Строение и функции
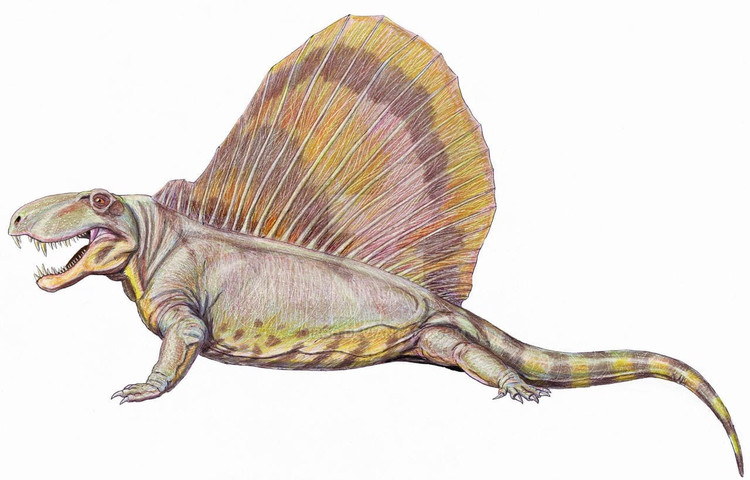
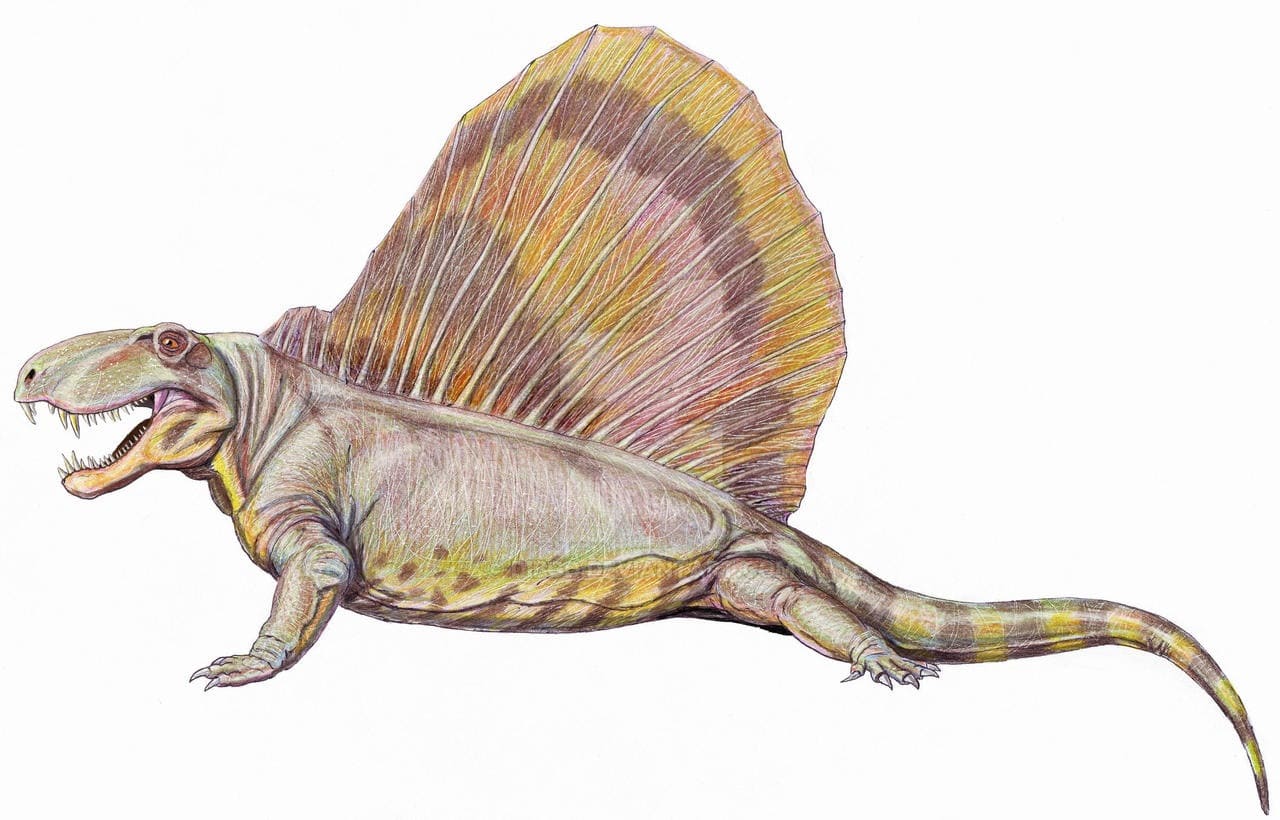
Реконструкция внешнего облика диметродона (Dimetrodon grandis, = Dimetrodon gigas), выполненная по существующей версии смонтированного скелета USNM 8635. Автор — Д. Богданов, 2009 г.
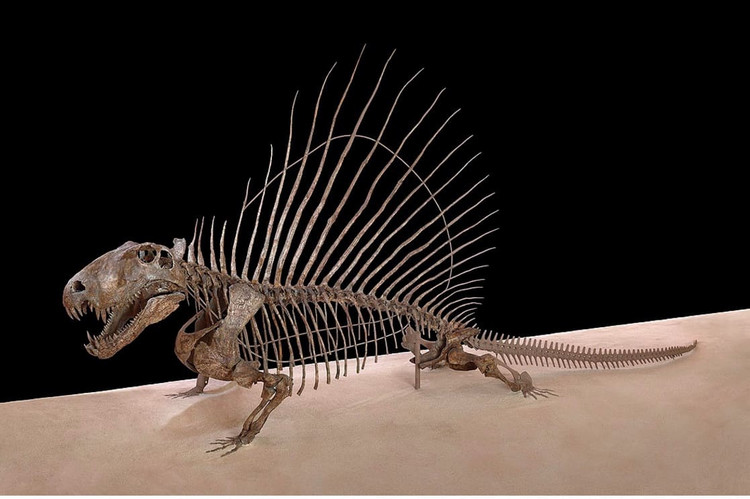
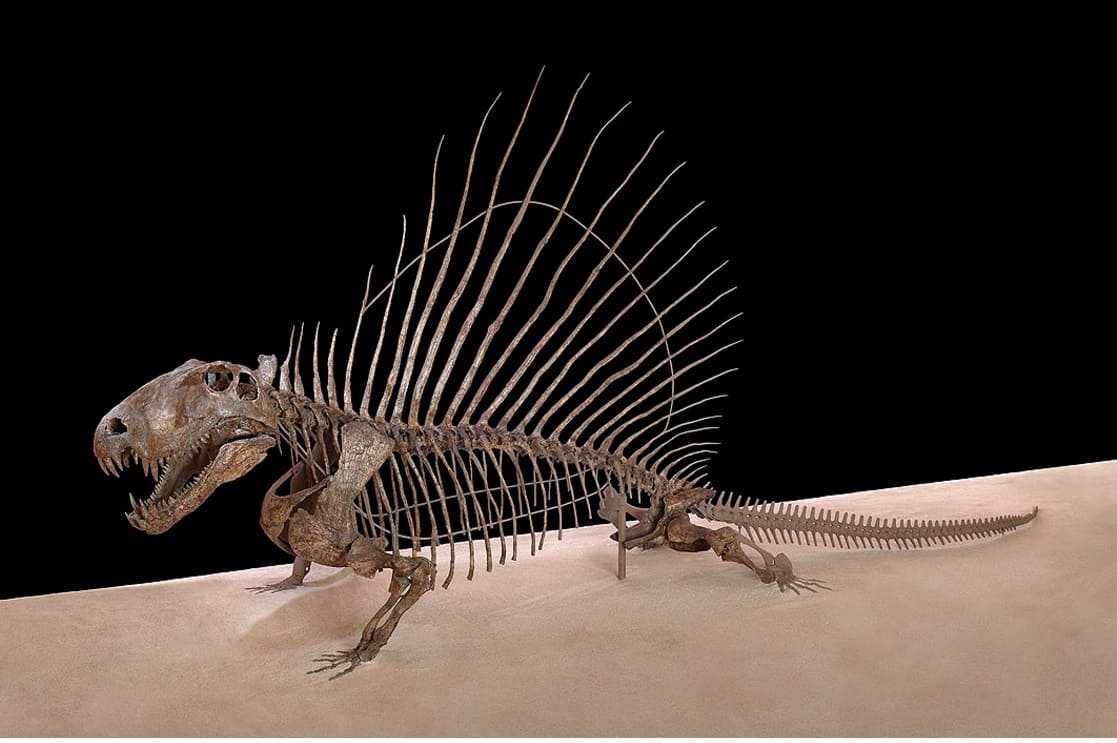
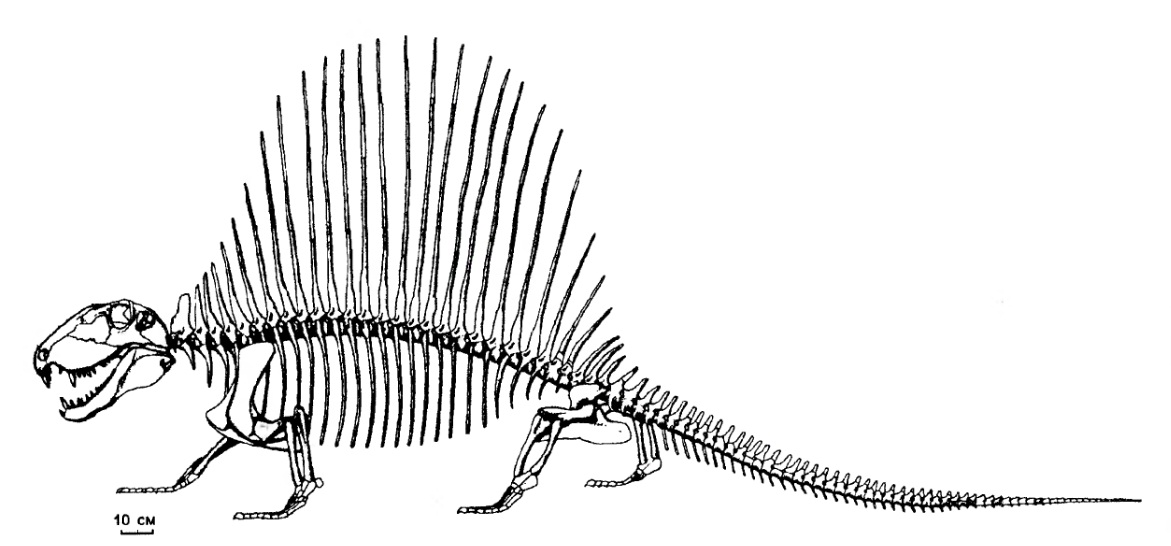
Современный вариант монтажа скелета диметродона (Dimetrodon grandis, = Dimetrodon gigas) USNM 8635 из ранней перми штата Техас, США.
Размеры Dimetrodon варьируют от мелких до очень крупных: общая длина составляет примерно от 0,8 до 4,5 м, а вес мог достигать около 250 кг.
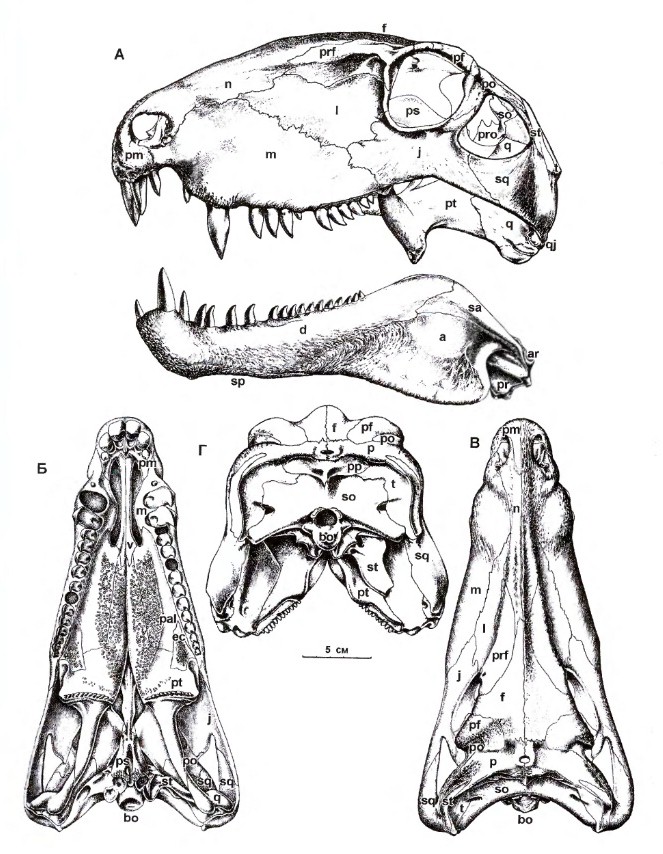
Прорисовки реконструированного черепа с нижней челюстью диметродона (Dimetrodon limbatus): А — сбоку; Б — снизу; В — сверху; Г — сзади.
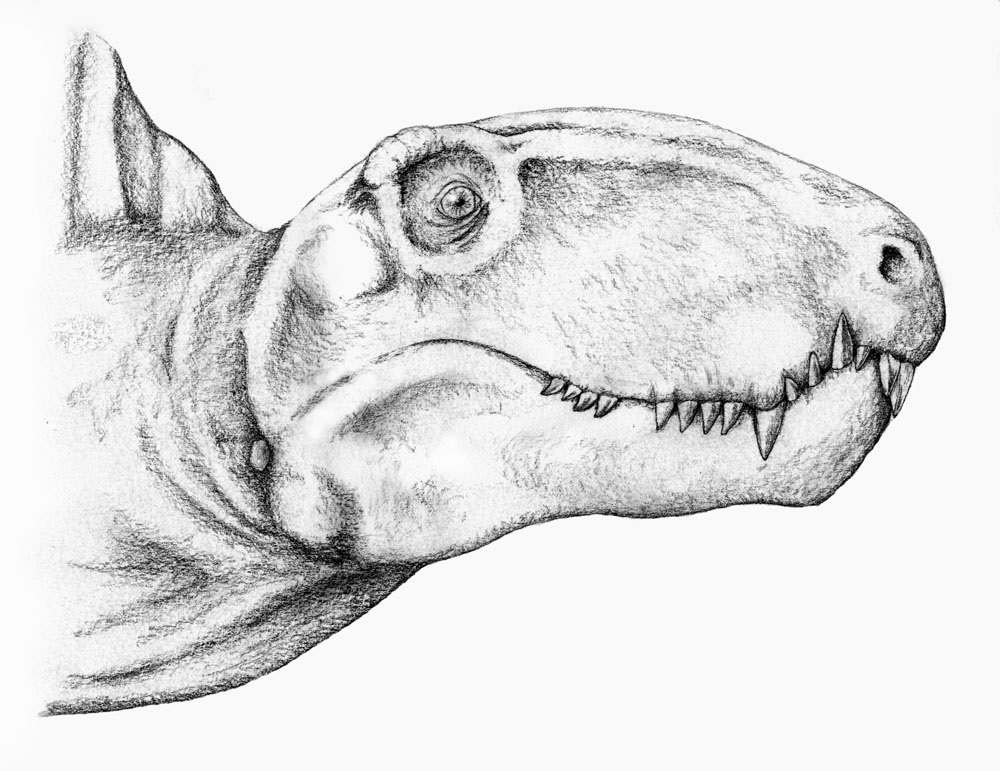
Череп высокий и несколько сжатый с боков. Предчелюстная кость отграничена от верхнечелюстной глубокой вырезкой. Передний край предчелюстной кости сильновыпуклый. Небольшие ноздри расположены в передней части морды. Слезная кость не достигает ноздри. Глазницы сдвинуты далеко назад. Несмотря на отсутствие барабанной перепонки, особенности строения головного мозга, а также костного лабиринта и полукружных канальцев внутреннего уха свидетельствуют, что слух мог быть развит примерно на уровне таких ныне живущих рептилий, как змеи.
Портретная реконструкция диметродона (Dimetrodon limbatus). Автор — Mojcaj.
Зубная система демонстрирует слабо выраженную гетеродонтию. Предчелюстная кость с 3 крупными коническими зубами. В передней части верхнечелюстной кости 2 или 3 зуба клыковидно увеличенные, загнутые назад, каплевидные в сечении, с острыми передним и задним кантами, которые у некоторых видов пильчатые. Клыковидный зуб нижней челюсти входит в вырезку между предчелюстной и верхнечелюстной костями. Верхне- и нижнечелюстные зубы перед клыковидными зубами редуцированы, а за ними постепенно уменьшаются в заднем направлении. Расположенные спереди челюстей клыковидные зубы позволяли быстрее умерщвлять жертву, а их канты эффективно разрезали плоть.
Прорисовка реконструированного скелета диметродона (Dimetrodon limbatus).
Остистые отростки шейных и туловищных позвонков резко удлинены, образуя в задней части шеи и на спине "парус" высотой до 1 м и более. Отростки Dimetrodon легко отличимы от таковых других пеликозавров, поскольку в дополнение к своей длине несут спереди исзади бороздки, делающие их гантелеобразными в сечении, а также у них отсутствуют поперечные выступы и бугорки, характерные для остистых отростков позвонков Edaphosaurus. Судя по всему, при жизни между отростками была натянута кожная перепонка. Верхушки отростков торчали из перепонки и были покрыты роговыми чехлами. "Парус" развивался постепенно в течение жизни, известны находки остатков молодых особей диметродонов с невысокими остистыми отростками. Во взрослом состоянии самки были мельче самцов и имели более низкий "парус".

Скелет молодой особи диметродона (
Dimetrodon sp.) в позе залегания из ранней перми юго-запада штата Техас, США.
Назначение спинного "паруса" диметродонов остается предметом дискуссий. Согласно различным догадкам, он использовался для теплообмена и повышения скорости метаболизма в светлое время суток, для демонстраций и визуального увеличения размеров тела, в качестве стабилизатора при плавании, или что вместо "паруса" на остистых отростках позвонков крепился жировой горб. На сегодняшний день наиболее вероятной представляется демонстративная функция "паруса".
Плечевая кость очень широкая в нижней половине, с энтэпикондилярным отверстием. Сочленовные поверхности костей хорошо развиты.

Слева — плиты раннепермской породы с отпечатками следов ихнотаксона
Dimetropus (по всей видимости, принадлежащие крупному
Dimetrodon) из Национального парка доисторических следовых дорожек на юге штата Нью-Мексико, США. Отдельные отпечатки достигают 15-20 см в длину. Справа — слепок плиты с приписываемыми
Dimetrodon правыми и левыми следовыми отпечатками, между которыми расположена вмятина от туловища, вероятно, образовавшаяся во время отдыха животного.
По современным представлениям, диметродоны могли выпрямлять конечности под туловищем и передвигались несколько по-другому, чем ящерицы и крокодилы, являясь весьма быстрыми охотниками своего времени. Что же касается скорости обмена веществ, то, согласно некоторым недавним исследованиям, у диметродонов она примерно соответствовала таковой современных варанов и крокодилов, что для пермского периода было весьма прогрессивным.
Старая реконструкция шагающего
Dimetrodon, изображающая его с широко расставленными в стороны конечностями.
Новая реконструкция идущего Dimetrodon с полувыпрямленными конечностями.