в оглавление
назад
Волосяной покров (продолжение)
Особую категорию кожных производных составляют вибриссы (vibrissae) — особенно крупные, резко выдающиеся над общим меховым покровом слабоизогнутые жесткие волосовидные образования с мостовидным типом строения кожицы. Они единично или рядами располагаются на голове, особенно обильны вокруг рта, на верхней челюсти по обе стороны носа (т. н. «усы»), имеются в виде отдельных пучков над глазами и в нижней части подбородка. Вибриссы присутствуют и на других, наиболее подверженных механическим воздействиям частях тела — на кончиках ушей, на нижней стороне шеи, на груди, на передней и внешней стороне плеч, вблизи запястий и предплюсн, на боках туловища. У некоторых лазающих форм (например, у белок (Sciurinae)) вибриссы имеются и на брюхе, а у живущих под землей кротов (Talpinae) — даже на кончике хвоста. Вибриссы могут двигаться либо волевым усилием («активные»), либо непроизвольно, либо вообще не двигаться («пассивные»). У грызунов вибриссы снабжены крупными мышцами и активно двигаются. У мартышковых обезьян (Cercopithecidae) редкие (2-3 пучка) лицевые вибриссы не имеют собственных мышц и приводятся в движение мимической мускулатурой. У лошадей (Equidae), полорогих (Bovidae) и свиней (Suidae) они неподвижны. При общей редукции волосяного покрова вибриссы сохраняются и даже могут равномерно распределяться по телу — как у сирен.
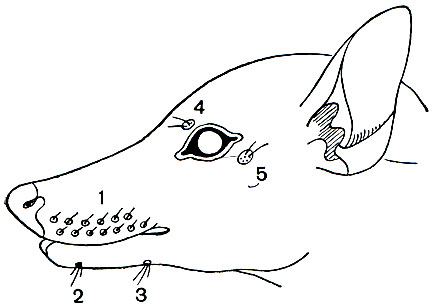
Схема головы лемура с полным набором пучков вибрисс: 1 — верхнечелюстной; 2 — подбородочный; 3 — предчелюстной (непарный); 4 — надглазничный; 5 — заглазничный.
Функция вибрисс — осязательная: они формируют вокруг головы животного сенсорное поле, с помощью которого оно получает информацию о находящихся вблизи объектах. В основании и в стенках волосяных мешков вибрисс располагаются многочисленные нервные окончания, воспринимающие давление от соприкосновения стержня с посторонними предметами. Волосяные мешки вибрисс головы иннервируются ветвями тройничного нерва и регулируются поперечнополосатыми мышцами. Окружающие их корни венозные лакуны служат, по-видимому, амортизаторами сотрясений, передаваемых стержнем. Каждый вибрисс посылает в мозг информацию только в том случае, если его будут сгибать в определенном направлении. Вибриссы разных участков тела настроены на восприятие различно направленных движений. Анализируя полученную информацию, мозг учитывает характер и направление движения частей тела животного, снабженных вибриссами. Это позволяет получить исчерпывающую информацию о том, имеет ли дело хозяин вибрисс с неподвижным предметом или с живым существом, каковы его размеры и вес.

У новорожденной домовой мыши (Mus musculus) имеются вибриссы по бокам морды (1). Каждый вибрисс соединен с нейроном. Пучки нейронов имеют определенную локализацию в мозге (2). Удаление вибрисс (3) (закрашенные черные кружочки) приводит к тому, что соответствующие пучки нейронов прекращают развиваться и в конечном счете исчезают.
Необходимость вибрисс не вызывает сомнений, особенно там, где пасует зрение. Кошки (Felidae), преимущественно охотящиеся ночью, да еще в густых зарослях, имеют более развитые вибриссы, чем собаки (Canidae), предпочитающие открытые пространства и не стесняющиеся совершать свои охотничьи вылазки засветло. Отличные вибриссы имеют норные животные — грызуны и насекомоядные. Используют вибриссы и водные звери — бобры (Castor), выдры (Lutrinae), ластоногие (Pinnipedia). Им приходится заглядывать под коряги и камни, ловить добычу в зарослях водных растений, хватать ее со дна. На илистом грунте при малейшем движении поднимается облако мути, и зрение мгновенно делается бесполезным. Вынесение чувствительных рецепторов на морду и челюсти — рабочие аппараты хищника — облегчает выполнение целенаправленных движений при поимке добычи.

Длинные вибриссы большой песчанки (Rhombomys opimus) создают вокруг ее морды чувствительное поле, сравнимое по диаметру с длиной туловища этого зверька. Вибриссы направлены слегка вперед, поэтому песчанка, странствуя в темноте, еще за несколько сантиметров от кончика носа почувствует, что уперлась в стенку.
Видимо, вибриссы используются и как органы дистантной рецепции. Маленькая птица, взлетевшая из-под носа камышового кота (Felis chaus), пробирающегося в ночной темноте по густым зарослям, вызовет движение воздуха, которое, судя по чувствительности вибрисс, зверь не может не заметить. В воде, как в более плотной среде, дистантная рецепция функционирует надежнее. Сивуч (Eumetopias jubatus), преследующий рыбу, должен на близких дистанциях ощущать движение хвоста удирающей жертвы. Вероятно также, что вибриссы используются как органы активной локации. Волны давления, вызываемые бегущей кошкой, а тем более плывущим в глубине тюленем, встретив на пути солидное препятствие, отразятся от него и, вернувшись назад, не минуют зверя. Активная локация должна использоваться норными животными. Крот, видимо, с определенным умыслом строит свои замысловатые галереи весьма узкими. При движении по ним тело животного плотно прилегает к стенкам норы. Действуя как поршень, крот проталкивает впереди себя столб воздуха. В норе волны давления значительно сильнее, чем на открытом воздухе. Надо думать, крот не оставляет без внимания информацию, которую несут отраженные волны.

Хорошо развитые вибриссы моржа (Odobenus rosmarus) помогают ему находить пищу на морском дне.
У усатых китов (Mysticeti) несколько рядов вибрисс расположено на поверхности головы и по краям верхней челюсти. Они представляют собой не рудименты некогда хорошо развитого волосяного покрова, а вполне специализированные органы осязания в виде редких щетинковидных волосков толщиной 0,2-0,4 мм и в длину до 1 см. Общее их количество невелико: около 250 у гренландского кита (Balaena mysticetus), 50-100 у полосатиков (Balaenoptera). Зачем вибриссы усатым китам? Большинство из них питается мелкими кальмарами, рыбами и планктонными ракообразными. Между тем глаза у кита расположены так, что он не видит своей жертвы, когда она находится совсем близко. Это «мертвое» пространство, и информацию отсюда поставляет в мозг лишь осязательный аппарат вибрисс. Когда голова животного попадает в скопление криля и рачки начинают беспрерывно задевать его усы, кит узнает, что пора открывать рот. Чувствительность вибрисс феноменальна. К каждой волосяной сумке подходит от 400 до 10000 нервных волокон.
Среди зубатых китов (Odontoceti) вибриссы распространены значительно меньше. У нарвала (Monodon monoceros) и белухи (Delphinapterus leucas) их не бывает. У кашалота (Physeter macrocephalus) и гринд (Globicephala) они исчезают еще до рождения. Большинство настоящих дельфинов (Delphinidae) обладают вибриссами в младенческом возрасте. В 1-2 месяца начинается их редукция, а в 3-5 месяцев они уже исчезают. У взрослых особей в коже морды остаются «ямки» после дегенерации этих ювенильных вибрисс, и, видимо, происходит инволюция их луковиц в специализированные тактильные органы, реагирующие на изменение давления воды и низкочастотные колебания. У гвианского дельфина (Sotalia guianensis) зафиксирована способность этих органов улавливать слабые электрические импульсы, происходящие от добычи. В зрелом состоянии настоящие вибриссы сохраняются лишь у речных дельфинов, таких как гангский дельфин (Platanista gangetica) и амазонская иния (Inia geoffrensis). Эти китообразные живут в мутной речной воде и вынуждены ловить добычу со дна, а то и копаться в иле. Их вибриссы массивнее, чем у усатых китов, достигая в поперечнике 1 мм, а в длину 1,5-2 см. Они слегка сплющены и в отличие от вибрисс наземных животных до самого кончика почти не суживаются. Каждый волосок посредине согнут, и его кончик направлен в сторону кожи.
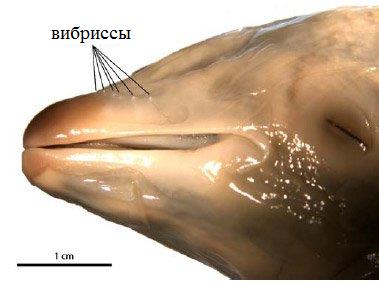
Вибриссы на морде эмбриона дельфина.
Между вибриссами и собственно волосами много существенных различий. Движение вибрисс обеспечивается поперечнополосатой мускулатурой, волос — гладкой (единственное исключение или, возможно, следствие параллельности развития — волосы однопроходных, которые снабжены поперечнополосатой мускулатурой). Соединительнотканная капсула у вибриссной сумки развита значительно сильнее, чем у волосяной. Вибриссы не имеют сопутствующих потовых желез, а их сальные железы развиты слабо. Различия между волосами и вибриссами касаются также характера кровоснабжения и иннервации, строения (у вибрисс отсутствует сердцевинный слой), характера линьки (вибриссы сменяются по одной по мере стирания, независимо от общей линьки), принципа функционирования (вибрисса как подвижный рычаг) и самого их функционального значения (чувствительного у вибрисс и теплоизолирующего у волос). Косвенными доказательствами возможного разделения путей развития волос и вибрисс служат также опережающее развитие иннервации еще не сформированной луковицы вибриссы в онтогенезе, возможность обратного развития вибрисс в рецепторы (как это происходит у дельфинов), глубокое залегание луковиц вибрисс в подкожной клетчатке и обособление их мышц от подкожной поперечнополосатой мускулатуры и, наконец, представительство каждой вибриссы в сенсорных зонах мозга.
Зверообразные являлись водными и полуводными формами с мягкой эластичной и, видимо, железистой кожей, которая в процессе их выхода на сушу ороговевала и становилась влагонепроницаемой. Железы опускались глубоко в кожу, изолируясь от защитных кератиновых структур, а последние превратились в волосы и чешуи. Предполагается, что некоторые цинодонты уже приобрели такие прогрессивные особенности млекопитающих, как теплокровность и выработка молока для питания детенышей. Развитие шерстного покрова при этом имело теплоизолирующее значение и было важным для становления гомойотермии. Впрочем, существует и противоположное мнение, согласно которому первоначальной функцией волосяного покрова была защита организма от перегрева. Помимо прочего, некоторые ископаемые остатки позволяют предполагать наличие на конце морды и по всей верхней губе ряда цинодонтов «протовибрисс». Высказывалась даже предположение, что волосы млекопитающих произошли именно от вибрисс, которыми было покрыто тело их зверообразных предков. Таким образом, волосовидные образования, вероятнее всего, уже имелись у наиболее прогрессивных цинодонтов или у древнейших млекопитающих, причем у ветви, давшей однопроходных, это были только волосы как таковые, а у ветви, приведшей к возникновению сумчатых и плацентарных, — и волосы и вибриссы. У современных однопроходных вибрисс нет (у них развиты кожные механо- и электрорецепторы), у сумчатых вибриссы присутствуют на морде и конечностях, а у плацентарных их распространение на теле может быть самым разнообразным.
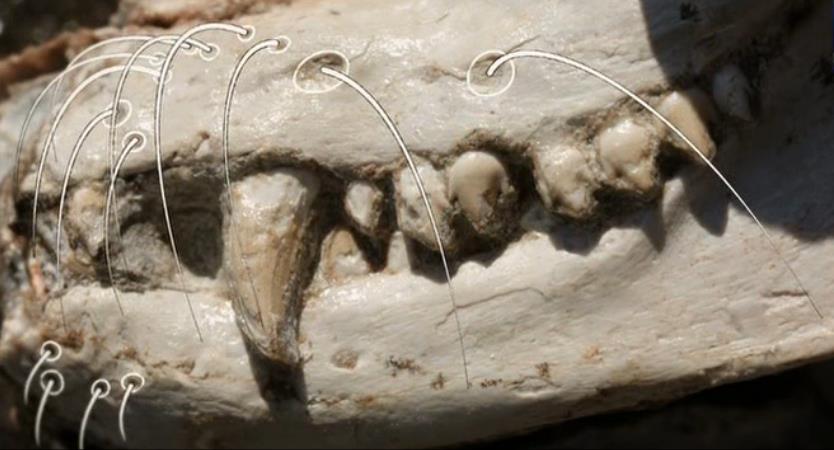
Ямочки на челюстях цинодонта тринаксодона (Thrinaxodon liorhinus) — точки отхождения «протовибрисс» и/или места прохождения сосудисто-нервных пучков, снабжавших губы.
Волосы — ткань, свойственная исключительно млекопитающим и не гомологичная каким-либо производным кожи других амниот. Существует предположение, что волосы произошли от роговых осязательных волосков чешуйчатых рептилий, или прототрихий (предшественников волос) бесхвостых амфибий, которые действительно очень напоминают эмбриональные волосы, или органов боковой линии личинок водных хвостатых амфибий, которые могли соответствующим образом видоизмениться в связи с выходом на сушу. Действительно, в покровах зародышей млекопитающих развитию чешуй и волос предшествует увеличение числа чешуевидных клеток, что иногда считают кратковременным эмбриональным повторением стадий развития органа предков. Известно заболевание «черный волосатый язык», при котором у человека нитевидные сосочки языка гипертрофируются и приобретают вид настоящих волос — т. е. в принципе такое видоизменение роговых структур возможно. Однако сейчас превалирует мнение, что волосы — новоприобретение филогенетической линии млекопитающих, эволюционно не связанное с какими-либо производными наружных покровов их предков.
Возможно, вибриссы развились у предковых форм млекопитающих из эмбриональных закладок примитивных механорецепторов, в то время как волосы — от чешуевидных эпидермальных закладок. Жесткий прямой стержень вибрисс и отсутствие у них характерного для волос ворса может свидетельствовать о том, что они появились первыми — у таких предковых форм, толстая шкура которых не была прочно прикреплена к туловищу. Волосы же возникли у более поздних форм, у которых, по мере оформления подкожных структур, кожа стала тоньше, но прочнее соединялась с туловищем, и растущие волосы приобрели наклон вследствие натягивания, растягивания и роста такой закрепленной кожи. Этот процесс прослеживается и в онтогенезе: первыми, на этапе еще окончательно не оформленных и не прикрепленных к подкожной мускулатуре слоев кожи, закладываются вибриссы, а волосы — позднее, в полностью сформированной, более тонкой и прикрепленной коже. У зрелорождающихся форм вибриссы в процессе индивидуального развития закладываются раньше, но развиваются медленнее и оформляются в более поздний эмбриональный период, чем у незрелорождающихся.
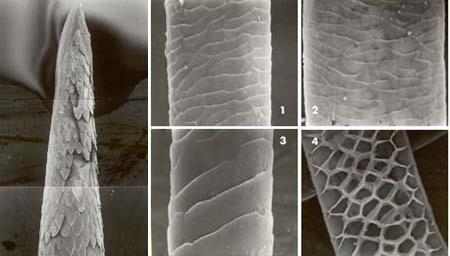
Слева — вершина иглы-гарпуна североамериканского дикобраза (Erethizon dorsatum).
Справа — структура волос пятнистого оленя (Cervus nippon): 1 — кутикула основания остевого волоса; 2 — кутикула гранны; 3 — пуховой волос; 4 — поперечный срез через сердцевину остевого волоса.
Весьма интересен процесс эволюционного становления игл. Их образование происходило в разных группах млекопитающих параллельно и в связи с конкретными приспособлениями к образу жизни. О том, что иглы — модификация волос, свидетельствуют многочисленные переходные формы (щетины, полуиглы). У эмбрионов ежа иглы закладываются позднее волос и образуются путем слияния нескольких луковиц, что довольно обычно у видов, у которых имеются пучки волос, растущие из общей луковицы. Однако образование иглы у ежа — не механическое объединение луковиц, а более сложный процесс. Каждая игла формируется в результате разной скорости развития группы луковиц, в которой центральная, самая крупная, луковица развивается быстрее других и, разрастаясь, «захватывает» более мелкие и медленнее растущие. В итоге игла оказывается состоящей из разделенных толстыми продольными перегородками тяжей сердцевинных клеток, среди которых центральный тяж наиболее крупный. У представителей совсем другой группы — дикобразов (Histricidae) — иглы устроены проще: перегородки гораздо тоньше, а сердцевинные клетки либо одинаковы и по форме и по размерам, либо немного крупнее в центральной части стержня. В бороздчатых иглах и полуиглах некоторых хомяков (Cricetidae) и мышей (Muridae) сердцевина сохраняется лишь в боковых частях иглы и разделена утолщенным корковым слоем, что предполагает формирование этих иголок из одной крупной закладки волоса без сердцевины и двух более мелких — с сердцевиной. Щетины свиней и пекари очень похожи на «комбинированные» иглы и также состоят из разделенных толстыми перегородками тяжей сердцевинных клеток. В иглах (или полуиглах) тенреков и ехидн продольные перегородки отсутствуют, сердцевинные клетки имеют сходные размеры и отличаются утолщенными стенками и слабым развитием полостей — т. е. эти структуры формируются из единой закладки.
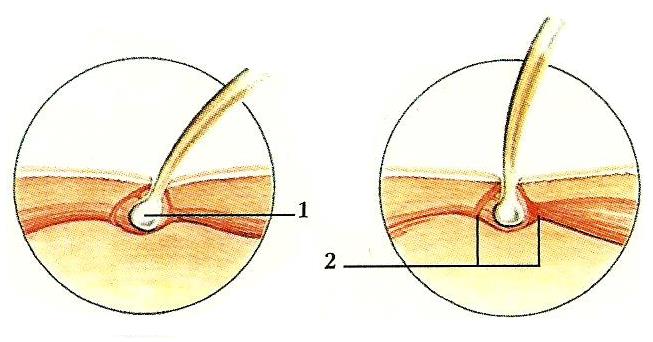
Каждая иголка обыкновенного ежа (Erinaceus europaeus) с одной стороны заострена, а с другой стороны имеет уплотнение, напоминающее волосяную луковицу (1). Луковица, находящаяся в коже, позволяет игле вращаться. При опасности еж ощетинивается, подняв иголки торчком, а в спокойном состоянии иглы прижаты к телу. За эти действия отвечает пара мышц-антагонистов (2).
У человека количество волос примерно соответствует таковому у человекообразных обезьян, но они гораздо тоньше и короче. Редукция волос в данном случае связывается со становлением бипедии (двуногости), фетализацией — замедлением темпов развития в онтогенезе отдельных органов и их частей, а также половым отбором — предпочтением половых партнеров с голой кожей и получения наслаждения при любовных играх. Возможно, роль важного фактора сыграл тепловой стресс, неизбежный при переходе от жизни в тенистых лесах к обитанию на открытых пространствах саванн. При формировании адаптивного типа конституции гоминид основным механизмом терморегуляции стало интенсивное потоотделение. Развитый волосяной покров затруднял отдачу тепла и был утерян. Замедленное развитие под гормональным контролем приводит к подавлению ряда признаков, в частности сильной обволошенности, так что с определенной точки зрения человека можно рассматривать как зародыша примата, достигшего половой зрелости (своеобразная неотения). Как и у человека, выношенные плоды гориллы (Gorilla gorilla) и шимпанзе (Pan troglodytes) имеют длинные волосы на голове, но слабо оволосненные лицо и туловище, а облысение у взрослых особей проходит сходно с таковым у человека.
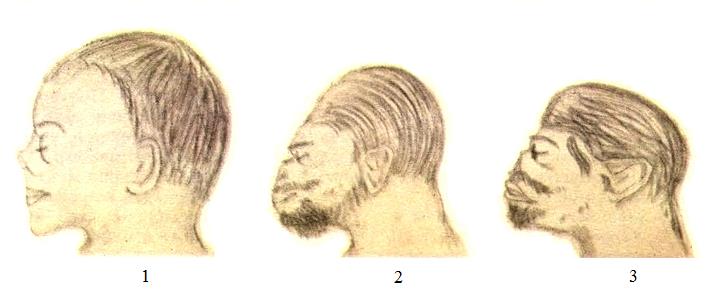
Головы зародышей человекообразных приматов: 1 — человек (Homo sapiens); 2 — горилла (Gorilla gorilla); 3 — шимпанзе (Pan troglodytes).
Эмбриональные волосы, или лануго, развиваются у человека раньше более грубых и крупных «терминальных» волос. Закладки эмбриональных волос появляются к концу 3-го месяца на лице и голове, затем они развиваются по всему телу сверху вниз, а начиная с 8-го месяца лануго постепенно утрачивается. Раньше считали, что волосы плода человека — рекапитуляция (повторение у эмбрионов признаков взрослых предков) волос человекообразных обезьян. Однако оказалось, что лануго имеется и у них, замещаясь впоследствии постоянным волосяным покровом. Показана также функциональная значимость эмбриональных волос. Их реакция в ответ на движение околоплодных вод (ворс направлен против него, и волосы действуют как рычаги с длинным плечом) усиливает деятельность кожных рецепторов и поддерживает мышечный тонус, обеспечивающий определенную позу плода. Утраченные и плавающие в амниотической жидкости волоски частично потребляются плодом через ротовое отверстие. Баластное вещество, содержащееся в них, служит для возбуждения перистальтики кишечника зародыша. Наконец, лануго создает предпосылки для фиксации сырообразной смазки, которая появится на коже плода позже. Она состоит из слущенных кожных клеток, секретов, эмбриональных волос и биологически является очень ценным материалом, который среди прочего защищает нежную кожу от агрессивных веществ околоплодных вод. Что же касается вибрисс, то они у человекообразных приматов полностью отсутствуют — нет даже их закладок. Функционально их частично замещают тилотрихии — чувствующие волоски, равномерно распределенные по всему телу.

Лануго 20-недельного зародыша человека (Homo sapiens).
Несмотря на все различия, как вибриссы, так и разные типы волос и их производных — иглы, полуиглы, колючки, полуколючки, щетина, кроющие, направляющие, остевые разных категорий, пуховые, тилотрихии, монотрихии, осметрихии и др. — обладают общей микроструктурой. Их стержень представляют собой ороговевший цилиндр различной конфигурации, покрытый одно- или многослойной кутикулой, заполненный плотными веретеновидными корковыми клетками и не всегда присутствующим центральным нежным сердцевинным слоем — ороговевшими остатками стенок сердцевинных клеток метаболически активной луковицы. Волосяная луковица является динамичной комплексной системой, подверженной возрастным и сезонным изменениям под гормональным контролем, с достаточно большой функциональной пластичностью, позволяющей изменять параметры волоса. Считается, что луковицы возникают в онтогенезе лишь единожды, однако экспериментально доказано новообразование волос при регенерации кожи. При онтогенезе волосяного покрова имеет место гетерохрония — разница во времени закладки и развития различных типов волос. Как уже говорилось, сначала формируются самые сложные структуры — вибриссы, затем чувствующие волосы — снабженные капиллярами и нервами тилотрихии, а еще позже — волосы разных типов — от сложных остевых (из первичных луковиц, снабжены мышцами и железами), полуигл, игл, щетин до просто устроенных пуховых (из самостоятельных закладок или из вторичных луковиц). Первичные луковицы происходят из эпидермиса, а вторичные являются продуктом почкования первичных.
вперед
в оглавление