Состав
Agriotherium hendeyi Jiangzuo et Flynn, 2019 — вид, известный из позднего миоцена (конец хемфильского века) США (штат Аризона).
Установлен по нижнечелюстному фрагменту с p4-m2 и альвеолами p1-p3, происходящему из позднехемфильского слоя (около 6 млн. лет назад) формации Оуибурис в Аризоне.

Зубочелюстной материал агриотерия (Agriotherium hendeyi) из штата Аризона, США.
Размеры относительно небольшие. Тело ветви нижней челюсти сужено.
Agriotherium inexpetans Qiu, Xie et Defa, 1991 — вид, известный из позднего миоцена - ? раннего плиоцена (верхи тортонского - верхи мессенского яруса / ? низы занклского яруса) Китая (провинция Ганьсу).
Установлен по щечным зубам (левый р4, талониды левого и правого m1, левый и правый m2, левый m3 от одной особи) из позднемиоценовых отложений формации Баодэ (около 9 млн. лет, что соответствует зоне MN 13 европейской шкалы) местонахожденя Цзегоу в провинции Ганьсу. Это древнейшие известные остатки Agriotherium в Азии. Для остатков из Сяосянь предполагается раннеплиоценовый возраст (? MN 15).
Размеры зубов небольшие. Метаконид m1 высокий, сближен с протоконидом; энтоконид редуцирован в размере, отделен от более высокого метаконида широкой щелью; бугорок позади энтоконида не выражен; талонид открыт сзади. m2 длинный и узкий, его тригонид не шире, чем талонид, с выраженным параконидом; протоконид выше метаконида. Принадлежность материала к роду Agriotherium, по мнению авторов описания, определяется тем, что гипоконид m1 выше, чем энтоконид (у Indarctos соотношение высоты этих бугорков обратное). Для образца из Сяосяня характерно слабое развитие парастиля на Р4.
Agriotherium myanmarensis Ogino, Egi, Zin-Maung-Maung-Thein, Thaung-Htike et Takai, 2011 — вид, известный из позднего миоцена или раннего плиоцена (верхи мессинского / занклский ярус) Мьянмы.
Установлен по нижнечелюстному фрагменту, извлеченному из позднемиоценовых или раннеплиоценовых речных наносов местонахождения Чайнгзаук в центральной части Мьянмы. Оттуда же происходит еще один фрагмент нижней челюсти.
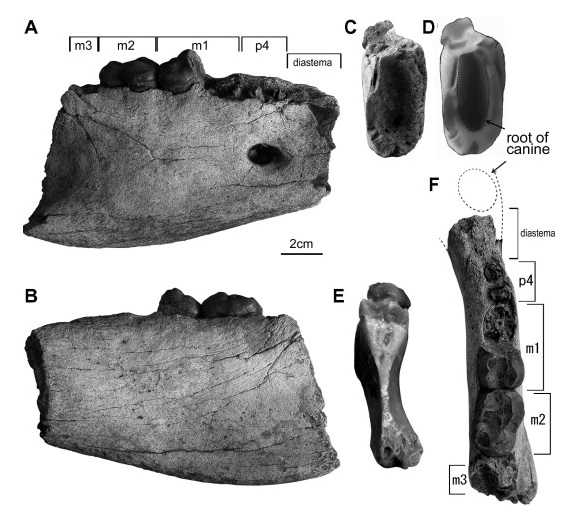
Нижнечелюстной фрагмент агриотерия (Agriotherium myanmarensis) из Чайнгзаука, Мьянма.
Нижняя граница тела нижней челюсти прямая. Талонид m1 достаточно редуцирован. Метаконид m1 крупнее гребня энтоконид-энтоконулид. Диастема между клыком и p4 очень короткая. Щечные зубы небольшие. Морфологически данный вид больше напоминает европейского Agriotherium insigne, а не восточно- и южноазиатских Agriotherium inexpetans, Agriotherium palaeindicus и Agriotherium sivalensis.
Agriotherium palaeindicum (Lydekker, 1878) — вид, известный из позднего миоцена (верхи тортонского - мессенский ярус) Пакистана (провинция Пенджаб). Синоним: Hyaenarctos palaeindicus Lydekker, 1878.
Установлен по левой верхнечелюстной кости с Р4-М2, происходящей из средних сиваликских слоев Джаби возле Хаснота. Ввиду незначительных морфологических отличий долгое время рассматривался в качестве мелких особей гораздо более позднего Agriotherium sivalensis.
Agriotherium africanum Hendey, 1972 — вид, известный из позднего миоцена - раннего плиоцена (мессинский - занклский ярусы) ЮАР, Уганды и Кении, а также, возможно, Эфиопии и Ливии. Синонимы: Agriotherium aecuatorialis Morales, Pickford et Soria, 2005; Agriotherium sivalensis africanum Baryshnikov, 2007.
Установлен по фрагменту левой верхнечелюстной кости с Р4 из отложений формации Варсватер местонахождения Лангебаанвег в ЮАР. Возраст слоев определяют в 4,9-4,8 млн. лет, т. е. как ранний плиоцен (аналог MN 14). Помимо типового образца, найдены другие остатки. Изолированный m2 из формации Нкондо в Уганде относится к концу миоцена. Вместе с материалом из раннеплиоценовых слоев формации Мабагет местонахождений Нгетабквоны и Чепримок в Кении находка из Уганды была описана как вид Agriotherium aecuatorialis. Отличия его от южноафриканского агриотерия незначительны (протокон Р4 более слабый и простой). Agriotherium sp. определен из позднемиоценовых или раннеплиоценовых (9-4 млн. лет назад) слоев формации Аду-Аса в среднем течении реки Аваш, Эфиопия. Также Agriotherium sp. установлен из конца позднего миоцена (мессинский ярус) отложений формации Сахаби в Ливии.
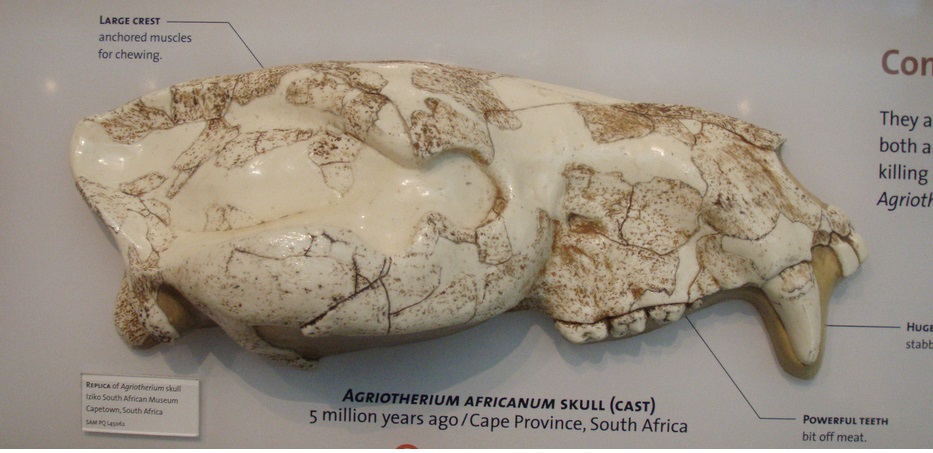

Неполные череп и нижняя челюсть агриотерия (Agriotherium africanum) из Лангебаанвега в Западной Капской провинции, ЮАР.
Размеры черепа крупные, кондилобазальная длина 2 черепов из Лангебаанвега составляет соответственно 42 и 46,5 см. Длина m1 варьирует в пределах 37,8-44 мм. Значительный разброс в величине зубов и костей из типового местонахождения свидетельствует о наличии размерного полового диморфизма. Также наблюдается большая вариабельность в строении щечных зубов, например в форме коронки М2, в степени развития метаконида m1 и т. д. Парастиль на Р4 хорошо развит.
Agriotherium insigne Kurten, 1968 — вид, известный из раннего - позднего плиоцена (занклский - пьяченцский ярусы) Испании, Франции, Великобритании, Болгарии, Польши, а также, возможно, Украины, Молдавии, России и Таджикистана. Синонимы: Ursus minutus Gervais, 1852; Hyaenarctos insignis Gervais, 1853; Agriotherium minutum Gervais, 1869; Agriotherium intermedium Stach, 1957; Agriotherium sivalensis minutum Baryshnikov, 2007.
Таксон Hyaenarctos insignis был установлен по фрагменту правой верхнечелюстной кости с Р4-М2 из раннего русциния (MN 14) Монпелье во Франции. Позднее отсюда же (Бутоне) были описаны нижние щечные зубы. Немного раньше из Монпелье по очень маленькому изолированному m3 с одним корнем был описан Ursus minutus. Поскольку оба таксона происходят из одного местонахождения и метрически сходны, они являются конспецифичными. Так как видовой эпитет minutus имеет приоритет, его было бы правильнее использовать для европейских находок. Они, в частности, были сделаны во Франции (Виалетт, MN16), Испании (Алькой, MN 14) и Англии (Красные краги). У образца из Монпелье длина ряда р4-m1 составляет 56,9 мм, ряда m1-m2 — 70,5 мм. По сравнению с Agriotherium sivalensis, парастиль на Р4 развит слабо, а коронка М2 несколько шире.
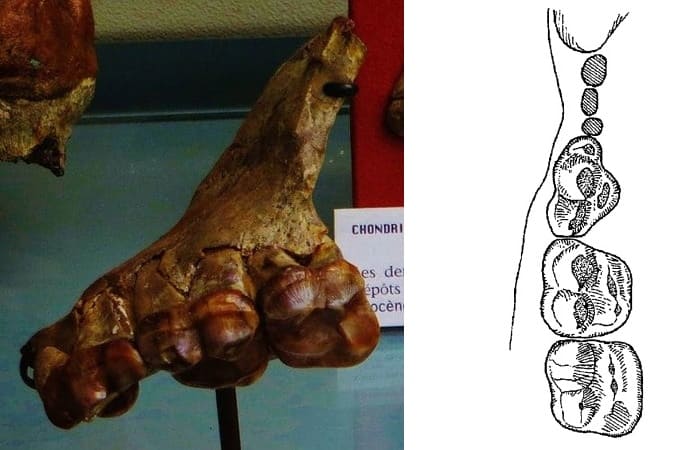
Типовой фрагмент правой верхнечелюстной кости и прорисовка жевательной поверхности правого ряда m1-m2 агриотерия (Agriotherium insigne) из Монпелье, Франция.
Еще один таксон, Agriotherium intermedium, был описан из позднего русциния (MN 15) Веже в Польше по фрагменту правой верхнечелюстной кости с Р4-М2, части левой ветви нижней челюсти с р4 и разрозненным зубам, скорее всего, принадлежавшим одной особи. Промеры типового материала: M1 — длина 25 мм, ширина 25 мм; М2 — длина 24,5 мм, ширина 25 мм; m1 — длина 37 мм, ширина 20,5 мм. Размеры несколько меньше, чем у Agriotherium sivalensis. Морфологические отличия сводятся к следующему: на M1 длина паракона равна длине метакона (у Agriotherium sivalensis первый бугорок отчетливо крупнее); передняя граница коронки М2 почти параллельна задней границе (у Agriotherium sivalensis они расположены по отношению друг к другу под острым углом); задний край М2 почти перпендикулярен язычному краю, с незначительной выемкой посередине (у Agriotherium sivalensis задний край зуба широко округлый и демонстрирует глубокую среднюю выемку); на m1 имеется крупный энтоконид и маленький бугорок, расположенный позади него, протоконид и метаконид разделены узкой щелью.
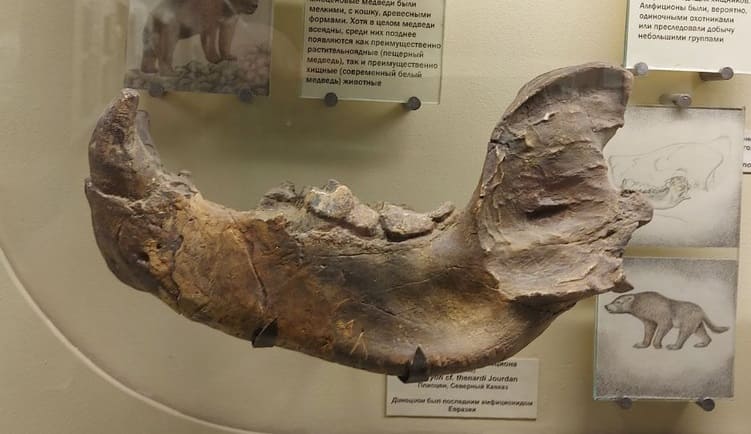
Левая ветвь нижней челюсти агриотерия (Agriotherium sp., = Dinocyon cf. thenardi) из села Косякино в Дагестане, Россия.
На территории бывшего СССР остатки, определенные в качестве Agriotherium spp., происходят из села Косякино на Северном Кавказе, села Кучурган и города Одесса в Украине и, возможно, и села Лучешть в Молдавии. К ним, вероятно, можно добавить материал из Джетымтау в Таджикистане. Все эти находки не были детально изучены и относятся к Agriotherium insignes лишь условно. Местонахождение Кучурган датируют ранним русцинием (MN 14), местонахождения Одесские катакомбы и Косякинский карьер — поздним русцинием (MN 15).

Нижнечелюстные фрагменты агриотерия (Agriotherium sp.) из села Кучурган в Одесской области, Украина.
Agriotherium sivalensis (Cautley et Falconer, 1836) Wagner, 1837 — типовой вид, известный из раннего плейстоцена (гелазский ярус) Пакистана (провинция Пенджаб). Синонимы: Ursus sivalensis Falconer et Cautley, 1836 (тип); Hyaenarctos sivalensis Falconer et Cautley, 1845; Agriotherium sivalense sivalense Baryshnikov, 2007.
В 1836 г. Кэтли и Фальконер описали новый таксон Ursus sivalensis из верхних сиваликских (пинджорских) слоев по остаткам, включавшим в себя почти полный череп, фрагмент правой ветви нижней челюсти с р4-m2 и кости посткраниального скелета, которые, вероятно, принадлежали нескольким особям. Название Agriotherium впервые было опубликовано в анонимной заметке за 1837 г., рассказывающей об открытии медведя в сиваликской фауне. Автором этого родового таксона общепринято признавать А. Вагнера.

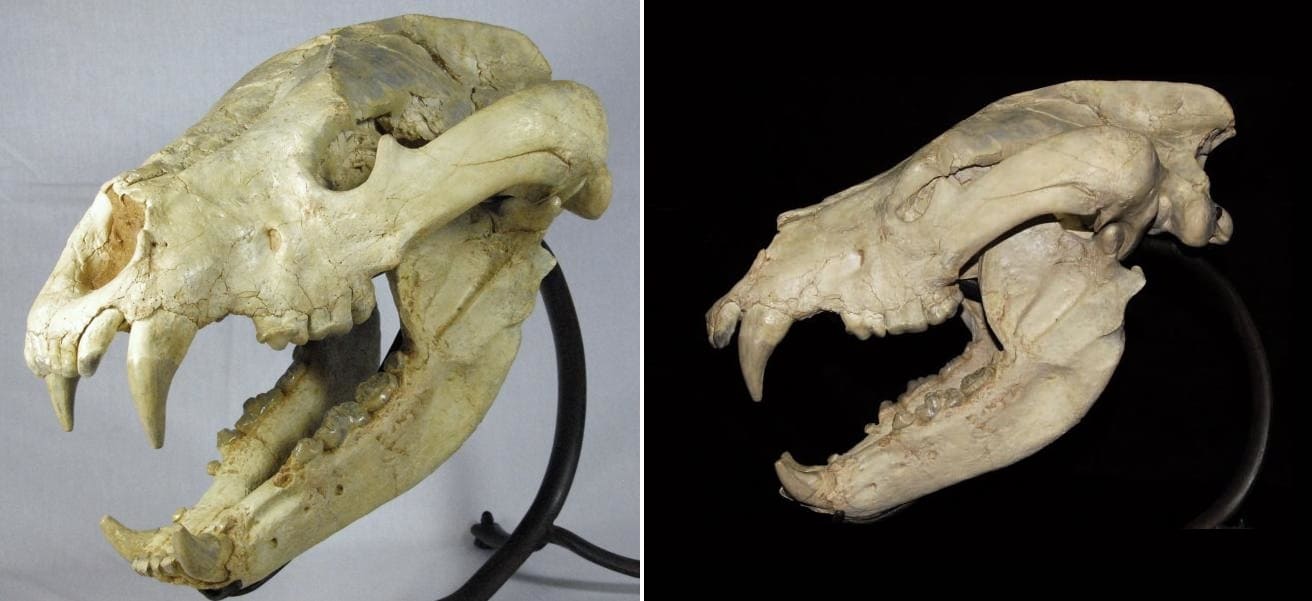
Типовой череп агриотерия (Agriotherium sivalensis) с Сиваликских холмов в провинции Пенджаб, Пакистан.
По величине Agriotherium sivalensis не уступал крупнейшим современным бурым медведям: реконструированная общая длина его типового черепа оценивается в 48,3 см. Лицевой отдел черепа короткий, расстояние от переднего края глазницы до основания резца резца I2 у типового образца составляет около 18,2 см. Глазницы широко расставлены, межглазничная ширина достигает 12,2 см. Глазница расположена над передней частью коронки М2. Подглазничное отверстие двойное, лежит на уровне M1. Надглазничные отростки небольшие, сагиттальный гребень сильно развит.
Клыки мощные и массивные, особенно верхние. Р2-3/р2-3 однокорневые. Р4 с с большим парастилем, коротким параконом, массивным и удлиненным метаконом. Для паракона характерны острые передний и задний кили, а также внутренний киль, спускающийся с вершины к переднему краю внутреннего выступа. Метакон образует режущий гребень, за ним нет метастиля. Внутренний выступ зуба длинный, массивный, несет крупный заостренный протокон, который расположен впереди относительно хищнической щели и имеет острые переднюю и заднюю грани. Перед ним может находиться маленький дополнительный бугорок (параконуль?). Основание протокона обрамлено краевым валиком внутреннего выступа.
Коронка М1 по форме близка к квадратной. Паракон и метакон почти равной величины, у их основания проходит четкий щечной цингулюм. Парастиля и метастиля нет, но на месте последнего цингулюм образует вздутие. Внутренняя доля зуба крупная, бассейн тригона имеет вид продольной долинки, расширяющейся кзади, которая не имеет складочек или бугорков и ограничена спереди препротокристой. Долинку пересекает поперечный киль, спускающийся с вершины метакона. На постпротокристе различим метаконуль, слабо дифференцированный от протокона. Переднеязычный угол коронки хорошо выражен. Внутренняя стенка зубной коронки крутая, без язычного цингулюма.
М2 похож по строению на M1, но с более длинным язычным краем. Паракон заметно крупнее метакона, оба бугорка одновершинные; парастиля нет. Протокон сравнительно крупный, одиночный или рассеченный поперечной щелью; метаконуль массивный; оба бугорка смещены в глубь коронки и обрамлены с лингвальной стороны краевым возвышением. От метакона к метаконулю идет поперечный гребень. По основанию протокона тянется язычный цингулюм, который постепенно поднимается, приближаясь к метаконулю. Талон почти не выражен.
р4 простого строения, с острой главной вершиной; зуб слабо расширен в заднем отделе. Обычно нет дополнительных зубчиков, но иногда присутствует небольшой передний. По язычному краю коронки проходит цингулид.
Нижний хищнический зуб m1 длиннее, чем m2. Его протоконид очень высокий, массивный. Метаконид крупный, одиночный, сильно смещен назад от протоконида и по высоте равен паракониду. Выемка между тригонидом и талонидом на наружной стороне коронки лежит на уровне протоконида. Талонид почти вдвое короче тригонида, но значительно шире. Гипоконид массивный, относительно невысокий, без обособленной внутренней доли; его протокристид без дополнительного бугорка. Энтоконид небольшой, имеет вид одиночного невысокого бугорка, вплотную примыкающего к метакониду. Бассейн талонида широкий, открыт сзади.
m2 шире, чем m1, его коронка больше расширена в переднем отделе, чем в заднем. Протоконид значительно выше метаконида, перед последним расположен добавочный бугорок. Энтоконид одиночный, гипоконид массивнее и сильнее смещен назад. m3 маленький, при взгляде сверху округло-треугольный. Коронка окружена краевым валиком, на котором обособлен лишь протоконид.
Череп с нижней челюстью агриотерия (Agriotherium sp.) из позднего миоцена уезда Хэчжен в провинции Ганьсу, Китай. Общая длина черепа — 50 см, высота — 37,5 см.
Образ жизни и вымирание
Скорее всего, агриотерии предпочитали открытые ландшафты, держась поблизости от воды. Полагают, что их типичным местообитанием являлась степь с редкими деревьями и кустарниковыми зарослями, где также жили различные представители гиппарионовой фауны. Не исключено, что эти медведи также могли водиться в лесу или горах (или хотя бы в предгорьях). Учитывая палеоареал агриотериев, не охватывающий особо холодные районы, а также высокую степень плотоядности этих зверей, можно предположить, что, в отличие от подавляющего большинства современных медведей, они не впадали в спячку и не были склонны к интенсивному накоплению жира к зиме.
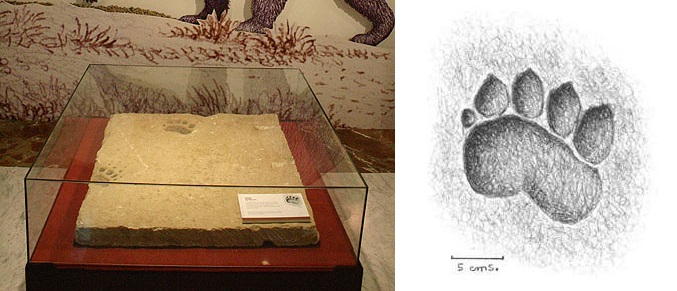
Следовые отпечатки агриотерия (Agriotherium sp.) из Испании и прорисовка отпечатка правой передней лапы.
Уже Кэтли и Фальконер (1868 г.) видели в агриотериях хищников или падальщиков, основываясь на их высококоронковых щечных зубах. Хищнические зубы Р4 и m1, служащие для разрезания плоти, у старых особей сильно изношены. Хендей (1980 г.) отмечал, что зубная система агриотериев отличается от таковой типичных падальщиков, например гиен, но не исключал возможности того, что звери охотились на таких больших копытных, как жирафы. Куртен (1968 г.), напротив, думал, что по образу жизни и питанию агриотерии не отличались от ныне живущих больших всеядных медведей.
По мнению Барышникова (2007 г.), крупные размеры и характерный для медведей экстерьер (несмотря на большую стройность телосложения относительно современных медведей) не позволяют считать агриотериев активными охотниками, поскольку им было трудно подкрадываться к жертве и совершать стремительные броски. Вряд ли они могли успешно конкурировать с такими быстрыми и проворными хищниками, как крупные кошачьи и псовые, многие из которых к тому же охотятся сообща. Вместе с тем значительные размеры и большая физическая сила позволяли агриотериям фуражировать обширные территории и отбирать добычу у менее крупных плотоядных или отгонять их от падали (клептопаразитизм).

Ранний плиоцен Южной Европы. Агриотерий (Agriotherium insigne) отбирает у молодого гомотерия (Homotherium crenatidens) добытого им митиланотерия (Mitilanotherium sp.). Автор — Р. Евсеев.
Сильные клыки, щечные зубы дробящего типа, высокий сагиттальный гребень, широкие скуловые дуги и нижняя челюсть с премассетерной ямкой указывают на сильное развитие челюстных мышц, позволявшие расчленять туши павших толстокожих, недоступные для других плотоядных. Хотя агриотерии скорее всего специализировались на поедании падали и отнятой добычи, они могли порой и сами охотиться, а в сезон поспевания плодов переходить на питание растительностью. В пользу всеядности свидетельствует крайне широкое распространение этих медведей. Неприхотливость в выборе пищи помогало агриотериям осваивать самые разнообразные местообитания. Однако, в любом случае, процент животной пищи в их рационе был более значителен, чем у любого из ныне живущих медведей, за исключением белого (Ursus maritimus), на что указывает все телосложение и зубная морфология. Изотопный анализ зубной эмали подтверждает, что агриотерии действительно потребляли значительное количество мясного корма, подобно некоторым популяциям бурого медведя (Ursus arctos).
Следует отметить, что даже бурый медведь, который имеет гораздо более тучное телосложение, время от времени может успешно охотиться на таких крупных и быстроногих животных, как олени и лоси. Несмотря на свою, казалось бы, неуклюжесть, бурый медведь способен на короткой дистанции развивать весьма внушительную для такого громоздкого животного скорость — более 50 км/ч. Помимо этого, бурый медведь очень вынослив и способен подолгу гнать преследуемое копытное. Агриотерии, как уже отмечалось выше, обладал более стройным телосложением и имел более длинные конечности. Логичным будет предположить, что эти звери мог развивать более высокую скорость, чем бурый медведь и обладали, скорее всего, еще большей выносливостью. Таким образом, получается, что независимо от того были ли агриотерии преимущественно плотоядными или наоборот растительноядными, в любом случае доля животного корма в их рационе была более значительной, чем у бурого медведя.

Ранний плиоцен Южной Африки. Агриотерий (Agriotherium africanum) рыщет в поисках пищи. На него опасливо поглядывает сиватерий (Sivatherium hendeyi). На заднем плане пасутся южноафриканские мамонты (Mammuthus subplanifrons). Автор — Р. Евсеев.
Вышеизложенное относится прежде всего к виду Agriotherium sivalensis. Несколько более мелкий и примитивный Agriotherium insigne потреблял, вероятно, больше растительной пищи, чем Agriotherium sivalensis, хотя сильные зубы дробящего облика предполагают присутствие кормов животного происхождения и в его в рационе. Предложенные палеоэкологические реконструкции хорошо согласуются с фактом быстрого формирования обширного ареала Agriotherium.
О окраске вымерших животных судить трудно. Однако можно предположить, что будучи обитателями степей и аналогичных биотопах, агриотерии имел достаточно светлую окраску, вероятно светло-коричневую, схожую с таковой у сирийского бурого медведя (Ursus arctos syriacus). Возможно, однако, что их окрас был более сероватым или же бледно-бежевым, похожим на окрас северных подвидов пумы (Puma concolor).
Вымирание столь успешного и широко распространенного рода скорее всего было обусловлено иссушением климата и сокращением кормовой базы, происходившей в конце плиоцена. Исчезли многие толстокожие и охотящиеся на них саблезубые кошки, соответственно уменьшился и объем доступной для агриотериев пищи. Вероятно, это в равной степени справедливо и для близкого рода Huracan, представители которого могли исчезнуть в Северной Америке после вымирания там тяжеловесных носорогов и добывающих их саблезубых барбурофелид.