в оглавление
назад
Зрение (продолжение)
Не весь свет, проникающий через зрачок и достигающий светочувствительной сетчатки, используется для зрения. Часть его поглощается внешним пигментным слоем. Для некоторых животных (преимущественно ночных) это означало бы слишком большие потери и так незначительного количества доступного света. Поэтому у таких видов позади сетчатки формируется отражательное дно глаза, или зеркальце (tapetum lucidum). Его функция — отражение неиспользованного света обратно на сетчатку для дополнительной стимуляции ее рецепторов. У млекопитающих встречается два основных типа зеркальца. Зеркальце волокнистого типа состоит из блестящих волокон соединительной ткани. Такое зеркальце характерно для копытных. Зеркальце клеточного типа слагается из нескольких слоев уплощенных эндотелиальных клеток, содержащих кристаллы гуанина. Такой тип обычен у хищных. Зеркальце обычно расположено в сосудистой оболочке за сетчаткой, но, например, у некоторых летучих мышей и у виргинского опоссума (Didelphis virginiana) оно погружено в саму сетчатку. Именно зеркальце благодаря отражению хотя бы минимального количества света при почти полной темноте обуславливает кажущееся свечение глаз. Подобное «свечение» глаз характерно для многих млекопитающих — например, хищных, некоторых копытных и приматов, но у человека встречается лишь как атавизм. Цвет, которым «светятся» глаза, зависит от количества крови в капиллярах сосудистой оболочки и содержания родопсина (пурпурного светочувствительного пигмента) в палочковидных элементах сетчатки, через которую проходит отраженный свет.

Эффект «свечения» глаз у кошек (Felis silvestris).
Установлено, что за цветовое зрение у позвоночных отвечают 4 семейства генов, кодирующих опсины колбочек: SWS1, SWS2, Rh2, LWS. Все 4 семейства генов были выявлены у птиц, рыб и рептилий, у амфибий — лишь 3. У млекопитающих ситуация значительно сложнее. У однопроходных выявлены гены семейств SWS2 и LWS, а также нефункциональный ген из SWS1; у сумчатых имеются гены из SWS1 и LWS, а также, возможно, из Rh2. Плацентарные имеют гены опсинов только из семейств SWS1 и LWS. Вместе с тем, млекопитающие хорошо распознают особенности формы и рисунка предметов или их частей, а также разнообразные движения. В наибольшей степени эти способности свойственны обезьянам.
У многих рептилий и птиц присутствует 4 типа колбочек, обеспечивающих четырехкомпонентное цветовое зрение. Кроме того, в колбочках этих животных содержатся окрашенные капельки жира, действующие как светофильтры и в сочетании с фотопигментами определяющие спектральную чувствительность рецепторов. В колбочках млекопитающих подобные светофильтры отсутствуют, вследствие чего их способность к цветовому зрению основывается только на избирательной чувствительности фотопигментов. Однако, обладая только 2 типами колбочек, большинство млекопитающих способно исключительно к двухкомпонентному зрению. Таковы, в частности, многие копытные, хищные и грызуны. При этом дифференциация цветов у них очень ограничена. Например, рыжая полевка (Myodes glareolus) различает лишь красный и желтый цвета, домашний бык (Bos primigenius) — синий и красновато-зеленый, кошка (Felis silvestris) — голубой, зеленый и желтый.
Слабое восприятие цветов связано с тем, что каждая длина волны стимулирует оба типа колбочек, но в разной степени и в соответствии с их относительной чувствительностью в этой части спектра. Если мозг может распознавать такую разницу, животное различает длину волны света по его интенсивности. Однако эти определенные отношение возбудимости характерны более чем для одной части спектра, поэтому некоторые длины волн воспринимаются одинаково. Длина волны, одинаково возбуждающая оба типа колбочек (в области пересечения кривых поглощения), воспринимается как белый цвет и называется «нейтральной точкой» спектра. В то же время, млекопитающие различают большое количество оттенков серого цвета: например, кошка — до 25. Это вполне закономерно, ведь их предки были ночными животными с преобладанием палочек в сетчатке.
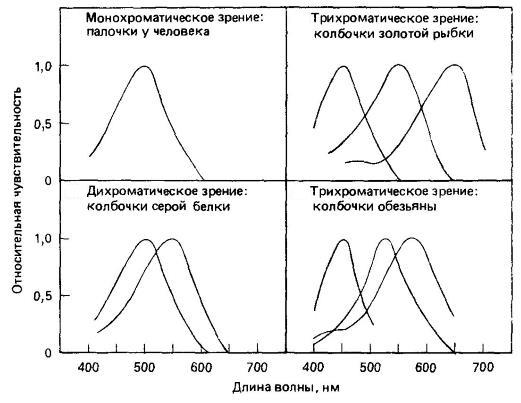
Типичные рецепторные механизмы при разных типах цветового зрения.
Цветовое смешение меньше выражено в зрительных системах узконосых и части широконосых обезьян, обладающих 3 фотопигментами. Однако некоторое смешение происходит и здесь: можно, например, вызвать впечатление любого цвета посредством разных сочетаний трех цветовых составляющих, специально подобранных по интенсивности и насыщенности. Без этого было бы невозможно зрительное восприятие цветной фотографии и цветного телевидения. Трехкомпонентному цветовому зрению сухоносых приматов сопутствует слабое сумеречное зрение в связи с небольшим числом палочек. Помимо обезьян, трехкомпонентное зрение среди млекопитающих присуще лишь некоторым сумчатым.
Что же касается мокроносых приматов, то цветовое зрение у них вообще отсутствует, поскольку это выражено ночные животные, воспринимающие свет лишь с помощью палочек. Не обнаружено цветовое зрение у виргинского опоссума (Didelphis virginiana), лесного хорька (Mustela putorius) и ряда других видов. Некоторые сумчатые, рукокрылые и грызуны способны видеть в ультрафиолетовом диапазоне. Со слабой цветовой чувствительностью в некоторой мере связана не слишком разнообразная (по сравнению с птицами) расцветка млекопитающих. Исключение в этом отношении составляют сухоносые приматы, в окраске которых как раз и встречаются яркие цвета — красный, желтый, зеленый, голубой.
Ганглионарные клетки внутренней поверхности сетчатки дают длинные нервные волокна, идущие в передний мозг. В месте их выхода не могут располагаться палочки или колбочки, чем и объясняется наличие здесь слепого пятна. У человека мозг кое-как заполняет пробел в изображении, используя информацию, получаемую от соседних участков. Хотя соединение сетчатки с мозгом и называют зрительным нервом, он отличается от любого нормального нерва в двух отношениях. Здесь, примерно как в обонятельном нерве, волокна, идущие к мозгу, принадлежат клеткам органа чувств, а не мозга. Кроме того, поскольку сетчатку эмбриологически правильнее рассматривать как часть самого мозга, зрительный «нерв» в действительности не является настоящим периферическим нервом, а скорее волокнистым трактом, соединяющим два отдела мозга.
Достигнув дна переднего мозга, волокна зрительного нерва входят в Х-образный зрительный перекрест (chiasma opticum). В мозге большинство аксонов ганглионарных клеток приходят в латеральные коленчатые тела таламуса, откуда идут к первичной зрительной коре. Из первичной зрительной коры сигнал передается во вторичные участки зрительной коры, некоторые из которых располагаются в височной и теменной долях. Аксоны зрительного нерва также проецируются на подкорковые ганглии мозга, минуя латеральные коленчатые тела: на предкрышечное поле, регулирующее диаметр зрачка, верхние бугры четверохолмия, участвующие в глазодвигательной функции, и супрахиазматическое ядро гипоталамуса, отвечающее за циркадные ритмы. При этом области мозга, получающие сигналы от сетчатки, если не всегда, то в большинстве случаев так упорядочены топографически, что создают мысленную «картину», воспроизводящую такое же расположение объектов, какое воспринимает сетчатка. Таким образом, зрительная информация по точкам проецируется на мозговые структуры, где и происходит обработка характеристик изображения (цвета, формы, движения, глубины и др.), причем для целостного восприятия эти свойства должны быть интегрированы. В то время как зрительные центры в коре больших полушарий млекопитающих устроены сложнее, чем у других позвоночных, значение зрительной коры их среднего мозга уменьшено.
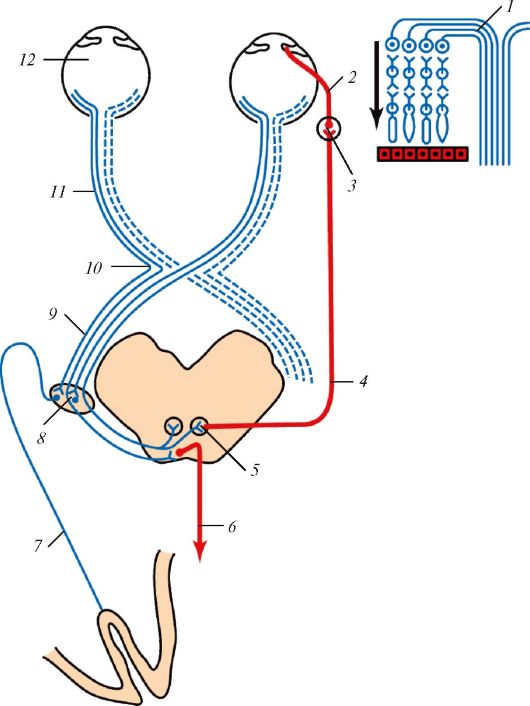
Проводящий путь зрительного анализатора млекопитающего с сильно развитым стереоскопическим зрением: 1 — схема строения сетчатки и формирования зрительного нерва (стрелка показывает направление света в сетчатке); 2 — короткие ресничные нервы; 3 — ресничный узел; 4 — глазодвигательный нерв; 5 — ядро глазодвигательного нерва; 6 — покрышечно-спинномозговой путь; 7 — зрительная лучистость; 8 — латеральное коленчатое тело; 9 — зрительный тракт; 10 — зрительный перекрест; 11 — зрительный нерв; 12 — глазное яблоко. Волокна из одноименных половин обоих глаз направляются в одноименную же половину мозга.
Как правило, снаружи глаз защищен 2 подвижными непрозрачными веками (palpebrae), из которых лучше развито верхнее. Веки часто снабжены ресницами, которые препятствуют засорению глаза. Внутренняя сторона век выстлана слизистой оболочкой — конъюнктивой. Зачастую здесь располагаются тарзальные, или мейбомиевы, железы (glandulae tarsales), выделяющие глазную смазку. Кольцо из волокон лицевой мускулатуры действует в качестве сфинктера, закрывающего веки. От прозрачной мигательной перепонки (membrana nictitans) у большинства млекопитающих сохранились лишь рудиментарные остатки во внутреннем углу глаза, но у некоторых из них (у кошек (Felidae), белого медведя (Ursus maritimus), ластоногих (Pinnipedia), верблюдов (Camelidae), трубкозуба (Orycteropus afer)) она вполне развита. Кроме того, у внутреннего угла глаза иногда помещается гардерова железа (glandula nictitans), выделяющая жирный смазывающий секрет (ее нет у приматов). В наружном углу глаза млекопитающих находится слезная железа (glandula lacrimalis), жидкие выделения которой промывают и предохраняют от высыхания глаз и внутреннюю поверхность век. Кроме того, в слезах содержится бактерицидный белок лизоцим. Носослезной проток (ductus nasolacrimalis), начинающийся во внутреннем углу глаза, отводит избыток жидкости в носовую полость. Таким образом, дополнительное значение слезной жидкости состоит в том, что она смачивает слизистую носа. Окологлазные железы наряду с веками и мышцами составляют вспомогательный аппарат глаза.
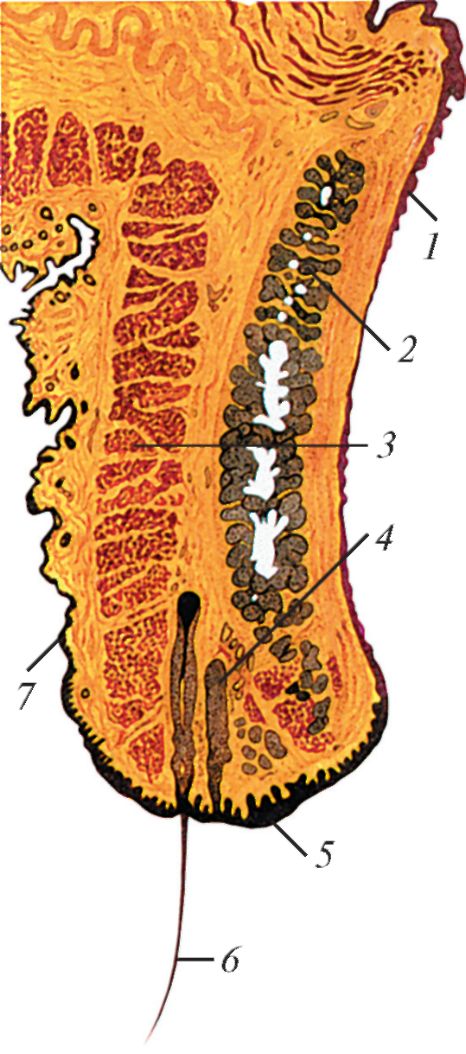
Строение века млекопитающего, фронтальный разрез: 1 — конъюнктива; 2 — хрящ века; 3 — вековая часть круговой мышцы глаза; 4 — ресничная железа; 5 — край века; 6 — ресница; 7 — кожа.
Острота зрения зависит от различных причин, но одним из главных определяющих ее факторов является величина глаз. Крупный глаз различает в осматриваемой картине больше подробностей не только потому, что эта картина в нем меньше изменена (линейное уменьшение картины в глазе кролика (Oryctolagus cuniculus) равно 112, человека (Homo sapiens) — 60, льва (Panthera leo) — 40), а и потому, что он отражается в большем числе зрительных клеток. И все-таки, глаза большинства млекопитающих относительно невелики. В частности, у человека они составляют около 1% от общей массы головы, тогда как у скворца этот показатель достигает 15 %. При этом млекопитающие малого размера имеют относительно большие глаза в отличие от крупных зверей, особенно если они родственны друг другу (например, кошка (Felis silvestris) и тигр (Panthera tigris)). Этого и следует ожидать, так как если глаз определенного размера обеспечивает удовлетворительное зрение для данного животного, то его укрупнение не даст преимущества в борьбе за выживание, а работа глаза никак не зависит от величины животного, которому он принадлежит.
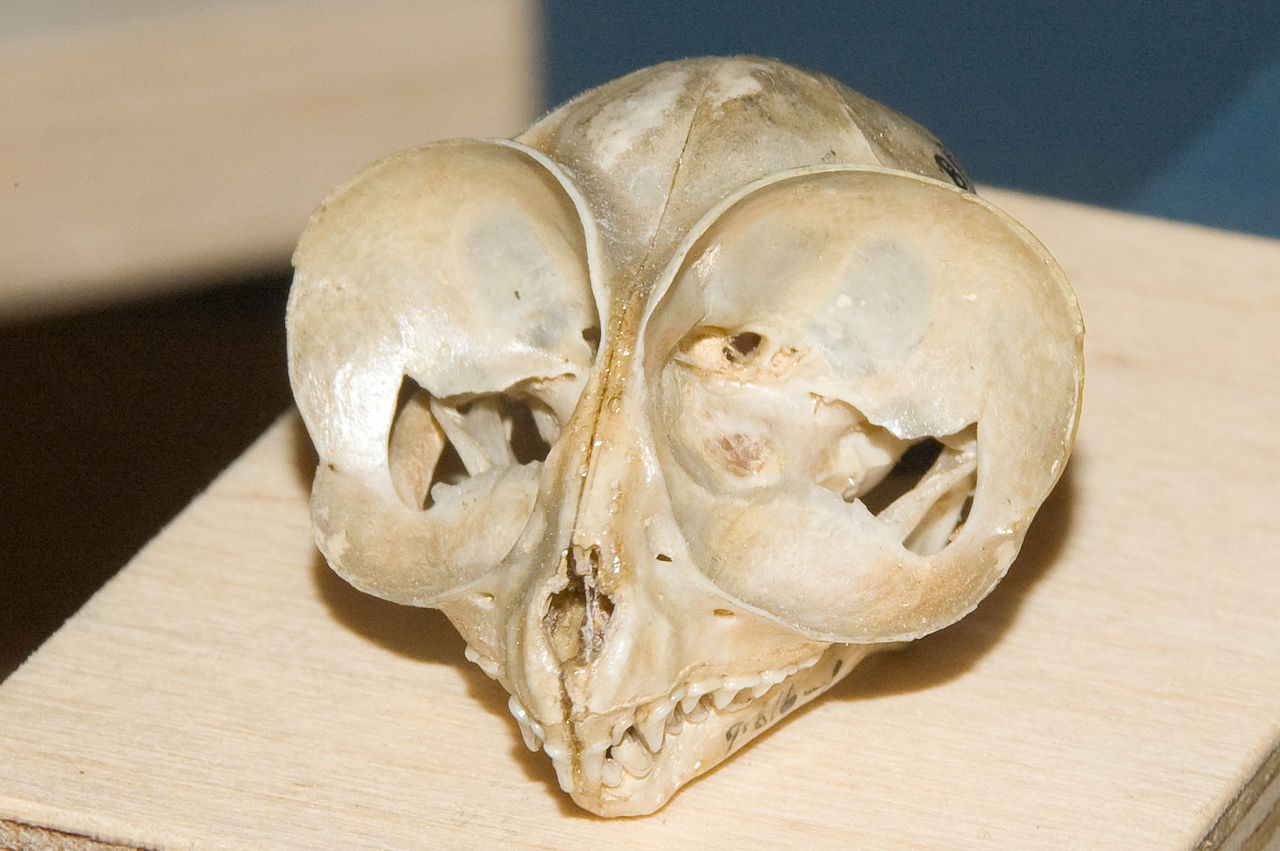
На черепе долгопята (Tarsius sp.) внимание к себе прежде всего привлекают огромные глазницы.
Хорошо развиты глаза зверей, ведущих преимущественно дневной образ жизни и населяющих открытые ландшафты (например, многих копытных); большая часть воспринимаемой ими информации поступает именно через зрительный канал. Значение зрения уменьшается у обитателей лесов, кустарниковых зарослей или травянистого покрова. Особенно крупных размеров достигают глаза млекопитающих с сумеречной или ночной активностью, для которых важен визуальный контроль — некоторых приматов (кошачьего лмура (Lemur catta), тонких лори (Loris), долгопятов (Tarsiidae), ночных обезьян (Aotus)), кошек (барханного кота (Felis margarita), манула (Otocolobus manul)) и т. п. Глаза ночных животных улавливают больше света благодаря широким зрачкам и большим хрусталикам; данных о повышенной чувствительности таких глаз к длинным волнам нет. У некоторых животных, например у галаго (Galago), череп сужен с боков, что приводит к цилиндрическому удлинению глаза.
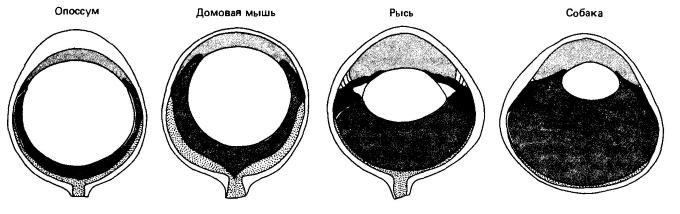
Сравнение глаз ночных млекопитающих — опоссума (Didelphis virginiana), мыши (Mus musculus) и рыси (Lynx lynx), — а также собаки (Canis lupus), обладающей дневным и ночным зрением.
У других ночных форм (например, у летучих мышей) глаза небольшие; в этом случае недостаток зрения компенсируется высокоразвитым слухом, обонянием и осязанием. У многих норников глаза в большей или меньшей степени редуцированы и регистрируют лишь изменения освещенности (у гоферов (Geomyidae), цокоров (Myospalax), слепушонок (Ellobius), прометеевой полевки (Prometheomys schaposchnikovi)). Иногда рудиментарные глаза полностью перестают функционировать и затягиваются кожей (у сумчатых кротов (Notoryctes), златокротов (Chrysochloridae), слепого крота (Talpa соеса), слепышей (Spalacinae)).
Глаза водных млекопитающих используются только для ближней ориентации, по своей выпуклости и большому показателю преломления они напоминают глаза рыб. Роговица у таких глаз уплощена, а хрусталик круглый, что свидетельствует о близорукости; слезные железы имеются, но выделяют жирный, а не водянистый секрет. Некоторые китообразные специально адаптированы к господствующим на глубине условиям освещения. Например, у глубоко ныряющего северного плавуна (Berardius bairdi) зрительные пигменты поглощают короткие волны сильнее, чем у неглубоко ныряющего серого кита (Eschrichtius gibbosus).
Поле зрения во многом зависит от положения глаз на голове. При бинокулярном, или стереоскопическом, зрении получаемые от обоих глаз картинки в большей или меньшей степени перекрываются, и два изображения, передаваемые в мозг, приблизительно одинаковы. Такое зрение обеспечивает гораздо более точную оценку расстояния, чем монокулярное. У большинства млекопитающих глаза расположены по бокам головы — это обеспечивает почти круговой обзор, при котором бинокулярное зрение ограничено лишь узким сектором прямо перед мордой. Реже глаза развернуты вперед; общий обзор при этом сокращается, но поле бинокулярного зрения расширяется. Первый тип преобладает у копытных и грызунов, постоянно ожидающих нападения врагов. Второй тип характерен для приматов, которым необходимо точно определять расстояния при прыжках с ветки на ветку, и для части хищников, особенно кошачьих, которые, нападая из засады, должны точно фиксировать расстояние до жертвы.
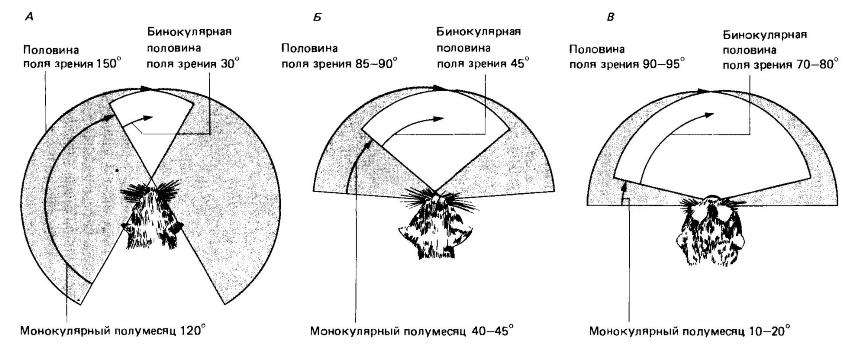
Поля зрения: А — у белки (Sciurus sp.); Б — у кошки (Felis sp.); В — у ночной обезьяны (Aotus sp.).
Важной анатомической особенностью, связанной с бинокулярностью зрения, является неполная декуссация в зрительном перекресте. У многих млекопитающих волокна от тех областей двух сетчаток, которые воспринимают одинаковые фрагменты внешней картины, направляются к одной и той же половине мозга. Таким образом, определенные группы волокон не переходят на другую сторону (т. е. полной декуссации не происходит), а меняют свое направление в зрительном перекресте под прямым углом и сопровождают соответствующие волокна из противоположного глаза. Например, у человека, где перекрывание полей зрения почти полное, практически все волокна от левых половин сетчаток направляются в левую половину мозга, а от правых половин сетчаток — в правую половину мозга. В результате зрительная область каждой из сторон мозга воспринимает половину всего поля зрения в виде «двойной экспозиции» (поскольку хрусталик проецирует на сетчатку перевернутое изображение, левая половина единого поля зрения обрабатывается у человека в правой половине мозга и наоборот). Благодаря дальнейшим сложным взаимодействиям между полушариями две половины картины совмещаются и осознаются как единое стереоскопическое изображение.
При рассматривании предмета, когда важна острота зрения, изображение приводится в фокус на центральной ямке — части сетчатки, которая содержит только колбочки и обеспечивает наибольшую остроту зрения. Человек (Homo sapiens) обладает одной расположенной в центре глаза ямкой с круглыми очертаниями. У гепарда (Acinonyx jubatus) и ряда других обитателей открытых местностей центральная ямка вытянута по горизонтали. У древесных млекопитающих, например у белки (Sciurus vulgaris), центральная ямка имеет форму диска; то же самое относится к сумеречным и ночным формам, например к ежу (Erinaceus europaeus), кошке (Felis silvestris) и мыши (Mus musculus). Для таких животных вертикальное направление, очевидно, не так важно, как и горизонтальное. У лошади (Equus ferus) нет центральной ямки, но есть «центральная линия». Это центральная область на сетчатке, относительно которой перпендикулярно глазному дну выстраиваются в линию рецепторы. Направление светового потока на центральную линию обеспечивает фокусировку изображения у лошади.

В связи с обитанием в мутной воде глаза гангского дельфина (Platanista gangetica) утратили хрусталик, их зрительный нерв деградировал, а слизистая оболочка начала выполнять осязательную функцию. Животное практически слепо, хотя по-прежнему способно улавливать интенсивность и направление света. Ориентируется и охотится гангский дельфин с помощью развитой эхолокации.
Небольшие различия в углах зрения левого и правого глаз позволяют воспринимать глубину и трехмерность пространства — ощущения, которые иначе достигнуты быть не могут. Для одновременного фокусирования глаз должна существовать некоторая конвергенция обеих линий взора. Чем ближе рассматриваемый предмет, тем большая необходима конвергенция. Направление обеих линий взора устанавливают наружные мышцы глаза, пока оба изображения на сетчатках не совпадут и головной мозг не будет регистрировать единое изображение. Если в это же время мозг отмечает степень конвергенции обоих глаз, возникает информация о расстоянии до предмета. Однако точное совпадение обоих изображений близких предметов на сетчатках невозможно. Расстояние между глазами будет определять разницу в положении двух изображений. Это расхождение (диспаратность) изображений на сетчатках тоже дает важную информацию о расстоянии до предметов. Оценка расстояния и глубины — сложный процесс, для которого требуется много данных помимо тех, какие доставляют конвергенция и диспаратность.
Высокий уровень организации зрительной системы открывает перед млекопитающими возможности не только для совершенной визуальной ориентации в пространстве, но и для усложнения и обогащения зрительных связей между особями. Возникли и широко используются «языки» формы, поз, жестов и мимики, служащие упорядочению отношений в популяциях и образованию группировок с согласованным поведением сочленов.
вперед
в оглавление