в оглавление
назад
Созревание половых клеток
В сперматогенезе — развитии мужских половых клеток (сперматозоидов) — сменяют друг друга несколько фаз. Начальная фаза сперматогенеза — разрастание семенных канальцев и размножение в них мелких клеток — сперматогониев; одновременно происходит процесс редукции интерстициальной ткани. В результате деления сперматогонии превращаются в более многочисленные сперматоциты I-го порядка. Дальнейший рост и созревание половых клеток приводят к образованию сперматоцитов II-го порядка. В результате нового деления диплоидное число хромосом превращается в гаплоидное, и такие клетки получают название сперматид. В цитоплазме клеток Сертоли сперматиды снабжаются питательными веществами и превращаются в зрелые половые клетки. Завершение их формирования происходит в придатке семенника. В начальных стадиях развития половые клетки имеют округлую форму. Зрелые сперматозоиды состоят из содержащей ядерный материал головки, шейки и длинного подвижного хвоста. Сперматозоиды чрезвычайно малы (измеряются микронами) но крайне многочисленны; даже у мелких млекопитающих их суммарное количество может исчисляться сотнями миллионов или миллиардами.
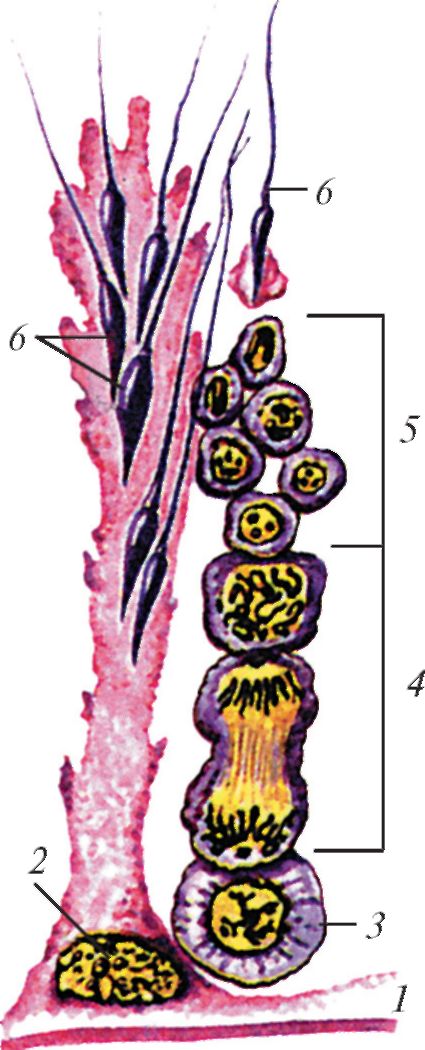
Слева — схема сперматогенеза у млекопитающего: 1 — стенка извитого семенного канальца; 2 — клетка Сертоли; 3 — сперматогония; 4 — сперматоциты; 5 — сперматиды; 6 — сперматозоиды.
Справа — строение сперматозоида млекопитающего: I — головка; II — средняя часть; III — жгутик; 1 — ядро; 2 — акросома; 3 — ядерная оболочка; 4 — шейка; 5 — полосатые столбы; 6 — митохондрии; 7 — аксонема; 8 — цитолемма.
В оогенезе — развитии женских половых клеток — сменяют друг друга три периода: размножение, рост и созревание. Яйцевые клетки, или оогонии, образуются в яичниках из первичных клеток зачаткового эпителия. В первый период развития оогонии многократно делятся, в результате чего возрастает их количество. Во второй — дефинитивные яйцеклетки растут и превращаются в ооциты I-го порядка. Вокруг каждого ооцита формируется фолликул из мелких фолликулярных клеток; соединительнотканные клетки иногда образуют его дополнительную наружную оболочку. Фолликулярные клетки служат для питания растущей яйцеклетки и являются главным источником эстрогенов. По мере своего развития фолликул постепенно перемещается в поверхностную часть яичника. Ко времени размножения ооцит значительно увеличивается за счет накопленного желтка, его оболочка разрастается, а между ней и непосредственно яйцеклеткой образуется пространство, заполненное серозной жидкостью. Фолликул превращается в выдающийся на поверхности яичника граафов пузырек. В период репродуктивной активности оболочка созревшего граафова пузырька лопается, а наполняющая его жидкость вместе с яйцеклеткой выходит в полость тела — этот процесс называется овуляцией. Не достигший овуляции фолликул дегенерирует и превращается в атретическое тело.
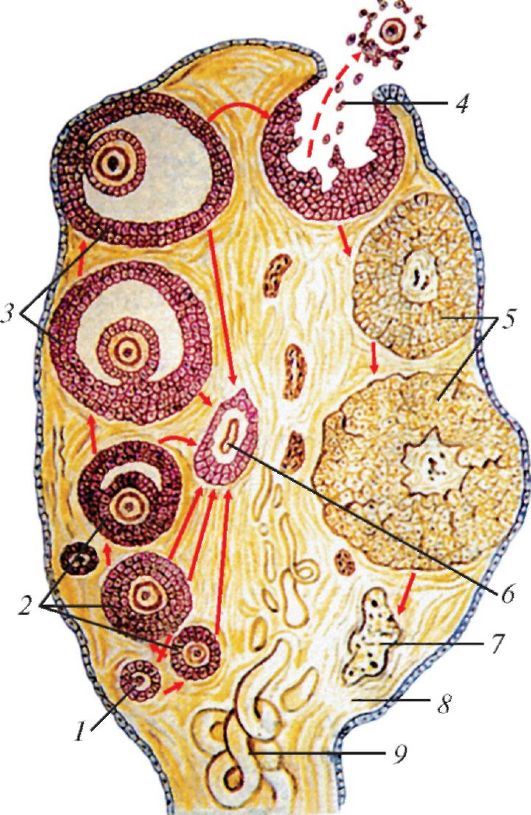
Схема развития фолликулов, овуляции и образования желтых тел в яичнике млекопитающего: 1 — примордиальный фолликул; 2 — первичные фолликулы; 3 — вторичные фолликулы (граафовы пузырьки); 4 — овуляция; 5 — желтые тела; 6 — атретическое тело; 7 — рубец на месте желтого тела; 8 — строма яичника; 9 — кровеносный сосуд.
Овуляция происходит либо спонтанно (самопроизвольно), регулярно повторяясь через определенный промежуток времени, либо как реакция на половой акт (индуцировано). Первый тип овуляции широко распространен, второй свойственен относительно немногим млекопитающим (большинству куниц (Mustelidae), некоторым кошкам (Felidae), зайцам (Leporidae), сусликам (Spermophilus), землеройкам (Soricidae)). Индуцированная овуляция происходит не сразу после спаривания с самцом, но с некоторой задержкой (10-12 часов у крольчихи (Oryctolagus cuniculus), до суток и более у кошки (Felis silvestris), почти до 2 суток у самки хорька (Mustela putorius)). Это дает самке время спариться и с другими самцами, вследствие чего яйцеклетки оплодотворяются спермой разных партнеров и для будущего приплода характерно множественное отцовство — суперфекундация. У кошек спаривание с самки с несколькими самцами предположительно связано с многочисленными дефектами сперматозоидов, в норме встречающимися у большинства кошачьих самцов, т. е. тем самым самка увеличивает шансы на удачное оплодотворение своих яйцеклеток и увеличивает свою плодовитость. При однократном спаривании с одним самцом значительная часть яйцеклеток могла бы остаться неоплодотворенной. Другим способом устранения эффекта низкой оплодотворяющей способности являются многократные спаривания с одним партнером.
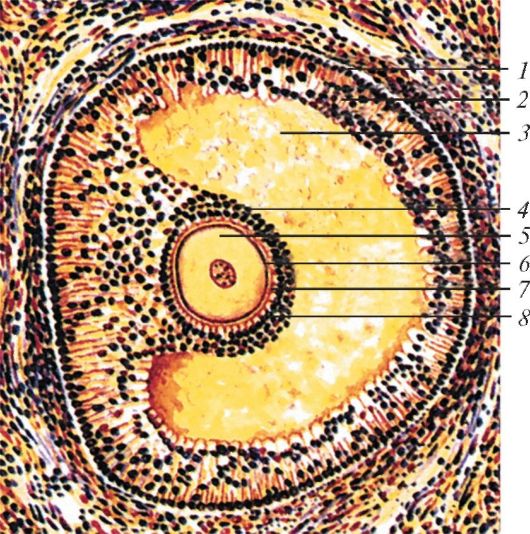
Строение граафова пузырька: 1 — наружная оболочка фолликула; 2 — внутренняя оболочка фолликула; 3 — полость фолликула с фолликулярной жидкостью; 4 — яйценосный холмик; 5 — яйцеклетка; 6 — блестящая оболочка; 7 — лучистый венец; 8 — фолликулярные клетки.
Образовавшийся на месте опустевшего граафового пузырька рубец заполняется кровяным сгустком. Затем здесь из оставшихся периферических клеток фолликулярного эпителия развивается новый клеточный слой — т. н. желтое тело (corpus luteum). Сформировавшееся таким способом небольшое желтое тело овуляции у самок, оставшихся холостыми, вскоре резорбируется. У оплодотворенных самок оно разрастается и превращается в желтое тело беременности, состоящее из центральной части, или ядра, и коркового слоя. Ядро содержит молодую соединительную ткань с сосудами и часто со сгустками крови; корковый слой слагается из соединительной ткани с лютеиновыми клетками, выделяющими пигмент липохром характерного желтого цвета. Желтое тело беременности выполняет функции временного органа внутренней секреции. Оно выделяет гормон прогестерон, который стимулирует имплантацию зародыша в стенку матки, одновременно задерживая образование новых графовых пузырьков и овуляцию до окончания беременности.
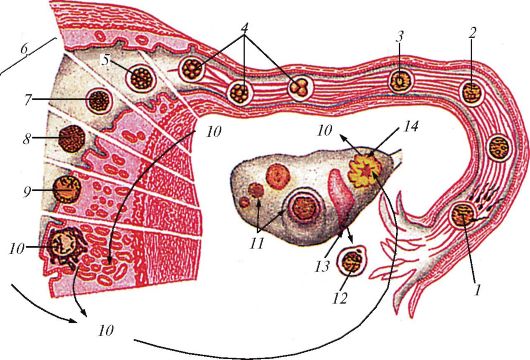
Схема путей яйцеклетки по маточной трубе и имплантации зародыша в эндометрий у человека: 1 — проникновение сперматозоида в яйцеклетку, начало второго созревания; 2 — слияние пронуклеусов; 3 — первое деление дробления; 4 — раннее дробление; 5 — морула; 6 — два дня до имплантации; 7 — ранняя гаструла; 8 — поздняя гаструла; 9 — начало имплантации; 10 — растущий имплантированный эмбрион вырабатывает хронический гонадотропин, который в дальнейшем поддерживает желтое тело и таким образом обеспечивает снабжение прогестероном, который поддерживает эндометрий; 11 — фолликулостимулирующий гормон стимулирует созревание фолликула; 12 — первое полярное тельце, веретено созревания; 13 — лютеинизирующий гормон вызывает овуляцию; 14 — лютеинизирующий гормон некоторое время поддерживает желтое тело.
Выпавшие из яичника яйцеклетки попадают через полость тела в воронку яйцевода и затем в фаллопиеву трубу. После выделения направительных телец они превращаются в ооциты II-го порядка и, наконец, в зрелые яйца с гаплоидным числом хромосом. Зрелые яйца млекопитающих имеют шаровидную форму. В связи с принципиальными отличиями в размножении у основных групп млекопитающих отличен и характер яиц. У однопроходных они достигают относительно крупных размеров — 2,5-4,0 мм, а в матке увеличиваются до 10-20 мм. Эти яйца богаты желтком (меробластический тип) и, подобно яйцам птиц и крокодилов, имеют хорошо развитую белковую оболочку, служащую основным запасом воды. Снаружи они покрыты плотной пергаментообразной оболочкой. У однопроходных одновременно созревает от 1 до 3 яиц. Яйца сумчатых значительно мельче (0,2-0,4 мм), что обусловлено обеднением яйца желтком; белковая оболочка также развита слабо. У большинства видов одновременно развиваются единицы яиц и лишь у опоссумов — иногда более 10. У плацентарных яйца не имеют белковой оболочки и практически вовсе лишены желтка (голобластический тип), вследствие чего они микроскопически малы (0,05-0,2 мм). Количество вызревающих единовременно яиц варьирует в соответствии с большей или меньшей плодовитостью отдельных видов, колеблясь от 1 до 20 и даже более.
Развитие зародыша
В процессе внутриутробного развития, или эмбриогенезе, друг друга последовательно сменяют стадии зиготы, морулы, бластулы, гаструлы, нейрулы и эмбриона.
Оплодотворение яйца происходит в фаллопиевой трубе (у тенреков (Tenrecidae) — в яичнике); неоплодотворенные яйца вскоре погибают и рассасываются. В результате слияния мужской и женской половых клеток образуется зигота. Для зиготы характерен двойной (диплоидный) набор хромосом, свойственный соматическим клеткам. На последующих стадиях эмбрионального развития зигота служит единственным источником для образования всех тканей и органов тела животного.
Образовавшись, зигота начинает дробиться посредством митоза, образуя под яйцевой оболочкой дочерние клетки затем развивается плацента. Ее связь с зародышем поддерживается за счет кровеносной системы аллантоиса — бластомеры. На стадии морулы бластомеры дифференцируется на два слоя. Более крупные из них образуют наружный слой — трофобласт, в дальнейшем участвующий в имплантации (обеспечивает прикрепление эмбриона к эпителию матки, инвазию внутрь эндометрия и разрушение кровеносных сосудов, оказывает иммуносупрессорное действие), а также в формировании эктодермы ворсинок хориона (эктодермальная часть плаценты). Бластомеры меньшего размера образуют внутренний слой — эмбриобласт, дающий начало собственно телу плода, а также мезодермальным и энтодермальным структурам внезародышевых органов (желточному мешку, аллантоису, амниону, мезодермальной части хориона).
У меробластических яиц однопроходных дробление захватывает лишь небольшой участок цитоплазмы на анимальном полюсе, где в дальнейшем формируется зародышевый диск. Щелевидная первичная полость (бластоцель) расположена между ним и подстилающим желтком. Зигота голобластических яиц проходит полное дробление, хотя обычно и неравномерное, так что бластомеры крупнее на стороне вегетативного полюса, чем на противоположном. Бластула получается шаровидной, с бластоцелем внутри.
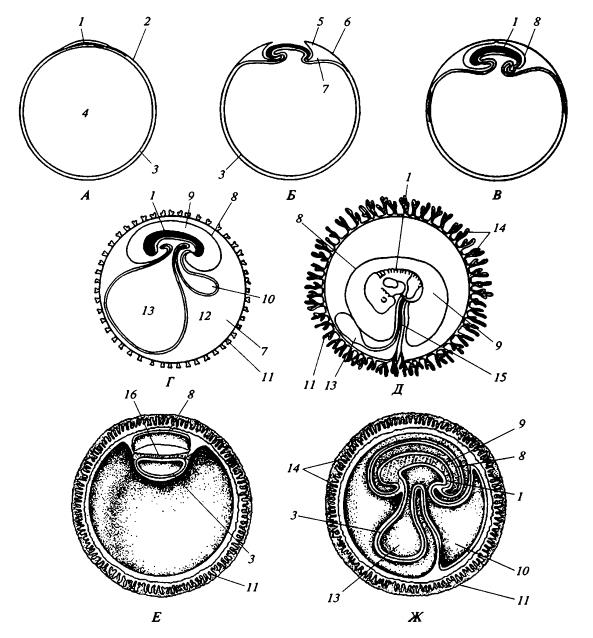
Схема развития зародышевых оболочек у млекопитающих: А-Д — последовательные стадии в типичном варианте; Е, Ж — на примере человека: 1 — зародыш; 2 — эктодерма; 3 — энтодерма; 4 — полость зародышевого пузыря, предшественник полости желточного пузыря; 5 — амниотическая складка: 6 — трофобласт; 7 — внезародышевый цепом (экзоцелом); 8 — амнион; 9 — амниотическая полость; 10 — аллантоис; 11 — хорион; 12 — мезодерма; 13 — полость кишечника и желточного мешка; 14 — ворсинки хориона; 15 — пупочный канатик; 16 — бластодерма.
Гаструляция голобластического яйца сопровождается двумя сопряженными процессами — инвагинацией, т. е. вворачиванием более крупных клеток вегетативного полюса, и эпиболией, т. е. обрастанием этих клеток пластом более мелких бластомеров. В результате образуется второй слой клеток, выстилающих первичную кишку, или гастроцель. Получается двухслойная гаструла, обладающая наружным, или эктодермальным, и внутренним, или энтодермальным клеточными слоями. Промежуточный клеточный слой — мезодерма — образуется в области губ бластопора, т. е. в зоне контакта эктодермы и энтодермы, откуда она постепенно распространяется внутрь зародыша.
Следующее важное событие в онтогенезе — нейруляция. Из эктодермы головного отростка выделяется нервная пластинка, окруженная нервным валиком. Пластинка погружается, сворачиваясь в трубку, и отшнуровывается от остальной эктодермы, которая смыкается над ней. Нервные валики погружаются параллельно с нервной трубкой и образуют по бокам от нее т. н. нервные гребни, скопления индивидуально мигрирующих клеток с очень высокими и разнообразными формообразовательными способностями. От спинной стороны первичной кишки — как раз напротив нервной трубки — отшнуровывается продольный тяж, хорда, а по бокам от нее тем же способом формируются мезодермальные зачатки. Позже они сегментируются, подразделяясь на сомиты, в которых за счет расщепления клеточных масс возникают полости.
Перистальтическим сокращением стенок фаллопиевой трубы и соответствующими движениями ресничек ее эпителия развивающееся яйцо продвигается в матку. Для нее характерно сильное развитие крупных кровеносных сосудов, доставляющих зародышу питательный материал, и сильных мускулистых стенок, с помощью которых созревший зародыш выводится наружу. При переходе в матку зародыш освобождается от покрывающей его яйцевой оболочки и функцию наружной оболочки принимает на себя трофобласт. В слизистой оболочке матки имеются железы, секрет которых — эмбриотроф («маточное молоко») в первое время обеспечивает питание зародыша. Затем зародыш прикрепляется к внутренней поверхности матки. Клетки трофобласта оказывают биохимическое воздействие на сильно измененную, набухшую маточную стенку, вызывая в месте соприкосновения ответные реакции с ее стороны. Зародыш постепенно углубляется в эндометрий, пока не достигает его внутреннего слоя — происходит имплантация.
Вступив в столь плотный контакт с материнским организмом, зародыш начинает быстро расти, происходит дифференцировка его элементов. Жизнедеятельность зародыша обеспечивают зародышевые оболочки, которые образуются за счет внезародышевых частей зародышевых листков. Ближе всего к зародышу расположен заполненный жидкостью амнион. Он формируется либо боковыми складками внезародышевой эктодермы и мезодермы, которые приподнимаются и смыкаются над зародышем (у большинства млекопитающих), либо путем образования полости среди зародышевых клеток, постепенно преобразующихся в окружающую зародыш оболочку (у приматов). Амнион сильно разрастается и окружает зародыш почти со всех сторон. Благодаря этому зародыш защищен от вредных сотрясений, которые испытывает организм матери при быстром передвижении, прыжках и т. д., и развитие его фактически происходит в водянистой среде. Хорион непосредственно контактирует со стенкой матки, обеспечивая обмен веществ между организмом матери и плодом. Он образуется из трофобласта, который затем подстилается внезародышевой мезодермой. На поверхности хориона расположены ворсинки, которые вначале представляют собой разрастания эктодермальных клеток трофобласта в тканях матки (первичные ворсинки). Аллантоис служит эмбриональным органом дыхания и для сбора продуктов обмена веществ зародыша. Он закладывается как вырост заднего отдела кишечной трубки. Разрастаясь, аллантоис внедряется в пространство между амнионом и хорионом. Его сосуды врастают в ворсинки хориона (вторичные ворсинки) и вместе с ними формируют плодную часть аллантохориальной плаценты. В мезенхиме аллантоиса образуются сосуды пуповины, связывающие плаценту с зародышем. На более поздних стадиях развития из внутризародышевой части аллантоиса образуется мочевой пузырь или формируется клоака. Выстланный энтодермой желточный мешок выполняет функцию переноса веществ между материнским организмом и зародышем у сумчатых и на начальных стадиях развития плацентарных. У последних он пустой. Из части желточного мешка формируется кишечная трубка.
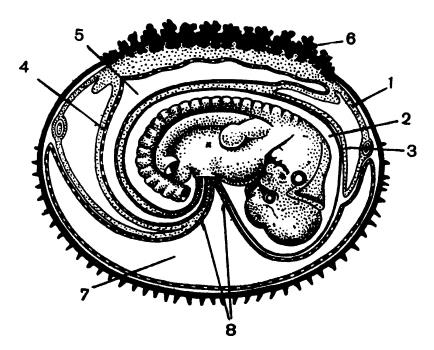
Зародыш кролика (Oryctolagus cuniculus) в конце 12-го дня развития: 1 — серозная оболочка; 2 — амниотическая полость; 3 — амнион; 4 — аллантоис; 5 — полость аллантоиса; 6 — утолщенная часть хориона; 7 — полость желточного мешка; 8 — пупочный канатик.
Судьба зародышевых листков, за счет которых формируются все ткани и органы животного организма, заслуживает более подробного рассмотрения. Итак, из эктодермы вначале развивается эпителий кожного типа, который затем образует общую (ненейральную) эктодерму и нейродерму.
Общая эктодерма образует поверхностный (эпителиальный) кожный слой, или эпидермис, покрывающий тело снаружи и заходящий внутрь на обоих концах пищеварительного тракта, выстилая ротовую полость и самый конец клоаки (в случае, если она имеется). Производными эпидермиса являются волосы, различные роговые образования (роговые чехлы, ногти, чешуи) и кожные железы (млечные, потовые и др.). Общей эктодермой также образован сенсорный эпителий полости носа и внутреннего уха, глазной хрусталик, зубная эмаль.
Нейродерма служит источником образования центрального и периферического отделов нервной системы с их вспомогательными эпендимоглиальными элементами. Нервные гребни дают сенсорные ганглии, миелиновые оболочки нервных волокон и некоторые ненейральные структуры, например пигментные клетки, одонтобласты (откладывающие зубной дентин), висцеральный скелет и отчасти мозговой череп, сухожилия висцеральных мышц головы и т. д. Сетчатка глаза также формируется из нейродермы.
Из энтодермы развивается выстилка кишечной трубки, за отмеченными выше исключениями, а также ее железистый аппарат, включая печень и поджелудочную железу. Легкие, будучи производными кишечной трубки, изнутри тоже выстланы энтодермой. В области ротоглотки энтодерма, соединяясь с эктодермой, образует переходную пластинку, участвующую в образовании эпителиальной выстилки ротоглотки, пищевода, дыхательных путей, стромы тимуса, а также железистой ткани ряда эндокринных желез — гипофиза (аденогипофиз), щитовидной и паращитовидных желез. Кроме того, энтодермой сформированы концевые участки мочеполового аппарата.
Мезодерма сначала представлена сомитами, которые метамерно располагаются по обеим сторонам хорды и посредством сегментальных выростов, или нефротомов, соединены с вентральными несегментированными боковыми пластинками мезодермы, или спланхнотомами. Каждый сомит, начиная с третьего от краниального конца тела зародыша, дифференцируется на три участка, из которых дорсолатеральный, или дерматом, служит зачатком соединительнотканной основы дермы, дорсальный участок, или склеротом — зачатком хрящевой и костной тканей скелета, а третий участок, располагающийся между дерматомом и склеротомом, является зачатком скелетной мускулатуры, или миотомом. Нефротом, располагаясь вдоль тела зародыша на протяжении от его краниального конца до каудального, принимает участие в развитии органов мочеполового аппарата.
Спланхнотом представлен соматоплеврой и спланхноплеврой. Соматоплевра образуется экто- и мезодермой, соединенными между собой мезенхимой и с внутренней поверхности выстланной пристеночным, или париетальным листком. Спланхноплевра образуется энтодермой и спланхнической мезодермой, составляющими основу стенки пищеварительной трубки, покрытой снаружи внутренностным, или висцеральным, листком, являющимся серозной оболочкой внутренних органов. Между париетальным и висцеральным листками заключается вторичная полость тела, или целом.
Мезенхима, или первичная соединительная ткань, заполняет все промежутки между зародышевыми листками и зачатками органов, как в теле зародыша, так и за его преде-лами. Именно из нее формируются кровь и лимфа, развиваются кровеносные сосуды, кроветворные органы, сердечная мышца и различные виды соединительной ткани.
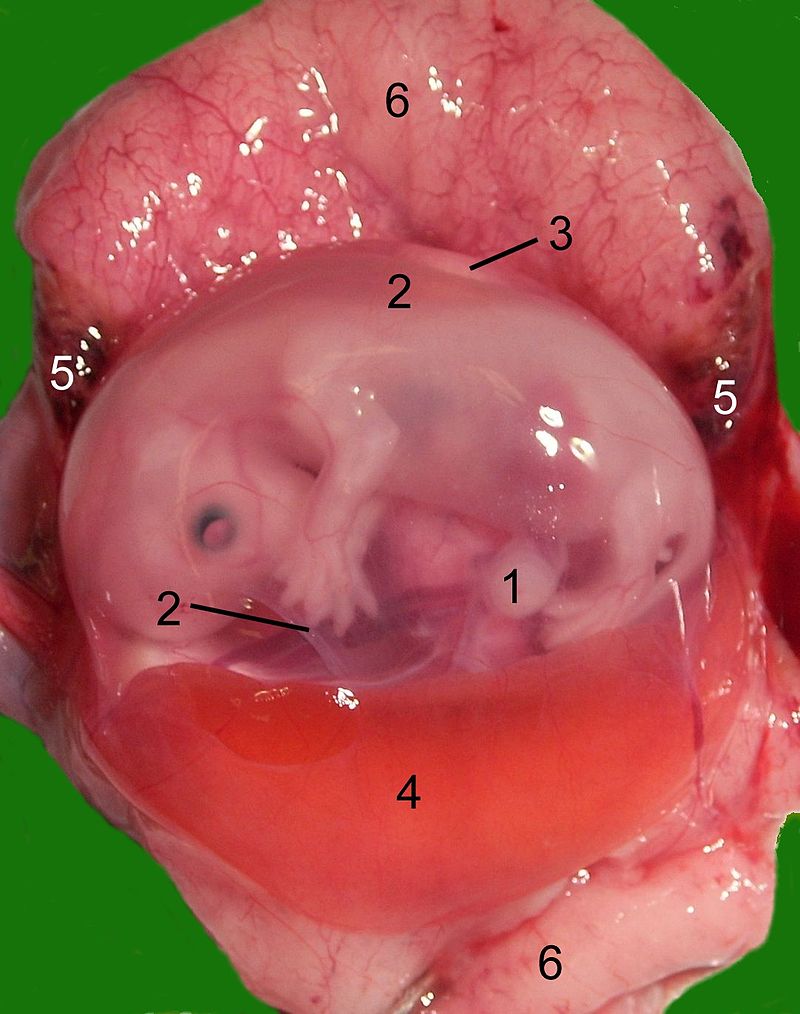
Извлеченный из матки зародыш домашней кошки (Felis silvestris catus): 1 — пуповина; 2 — амнион; 3 — аллантоис; 4 — желточный мешок; 5 — развивающаяся гематома; 6 — материнская часть плаценты.
Вынашивание зародыша
У однопроходных формируются небольшие, покрытые мягкой скорлупой яйца, сходные по строению с яйцами рептилий. Питание зародыша происходит за счет желткового мешка. Выходящий из яйца эмбрионоподобный детеныш нуждается в длительной материнской заботе. Были ли яйцекладущими различные мезозойские млекопитающие, конечно, точно неизвестно, однако у некоторых из них просвет таза кажется слишком узким, чтобы пропустить яйцо. Это наводит на мысль о раннем приобретении живорождения, по крайней мере, в некоторых группах.
У сумчатых и плацентарных для связи плода с материнским организмом развивается временный орган — детское место, или плацента (placenta). Она формируется вследствие срастания слизистой оболочкой матки (материнская часть плаценты) с желточным мешком и хорионом (зародышевая часть плаценты). Примечательно, что вокруг зародыша сумчатых на начальных стадиях развития формируется скорлуповая оболочка, но в дальнейшем она исчезает.
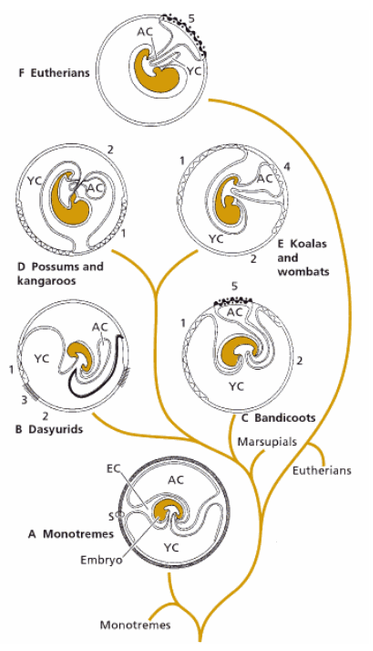
Различные типы плацент млекопитающих: А — однопроходные; В — хищные сумчатые; С — бандикуты; D — поссумы и кенгуру; Е — коала и вомбаты; F — эвтерии; 1 — сосудистая хориовителлиновая плацента; 2 — несосудистая хориовителлиновая плацента; 3 — синцитиальная хориовителлиновая плацента; 4 — комбинированная (несосудистая) аллантохориальная плацента; 5 — синцитиальная (сосудистая) аллантохориальная плацента; АС — аллантоис; ЕС — внезародышевая полость; S — внешняя оболочка; YC — желточный мешок.
У сумчатых взаимоотношения материнского организма с плодом сравнительно просты. Поверхность его желточного мешка более или менее плотно срастается со слизистой оболочкой матки, вызывая частичный гистолиз ее поверхностных слоев. Специальное образование, обеспечивающее приток питательных веществ, отсутствует; зародыш питается за счет желтка яйца, секреторных выделений клеток стенки матки (гистотрофия) и транссудата крови матери (гематотрофия). Газообмен зародыша осуществляется обильно снабженной сосудами частью желточного мешка. Такая примитивная протоплацента именуется желточной, или омфалоидной.
Зародыш плацентарных входит с организмом матери в гораздо более тесный контакт. В начале его развития функционирует желточная плацента, которая затем в обязательном порядке заменяется настоящей аллантохориальной (у некоторых плацентарных, например, у лошадей (Equus), желточная плацента действует довольно долго). Сформировавшись, аллантохорион прирастает к слизистой оболочке матки, а его ворсинки внедряются в богатые кровеносными сосудами внутренние слои слизистой, в этом месте разрыхляющиеся. Именно сросшиеся части аллантохориона и слизистой оболочки матки и образуют аллантохориальную плаценту. Она является очень сложным органом, и ее развитие продолжается довольно долго (так, например, у человека образование плаценты заканчивается только на 3-м месяце беременности).
Прямое сообщение между сосудами ворсинок и слизистой матки отсутствует, и весь обмен веществ совершается путем осмоса через сильно утонченные стенки ворсинок, сквозь которые легко проходят небольшие молекулы, но не могут пройти ни белки крови, ни ее форменные элементы. Разросшиеся ворсинки значительно увеличивают всасывающую поверхность: например, у человеческого зародыша их суммарная длина достигает 48 км. Из ножки эмбриона плацентарных развивается длинный пупочный канатик (funiculus umbilicalis), содержащий пупочную артерию и пупочную вену. Пупочная артерия отходит от аорты зародыша и образует в аллантохорионе капиллярные сети, которые и вступают в тесное соприкосновение с сосудами материнского организма. Капилляры хориона собираются в пупочную вену, впадающую в заднюю полую вену зародыша.
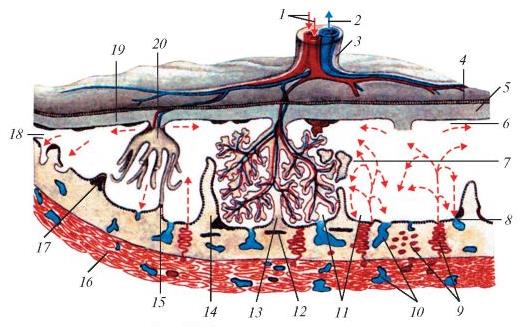
Схема строения плаценты человека: 1 — пупочная артерия; 2 — пупочная вена; 3 — пупочный канатик; 4 — амниотический эпителий; 5 — пластинка хориона; 6 — субхориоидальная щель; 7 — клеточный островок; 8 — базальный трофобласт; 9 — спиральные артерии; 10 — децидуальные вены; 11 — межворсинчатое пространство; 12 — фибриновая полоска; 13 — зона контакта ворсинок; 14 — плацентарная перегородка; 15 — миометрий; 16 — децидуальная оболочка; 17 — трубчатый фибрин; 18 — краевой синус; 19 — базальная децидуальная оболочка; 20 — котиледон. Стрелками обозначено направление тока материнской крови в межворсинчатом пространстве плаценты.
Питание плода осуществляется за счет кислорода и питательных веществ, поступающих из крови матери, т. е. гемотрофно (легкие плода не функционируют до самого рождения). В свою очередь, через плаценту идет выделение из зародыша продуктов обмена и углекислоты. Также плацента предохраняет зародыш от проникновения инфекций и вредных веществ. Через плаценту (а после рождения — через молоко) потомство снабжается материнскими антителами, что повышает его сопротивляемость болезням. Самой плацентой продуцируются секреты, действующие как на плод, так и на организм матери. Особое значение плаценты состоит в том, что она образует между организмом матери и эмбрионом активный барьер, препятствующий его отторжению вследствие несовместимости тканей. В комплексе вышеперечисленное позволяет плацентарным млекопитающим иметь продолжительное внутриутробное развитие и рождать более сформированных детенышей.
Усложнение структуры аллантохориальной плаценты происходит параллельно с установлением более тесной связи развивающегося зародыша с организмом матери. Наиболее простое строение имеет эпителиохориальная плацента, или полуплацента, встречающаяся у лемуров (Lemuriformes), нежвачных (Suina), верблюдов (Camelidae), лошадей (Equidae) и ряда других млекопитающих. При ее образовании на поверхности хориона появляются ворсинки в форме небольших бугорков. Они погружаются в соответствующие углубления слизистой оболочки матки, не срастаясь с ней и не производя никаких разрушений в ее тканях. При родах ворсинки выходят из своих углублений безболезненно и без кровотечений. Десмохориальная плацента свойственна жвачным (Ruminantia) и характеризуется установлением более тесной связи хориона зародыша со стенкой матки. В месте соприкосновения с ворсинками хориона эпителий слизистой оболочки разрушается. Разветвленные ворсинки погружаются в соединительную ткань, приближаясь, таким образом, к кровеносным сосудам матери. Эндотелиохориальная плацента встречается у хищных и характеризуется разрушениями не только эпителия, но и соединительнотканной прослойки. Ворсинки хориона тесно соприкасаются с сосудами и отделены от материнской крови только их тонкой эндотелиальной стенкой. Гемохориальная плацента свойственна насекомоядным, грызунам и большинству приматов, включая человека. При установлении связи плода с материнским организмом в матке происходят глубокие изменения: частично разрушается соединительная ткань и даже стенки сосудов. На месте разрушенных тканей образуются соединительнотканные лакуны, наполненные кровью матери, которая изливается из ее сосудов. Ворсинки хориона омываются кровью и всасывают из нее необходимые вещества. Согласно расчетам, по эффективности транспортирования питательных веществ от матери к зародышу гемохориальная плацента в 250 раз превосходит эпителихориальную. Однако у ряда грызунов описана еще более совершенная гемоэндотелиальная плацента. В этом случае дополнительно происходит сильное истончение хориального эпителия, так что его наличие обнаруживается только методами электронной микроскопии. Таким образом, кровь матери у этих животных максимально приближена к внутренней стенке капилляров плода. Вследствие тесного контакта между зародышем и материнским организмом роды у млекопитающих со сложными плацентами сопровождаются отторжением значительной части стенки матки и более или менее обильными кровотечениями.
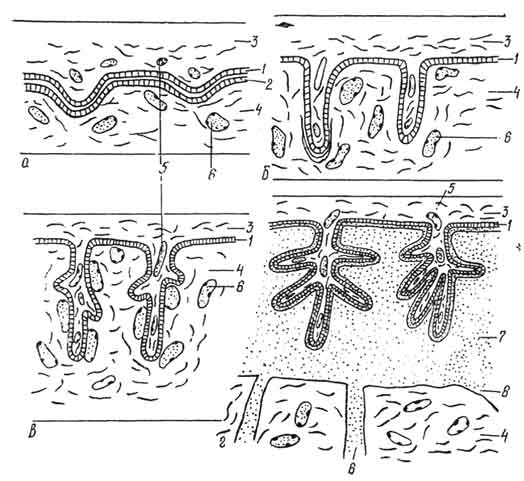
Типы гистологического строения плацент: а — эпителиохориальная; б — десмохориальная; в — эндотелиохориальная; г — гемохориальная; 1 — эпителий хориона; 2 — эпителий стенки матки; 3 — соединительная ткань ворсинки хориона; 4 — соединительная ткань стенки матки; 5 — кровеносные сосуды ворсинок хориона; 6 — кровеносные сосуды стенки матки; 7 — материнская кровь.
Плаценты различаются и по типу размещения ворсинок. В диффузной семиплаценте (semiplacenta diffusa) ворсинки равномерно рассеяны по всей поверхности хориона; это характерно для мокроносых приматов, китообразных, большей части копытных — свиней, верблюдов, лошадей и т. д., а также некоторых ксенартр. В котиледонной, или дольчатой семиплаценте (semiplacenta cotyledonaria, или semiplacenta multiplex) ворсинки собраны в сидящие на бугорках группы — котиледоны, распределенные по хориону относительно редко; такая плацента свойственна жвачным. В кольцевой, или зональной, плаценте (placenta zonaria) ворсинки распределены по хориону в виде широкого пояса, охватывающего зародыш поперек; подобное наблюдается у трубкозубых, сирен, хоботных, некоторых броненосцев и многих хищных с ластоногими включительно. В дискоидальной плаценте (placenta discoidalis) ворсинки сосредоточены в одном месте хориона, образуя диск; этот тип встречается у насекомоядных, рукокрылых, сухоносых приматов, грызунов, зайцеобразных и многих ксенартр — броненосцев (Dasypus), муравьедов (Myrmecophaga) и ленивцев (Bradypus).
По степени связанности с маткой различаются плаценты неотпадающие и отпадающие. Неотпадающая плацента (placenta adeciduata) характеризуется слабой связью с маточной стенкой, благодаря чему при родах матка не повреждается. Таковыми являются диффузная и дольчатая, в некоторых случаях — зональная и дискоидальная плаценты, свойственные части сумчатых, панголинам, нежвачным, верблюдам, китообразным, сиренам, лошадям и мокроносым приматам. Отпадающая плацента (placenta deciduata), наоборот, настолько тесно срастается со стенкой матки, что при родах часть ее слизистой оболочки отрывается и в составе последа выводится наружу. В такой плаценте различается материнская, или маточная — отторгающаяся часть слизистой оболочки матки, и зародышевая часть — ворсинки аллантохориона. К отпадающим принадлежат большинство зональных и дискоидальных плацент, характерных для ксенартр, насекомоядных, грызунов, рукокрылых, шерстокрылов и сухоносых приматов. Между двумя этими основными типами существует переходная форма, присутствующая у жвачных.
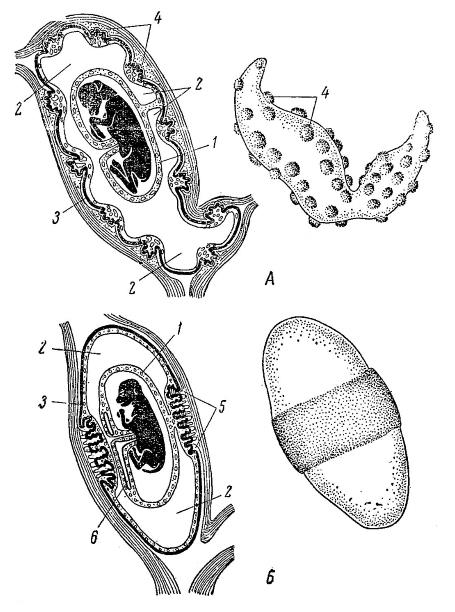
Схема строения плаценты; слева — разрез через матку, справа — внешний вид: А — дольчатая плацента жвачных; Б — кольцевая плацента хищных; 1 — амнион; 2 — аллантоидный пузырь; 3 — хорион; 4 — котиледоны; 5 — кольцевая плацента; 6 — рудимент желточного мешка.
Когда при родах плод проходит через шейку матки, его оболочки разрываются, и наружу выливается амниотическая жидкость. После рождения плода выделяется плацента, и обильное сначала кровотечение постепенно приостанавливается. В матке начинается регенерация эпителия слизистой оболочки. После родов в стенке матки на месте имплантации эмбрионов остаются темные, т. н. плацентарные пятна. Их подсчет используют для установления количества детенышей в помете и числа пометов в сезон размножения.
Примечательно, что крупные плацентарные млекопитающие обычно рождают относительно мелких детенышей по сравнению с плацентарными малых размеров. Например, даже стокилограммовый вес новорожденного слоненка едва составляет 4 % массы слонихи, тогда как масса только что родившейся летучей мыши (приблизительно 2,45 г) — почти треть веса (29,5 %) ее матери. Еще более выразительны эти данные, если исходить из массы всего помета: общая масса новорожденных кроликов составляет 19 % от массы самки, а новорожденных полевок — более 50 %. Эти различия объясняются, конечно, не только разницей в размерах, но и различной способностью питания плода через плаценту, имеющую относительно меньшую поверхность у крупных млекопитающих. Имеются у этого несоответствия и приспособительные причины. Впрочем, из данного правила есть немало исключений.
Развитие зародыша внутри организма матери стало возможным не только благодаря преобразованию зародышевых оболочек и возникновению плаценты, но и вследствие сложных приспособлений материнского организма к длительному состоянию беременности. В этом приспособлении большую роль играет печень, почки, железы внутренней секреции и другие органы.
вперед
в оглавление