всего, очень сильно уменьшается в объеме ее жидкая составляющая. Например, у человека мочевого пузыря достигает едва ли 1 % ее первоначального количества, отфильтрованного из клубочков. Кроме того, значительно изменяется состав мочи, поскольку питательные вещества типа глюкозы обычно извлекаются, количественные же соотношения других веществ становятся в корне иными. Эту подготовленную к выведению из организма мочу именуют конечной. Ее нормальный желтоватый цвет обусловлен продуктом распада гемоглобина — урохромом.
Как можно видеть, в процессе концентрирования первичной мочи не происходит ни активного переноса воды через стенку канальца, ни секреции мочевины. Эти вещества диффундируют пассивно, в соответствии с осмотическим градиентом. Активно происходит только перенос хлорида натрия в конечном участке восходящей части петли Генле, который не позволяет соли покинуть мозговое вещество почки, фактически запирая ее в этой зоне. Нисходящая часть петли, стенки которой непроницаемы для электролитов, без потерь транспортирует соли к вершине пирамиды (с нарастанием концентрации, поскольку вода свободно диффундирует наружу) и лишь на конце петли начинает выпускать ее в окружающую тканевую жидкость. Таким образом, длинная петля Генле выполняет роль противоточной умножительной системы, создающей градиент концентрации солей в тканевой жидкости мозгового слоя почки. Но важна и задача поддержания этого градиента, компенсации непрерывного разбавления солей извлекаемой из первичной мочи водой. Эту воду уносят из пирамиды прямые сосуды — петли артериол, куда кровь поступает после клубочков. Они образуют вторую противоточную систему.
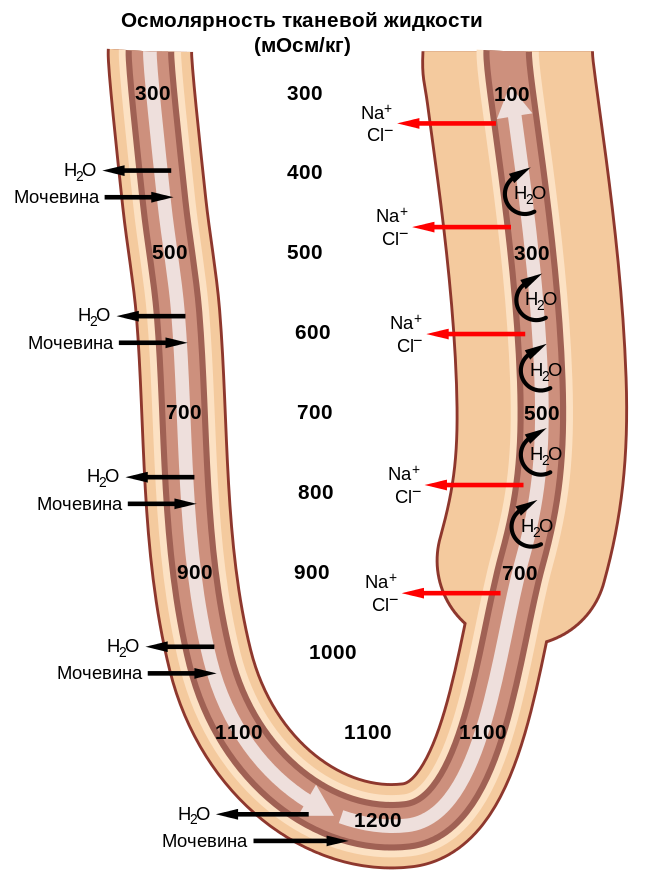
Реабсорбция в петле Генле.
Среди позвоночных животных только птицы и млекопитающие обладают способностью транспортировать соли сквозь стенку почечного канальца против градиента осмотического давления. У птиц данная способность выражена слабее, поскольку их петли Генле значительно короче и присущи не всем нефронам. В результате осмотическое давление мочи у них может лишь вдвое превышать соответствующий показатель для плазмы крови. У млекопитающих выделяют три типа нефронов: суперфициальные, расположенные поверхностные в коре и имеющие короткую петлю Генле (примерно 15 % всех нефронов); интракортикальные, играющие основную роль в ультрафильтрации мочи (70 %); юкстамедуллярные, выполняющие основную роль в процессах концентрирования и разведения мочи (15 %). Длина петли Генле у юкстамедуллярных нефронов в значительной мере определяется условиями обитания того или иного вида. Так, у нефронов млекопитающих, живущих в засушливых районах, петля Генле длинная, глубоко уходящая в мозговое вещество почки. Благодаря ее значительной длине поддерживается высокий осмотический градиент, в результате чего образуется высококонцентрированная моча, осмотическое давление которой может быть повышено по сравнению с плазмой крови в 25 раз. Напротив, у нефронов водных млекопитающих, не испытывающих проблем с дегидратацией, петля Генле значительно короче, что сильно ограничивает их возможности по концентрированию мочи. Наконец, наземные млекопитающие, не испытывающие регулярного дефицита влаги, имеют нефроны со средней длиной петли Генле, которые выделяют мочу промежуточной степени концентрирования.
Итак, клеткам тела для жизни необходима соответствующая среда, самая важная характеристика которой — постоянное присутствие в окружающей их тканевой жидкости определенного количества и соотношения некоторых простых солей в растворенном виде. Их ионы легко проходят через стенки капилляров, поэтому их количественные соотношения в крови и тканевой жидкости одинаковы. Поддержание необходимой концентрации солей во всех жидкостей тела требует поддержания равновесия между их поступлением в организм (главным образом через кишечник) и выделением. Главную роль в устранении лишней воды, которая могла бы слишком сильно разбавить тканевую жидкость, а также избытка солей играют нефроны. Помимо почек, некоторую роль в водном балансе играет толстый кишечник, стенки которого всасывают воду из непереваренных пищевых остатков перед формированием из них каловых масс. Часть воды в виде пара покидает организм через дыхательные пути. Кроме того, некоторое количество воды и солей выводится через кожные потовые железы.
Как уже говорилось, помимо регуляции содержания воды и солей нефроны выполняют другую важную функцию, удаляя из организма отходы и вредные вещества. При всасывании переваренной пищи в жидкости тела могут попадать соединения бесполезные или даже ядовитые для клеток; обычно почки их эффективно выводят. Но в первую очередь необходимо устранять конечные продукты обмена, особенно белкового. При метаболизме из любой органической пищи образуется двуокись углерода и лишняя вода, легко удаляемые. Однако возникают и другие вещества. В основном это простые азотистые соединения, среди которых преобладает высокотоксичный аммиак. Примерно с той же скоростью, с какой он покидает клетки и появляется в крови, почти весь аммиак преобразуется в печени в более безвредное соединение — карбамид, или мочевину. Другой метаболический продукт — мочевая кислота — образуется в результате распада пуринов. Незначительное количество (не более 3 %) азотистых соединений может удаляться потовыми железами, но в основном избыточного азота удаляется почками млекопитающих.
Большую часть выделяемых конечных продуктов белкового обмена у млекопитающих, как и у рыб и амфибий, и в отличие от рептилий и птиц, составляет не мочевая кислота, а мочевина. Так, если у птиц долевое значение мочевой кислоты равно 63-80 % и на долю мочевины приходится 1-10 %, то у млекопитающих соотношение обратное: мочевина составляет 68-91 %, а мочевая кислота — 0,1-8 %. Мочевина гораздо токсичнее мочевой кислоты, а выведение такой мочи влечет большой расход воды. Судя по всему, этот тип обмена развился у млекопитающих в связи с наличием плаценты, через которую зародыш может получать воду практически в неограниченном количестве из кровяного русла материнского организма. С другой стороны, через плаценту, точнее систему ее кровеносных сосудов, из развивающегося эмбриона могут неограниченно выводиться токсичные продукты белкового метаболизма.

Черный носорог (Diceros bicornis) метит свой участок обитания, разбрызгивая мочу.
Обеспеченные водой обитатели увлажненных биотопов имеют щедрый водный обмен. Они выделяют большое количество жидкой мочи, а основными механизмами, защищающими их организм от перегрева, является потение и испарение путем тепловой одышки. Однако для млекопитающих, существующих в аридных условиях, количество доступной влаги строго лимитировано. Поэтому в результате длительной эволюции у них развился сложный физиологический комплекс, помогающий экономить воду. Обитатели полупустынных биотопов большую часть воды получают, поедая сочные части суккулентных растений. Их кожно-легочные потери воды минимальны. Так, при температуре 20 °С у относительно влаголюбивого вида — тамарисковой песчанки (Meriones tamariscinus) — они достигают 170 куб. см, а у сухолюбивой большой песчанки (Rhombomys opimus) — только 50 куб. см на 1 кг массы за 1 ч.
Настоящие пустынные млекопитающие способны поедать почти сухие корма и обходиться без питья, довольствуясь той водой, которую они получают путем метаболического расщепления органических веществ, поступающих с пищей. Таковы, к примеру, тушканчики (Dipodidae). Их почки вырабатывают мочу, в 3-4 раза более концентрированную, чем у человека, а кал обезвожен в большей мере, чем у каких-либо других млекопитающих. Кроме того, они совсем не потеют. Тушканчики гибнут при температуре около 34 °C, поэтому старательно избегают перегрева: день проводят в норах, а активную деятельность начинают после наступления сумерек.
Пустынные копытные хорошо переносят жажду и могут обходиться без воды много дней. Лучше других к сухости климата приспособились верблюды (Camelus). Большинство млекопитающих погибает уже при потере 20 % воды, находящейся в их организме. В отличие от них, верблюды способны выдерживать потерю внутренней влаги в 40 %. Они могут не пить более 2 недель, но добравшись до воды, выпивают ее в количестве многих десятков литров. В кормный период верблюды запасают жир, расходуемый в голодное время — при этом образуется некоторое количество метаболической воды. Большая часть мочевины не выводится из их организма, а гидролизуется и используется в синтезе дефицитных белков. Имеются и другие адаптации, помогающие верблюдам экономить воду.

Одногорбый верблюд (Camelus dromedarius) выдерживает длительное иссушающее воздействие пустыни, теряя до трети собственного веса (вверху). Добравшись до воды, животное быстро восстанавливает свой нормальный вид (внизу).