Особенности строения черепа и зубов
Особый интерес представляет череп эласмотериев. Самая броская и запоминающаяся деталь черепа — высокое куполообразное полое внутри вздутие лобных костей. Именно это образование побудило В.А. Теряева предложить вместо замысловатого латинского названия этого носорога простое русское — горболоб. Стенки купола сравнительно тонкие, но изнутри они укреплены многочисленными гребнями, пересекающимися в различных направлениях. Более крупные из них делят внутреннюю полость на несколько хорошо выраженных секций. Самая обширная из них — подкупольная. Она имеет очень тонкие стенки, и эта часть купола чаще всего (в находках) бывает разрушенной. В боковых частях выделяется по 3 секции с обеих сторон, их объем уменьшается спереди назад, в то время как толщина наружных стенок купола увеличивается. Растет и густота укрепляющих это образование гребней.
Относительно функционального назначения лобного вздутия существует несколько точек зрения. Большинство исследователей считают, что это было основание для рога. Дело в том, что наружная поверхность купола шероховатая. Почти все черепа несут следы крупных кровеносных сосудов, основная масса которых сосредоточена у основания купола. Однако у некоторых черепов эта поверхность слабошероховатая. У сибирского эласмотерия (Elasmotherium sibiricum) из г. Зеленокумска и на черепе эласмотерия Пэйя (Elasmotherium peii) из г. Токмака с территории Украины отпечатки сосудов на вершине купола отсутствуют. Степень развития сети кровеносных сосудов, вероятно, можно связать с половым диморфизмом эласмотериев. Известен череп, предположительно принадлежащий самке. Он меньшего размера, его лобный купол выражен слабее, причем во фронтальной части он плавно переходит в носовые кости. Отпечатки крупных кровеносных сосудов на вершине купола отсутствуют.
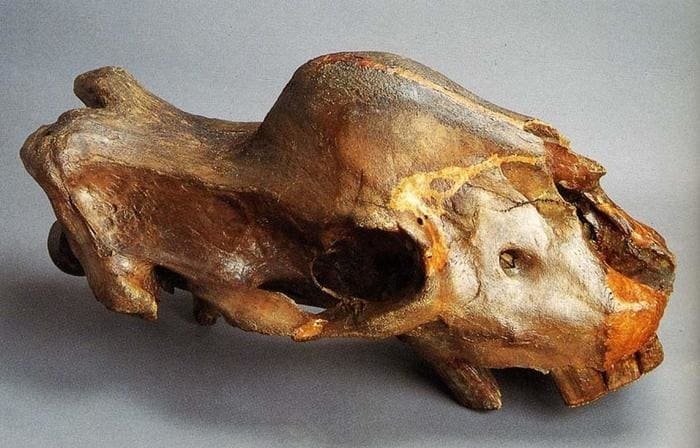
Череп молодой особи сибирского эласмотерия (Elasmotherium sibiricum), предположительно самки. Ростральная часть черепа разрушена. Зоологический музей Зоологического института РАН, Санкт-Петербург.
Обильное кровоснабжение и дополнительное укрепление лобного вздутия изнутри костными перемычками может служить косвенным доказательством наличия здесь какого-то образования. Однако у исследователей нет единства относительно его степени развития и формы, а также не ясно его функциональное значение. Современные носороги используют свой находящийся на конце морды рог для раздвигания густых зарослей и как турнирное оружие, однако остается неизвестным, насколько это было оправданно для гипотетического лобного рога эласмотериев.
С точки зрения А.К. Швыревой (2016 г.), наиболее удачной являются реконструкциия внешнего вида сибирского эласмотерия, выполненные профессором К.К. Флеровым (1953, 1959 гг.). Она представляет рог как колпачок мягких очертаний, плотно охватывающий купол. Этот ученый полагает, что данное образование было чрезвычайно прочным, рассматривая разделяющие его внутреннюю полость костные гребни в качестве контрфорсов, поддерживающих свод купола.
По мнению К.К. Флерова, эласмотерий представлял собой степное животное, питающееся, по меньшей мере отчасти, подземными частями растений, которые оно разыскивало, вспахивая почву мордой. Этим объясняются характерные особенности строения черепа этого зверя. Прежде всего, обращают на себя внимание его чрезвычайные прочностные характеристики. Узкая клинообразная форма лицевой части черепа образована крепкими носовыми, верхнечелюстными и предчелюстными костями. Они прочно сращены друг с другом и со сплошной костной носовой перегородкой. Следы обильного снабжения кровью указывают на возможность ороговения верхней стороны носовых костей (там, где, по мнению Брандта, находился малый рог). Очень развито костное небо с вырезкой, открывающейся на уровне III-го коренного. Большие выросты на переднем и верхнем крае глазниц защищали глаза при рытье.

Сибирские эласмотерии (Elasmotherium sibiricum) добывают подзменые части растений. Реконструкция выполнена К. Флеровым, 1953 г.
Зубы эласмотерия обладают очень высокими коронками с почти постоянным ростом. Их эмалевые петли, кроме основных, имеют еще и целый ряд мелких дополнительных извивов. От зубов всех других носорогов они отличаются своей крайней специализацией к интенсивному стиранию при питании очень плотным кормом, перемешанным с большим количеством попадающей вместе с ним почвы. Дополнительно длительный рост и почти абсолютная гипсодонтия зубов предохраняли животное от слишком скорой их потери.
Все шейные позвонки очень мощные; первый из них (атлант) имеет необычайно развитые поперечные отростки размахом до 70 см. Высота остистых отростков грудных позвонков может превышать 50 см. Это заставляет предполагать, что мускулатура, поддерживающая тяжелую голову, была развита крайне сильно. Толщина слоя мьппц шеи (трапециевидный мускул, широкий и другие) достигала, видимо, около полуметра. Лишь немного меньшей была толщина боковых шейных мышц (грудинно-головной, плечеголовной, плече-подъязычной и др.), о чем можно судить по строению атланта и сильному развитию затылочных гребней. К.К. Флеров делает вывод, что череп и шея приспособлены к сильному нажиму при какой-то огромной работе головой. Это дает ученому основание допустить возможность питания эласмотерия подземными частями растений.

Продольный распил задней части черепа сибирского эласмотерия (Elasmotherium sibiricum), демонстрирующий ячеистое строение подкупольной части и мозговую полость.
С подобным образом жизни связывает К.К. Флеров и назначение куполообразного возвышения на лобных костях черепа и ограничиваемых им полостей. Он считает, что эти образования являются не столько опорной структурой рога, сколько органом усиления обоняния. Внутренние полости купола — сообщающиеся с полостью носа гипертрофированные лобные пазухи — являются, по Флерову, дополнительным ольфакторным пространством. Именно в области задних концов носовых и передних концов лобных костей у многих других млекопитающих помещаются ольфакторные, т. е. обонятельные полости, связанные с усилением обоняния. Для животного же, питающегося подземными частями растений, такие полости становятся необходимыми, а их размеры — существенными. Колпаковидный рог же, скорее всего, только защищает лобный купол, нежели выполняет иную роль.
По всей видимости, обоняние эласмотерия действительно было обостренным, потому что обонятельные доли его головного мозга, по данным Дешазо (1958 г.), необыкновенно крупные и примыкают непосредственно к задней части подкупольной полости черепа. По мнению А.К. Швыревой (2016 г.), немаловажную роль в усилении обоняния эласмотерия играли и 2 пары небных щелей, расположенные в передней части костного неба. Надо полагать, что обоняние в жизни этого носорога играло ведущую роль по сравнению со слухом, а тем более — со зрением.
Использование клинообразной морды для выкапывания подземных частей растений представляло собой для эласмотериев ту самую "адскую" механическую работу, которая могла являться одной из причин, вызывающих перегрев организма и его гибельные последствия. Купол, по мнению А.К. Швыревой (2016 г.), являлся и органом терморегуляции, поскольку носовая и подкупольная полости связаны, а наиболее крупные следы кровеносных сосудов отпечатаны на самых тонких стенках наружной части купола. Роль градирни играл здесь сам купол, внутрь которого через нос поступал холодный воздух. Функцию охлаждающей жидкости выполняла кровь. Ток горячей крови, соприкасаясь со стенками купола, охлаждался, предотвращая общий перегрев организма.

Вид спереди на заднюю часть черепа подвзрослой особи сибирского эласмотерия (Elasmotherium sibiricum) из Саратовской области, Россия.
Сравнивая строение черепа и шейных позвонков ныне живущих носорогов и сибирского эласмотерия, В.В. Титов и В.С. Байгушева (2021 г.) подвергают сомнению предположение о наличии на его лобном возвышении массивного рога. Авторы указывают, что поверхность купола заметно отличается по своей структуре от шероховатых оснований для рогов на черепах других носорогов. Лобное возвышение эласмотерия больше напоминает пронизанное кровеносными сосудами костное основание для кератинового рога полорогих (Bovidae). Поэтому кажется более вероятным, что лобное возвышение скорее было покрыто слоем кератина, который в первую очередь служил защитой относительно тонкой костной стенки. Роговой слой нарастал от основания, хорошо заметного в нижней части костного купола, а его верхний конец был направлен верхне-задне .Поскольку возвышение на черепах самцов развито сильнее и имеет более рельефную поверхность, покрывающий его кератиновый "шлем" мог быть более массивным, чем у самок.
Исследователи также полагают, что купол служил для обострения обоняния и, вероятно, мог функционировать как резонирующая камера для усиления издаваемых звуков. Эта гипотеза основана на структурном подобии лобного возвышения эласмотерия с таковым ископаемой антилопы рузингорикса (Rusingoryx atopocranion), а также гребнями утконосых динозавров (Hadrosauridae). Предполагается, что все они использовали эти образования в качестве своеобразных рупоров. Если это действительно так, то любопытными выглядят данные о наличии у эласмотериев полового диморфизма и меньшем куполе на черепах самок. Представляется возможным допустить, что самцы были "горластее" последних и во время гона старались произвести впечатление на соперника своим громким ревом.
Положение кератинового нароста на лбу предполагает, что он не был задействован или использовался лишь ограниченно в битвах за самок и территориальных сражениях, в отличие от рогов сегодняшних носорогов. Поскольку единственный или более крупный из рогов у сегодняшних носорогов расположен на носу, его можно использовать гораздо эффективнее, поскольку при движениях головы мышцы шеи передают на рог больше силы или импульса. Из этого следует вывод, что схватки у эласмотериев носили высокоритуальный характер. Также, по мнению ученых, шероховатость на расширенном переднем крае сращения носовых и предчелюстных костей предполагает наличие на конце морды узкой ороговевшей подушки, служившей для раскапывания почвы в поисках подземных частей растений. На шее особо выделялись мощные мышцы, осуществляющие боковые и верхне-боковые движения головы.

Портретные реконструкции сибирского эласмотерия (Elasmotherium sibiricum) с кератиновым наростом на лобном куполе, выполненная по черепу самца из г. Атырау (Гурьева), Атырауская область, Казахстан. Выполнены Р. Учителем по представлениям В. Титова и В. Байгушевой, 2021 г.
Очевидно ландшафт, в котором жили эласмотерии, характеризовался сменой времен года. В засушливый и холодный сезон, когда надземная травянистая растительность исчезала, пространство ресурсов корректировалось за счет добычи подземных частей растений. Причем питание таким кормом, по мнению В.В. Титова (2008 г.), было для эласмотериев облигатным, а не факультативным, как считалось ранее. Вспахивая землю, словно плугом, мощным клинообразным рылом с ороговевшим концом и используя экстраординарное обоняние, эти носороги безошибочно находили сочные корневища, луковицы, клубни. Устойчивые к стиранию зубы успешно противостояли абразивным частицам, поступающим вместе с пищей.
У носорогов плоскость затылка образует с линией основания черепа угол, величина которого у разных их видов не одинакова. Ф. Цойнер (1934, 1936 гг.) произвел измерение этого угла и отметил, что его величина является показателем положения головы по отношению к шее и туловищу, а следовательно, говорит о том, какой ярус растительности мог служить источником пищи для ископаемых форм. Так, у ныне живущих индийского (Rhinoceros unicornis) и черного (Diceros bicornis) носорогов, питающихся преимущественно побегами и листьями кустарников на уровне более 1 м над поверхностью земли, затылочный угол менее прямого. В то же время у белого (Ceratotherium simum) и шерстистого (Coelodonta antiquitatis) носорогов, питающихся травянистой растительностью на высоте менее 0,5 м — угол близок к прямому или тупой, а голова, в отличие от индийского и черного, по отношению к туловищу наклонена вниз. Затылочный угол эласмотерия оказался наиболее тупым из всех измеренных. Это указывает на то, что его голова была сильнее, чем у других носорогов, наклонена по отношению к туловищу.
У черепа эласмотерия Пэйя из г. Токмака плоскость затылочной кости образует с линией основания черепа угол в 105 °, а у сибирского эласмотерия из г. Зеленокумска — в 106 °. У черепа этого животного из собрания Саратовского краеведческого музея данный угол достигает 110-115 °, а его череп, хранящийся в ГГМ РАН, имеет соответствующий угол близкий к прямому. Очевидно, в разных популяциях в пределах вида этот признак значительно варьировал, но, тем не менее, угол отклонения затылочной кости относительно линии основания черепа неизменно оставался больше прямого. Следовательно, эласмотериев следует отнести к носорогам с низкой посадкой головы. Это позволяет предполагать, что эласмотерий, подобно белому и шерстистым носорогам, являлся обитателем открытых ландшафтов и питался низкорасположенной травянистой растительностью.

Угол наклона плоскости затылочной кости к линии основания черепа носорогов.
На питание "подножным кормом" также указывают крупные глоточные бугры в основании черепа эласмотериев. Их величина и шероховатость обусловлены сильным развитием длинных мускулов шеи в связи с постоянным наклоном головы во время пастьбы. Чрезвычайно слабое развитие такого экологического признака как предглазничная ямка стало, по-видимому, следствием перехода к питанию низко расположенной растительностью.
Достигнув крайней степени стенофагии, эласмотерии приобретают специфические структурные изменения зубной системы. Мощная эмалеукрепленность коренных зубов и моляризация предкоренных явились результатом высокой степени абразивности потребляемой пищи. Почти полная гипсодонтия зубов эласмотериев — это также одно из приспособлений к питанию жесткими кормами. Таким образом, специфика строения зубной системы этих носорогов приобретает экологический смысл.
Вместе с тем, В.И. Жегалло и Н.Г. Носкова (2001 г.) полагают, что кормовая база таких крупных животных как эласмотерий не может быть обеспечена биотопами с низкой продуктивностью ксерофильных растений и должна быть приурочена либо к высокотравным степям, либо к околоводным пространствам. По мнению этих исследователей, абсолютно гипсодонтные щечные зубы эласмотерия обусловлены не столько жесткостью поедаемой растительности, сколько наличием в корме абразивных минеральных частиц, число которых возрастает при питании корнями или подземными частями побегов растений. Их доля значительно увеличивается при питании растениями околоводных стаций, произрастающими на влажном субстрате.
В связи с тем, что у эласмотериев отсутствовали резцы, очевидно, захват пищи происходил с помощью подвижной верхней губы и языка, опирающихся на кости ростральной части черепа и ложковидный симфиз нижней челюсти. О степени развития верхней губы можно судить по сильным шероховатостям, расположенным по резцовому краю и частично по диастеме. Заложенная в толщу губы мощная мускулатура способствовала не только раскрытию или смыканию ротовой щели, но позволяла производить разнообразные движения. Ороговевшее клиновидное рыло, исключающее роль губ и круговой мыщцы рта в захвате корма, отсутствие резцов, исключающее срезание кормового пучка, в сочетании с длинной диастемой, имеющей ороговевший край, и гипертрофией поперечных отростков I-го шейного позвонка, — все это позволяет утверждать, что у эласмотерия доминировал боковой способ захвата пучка и боковой отрыв его за счет резкого движения головы. Откус кормового пучка мог осуществляться, очевидно, с помощью скрытых в десне бугровидных костных образований, лежащих по зубному краю впереди диастемы. Фронтальный захват был возможен при взаимодействии мощного языка с ложковидным ороговевшим симфизом нижней челюсти.
В.И. Жегалло и Н.Г. Носкова отмечают, что конструкция самого аппарата захвата, где сочетается ложковидный симфиз и клиновидное рыло верхней челюсти, может быть объяснена и с позиции питания подземными частями водньх и околоводных растений. Ложковидная нижняя челюсть легко проникает в полужидкий грунт, зачерпывая вместе с ним подземные части растений. Клиновидная верхняя челюсть при смыкании прижимает растительный пучок и сохраняет его фиксированным до тех пор, пока боковым движением головы он не будет вырван из субстрата. Подобный механизм рассчитан, в первую очередь, на добычу растений, размножающихся корневищами (из однодольных — многие осоковые, рогозовые, наядовые, из двудольных — кувшинковые), прочно закрепленными в субстрате при помощи многочисленных придаточных корней. Принимая во внимание то, что водные и полуводные цветковые, как правило, достаточно неприхотливы к типу субстрата, и многие из них с легкостью растут на высокоабразивном песчаном грунте, можно выдвинуть дополнительный аргумент и в обоснование "абсолютной" гипсодонтии щечных зубов эласмотерия.
Тем не менее, В.И. Жегалло и Н.Г. Носкова не определяют эласмотериев как узкоспециализированных полуводных животных, как это предлагал В.А. Теряев. Если строение аппарата захвата свидетельствует в пользу его гипотезы, то пропорции скелета и особенности строения локомоторного аппарата эласмотериев демонстрируют адаптации к освоению открытых пространств. Это позволяет добавить к областям обитания этих носорогов луговые и степные биотопы, с которыми могли быть связаны пути миграций между предпочтительными кормовыми угодьями. Здесь имеет смысл возвратиться к представлениям К.К. Флерова об эласмотерии как о звере-пахаре, добывающем подземные части растений, взрыхляя почву своей заостренной мордой как плугом.
Сочетая элементы реконструкций, предложенных В.А. Теряевым и К.К. Флеровым, а также учитывая состав сопутствующей фауны, можно охарактеризовать палеогеографические условия обитания эласмотериев. По всей видимости, они населяли степную ландшафтную зону с входящими в нее интразональными околоводными сообществами растений, служившими этим носорогам предпочтительными кормовыми угодьями. В северной части ареала, находящегося в лесной зоне, эласмотерий был долинным интразоналом. К интразональным биотопам, помимо пойменных участков рек, могли относиться и зарастающие пруды, в зоне осушения которых развитие корневищных растений идет особенно интенсивно. Залесенных территорий, по-видимому, эласмотерий избегал, и одной из причин его вымирания можно считать увеличение влажности климата и наступление леса.
Особенности строения конечностей
Об образе жизни эласмотериев и условиях их существования не в меньшей мере, чем зубная система и особенности строения черепа, достоверно свидетельствуют мышечно-связочная скульптура и суставной рельеф костей конечностей.
Отсутствие в коллекциях V-й пястной кости и значительные размеры фасеток для этой кости на крючковидной и IV-й пястной костях послужили основанием для возникновения ошибочного представления о четырехпалости кисти эласмотериев, а также об его экологии как обитателя заболоченных, топких берегов древних водоемов (Теряев, 1930 г.). Находки на реке Большой Караман, являющейся притоком Волги (Беляева, 1939 г.), а также материалы с территории Ставропольского края показывают, что кисть и стопа эласмотериев имели по 3 функционирующих пальца. IV-й палец кисти представлен всего 1 маленькой рудиментарной костью, не имеющей фасеток для фаланг. Ставропольские материалы дали новые сведения о головчатой, трапецийной, гороховидной, сезамовидной костях кисти. Впервые предоставился полный материал для изучения строения кисти и стопы эласмотериев.
Средний палец кисти и стопы эласмотериев наиболее развит, имеет длинные и мощные III-и пястные и плюсневые кости, хорошо развитое копыто. Боковые пальцы намного меньше и сдвинуты относительно среднего назад. Основная тяжесть в момент опоры ложится на центральный луч автоподия. Об этом можно судить по расширению фаланг среднего пальца и уплощению их суставных поверхностей, а также по прочности связей костей запястья и заплюсны в центральном отделе. Очевидно, конечности эласмотериев демонстрируют тенденцию к редукции боковых пальцев и выработке приспособлений в направлении однопалости, что характерно для животных, передвигающихся по твердым субстратам (Флеров, 1953 г.).
Наоборот, конечности эласмотериев явно не были приспособлены для поддержания их громадной массы на мягких илах и вообще на вязких грунтах. Водные и болотные непарнопалые, как, например, тапиры и вымершие аминодонты, обладают четырехпалыми передними конечностями. Адаптированная к передвижению по топким болотам нога гиппопотама имеет боковые пальцы лишь немного меньше средних. Общая нагрузка тела эласмотериев на площадь опоры конечностей при его огромных габаритах была много больше, чем у гиппопотама или даже черного носорога (ширина стопы сибирского эласмотерия около 30 см, т. е. приблизительно такая же, как у гиппопотама и двурогого носорога), а вес эласмотерия был не менее 4 т, гиппопотама же — 3 т.
На передвижение эласмотериев по сухим и твердым грунтам указывает положение и степень уплощения передних стенок и суставных поверхностей костей запястья и заплюсны. Кости названных отделов кисти и стопы имеют асериальное расположение, что предотвращает боковое раздвижение костей в момент опоры. Этому препятствует и устройство дистальных суставных поверхностей костей предплечья и голени. Судя по расположению, форме и рельефу суставных фасеток, наибольшую амплитуду сгибательно-разгибательные движения имели место в момент переноса конечностей. В момент же опоры и максимального разгибания дистальные отделы образовывали тугой замок.
Для выяснения ландшафтной привязанности сибирского эласмотерия было произведено сравнение устройства дистальных отделов его конечностей с таковыми у ныне живущих носорогов, черного и белого. Оно показало, что сибирский эласмотерий по взаиморасположению костей запястья и заплюсны ближе стоит к белому носорогу. Не противоречат этому и угловые показатели черепов. Это дает основание сделать вывод, что сибирский эласмотерий был обитателем ландшафта, близкого по характеристикам к саваннам, но еще более остепненного.
Интересным представляется анализ соотношения длины сегментов конечностей сибирского эласмотерия и других носорогов. За 100 % была принята длина сегментов передних и задних конечностей ныне живущего белого носорога. Оказалось, что сибирский эласмотерий имел самые длинные и стройные конечности среди ископаемых и ныне живущих носорогов. Это дает основание предположить, что передвигался он быстрее и легче своих сородичей. Ускорение бега достигалось за счет значительного удлинения метаподиев, усилением в них срединного луча и частичной редукцией боковых, что характеризуется как приспособление к ускоренному бегу. А скоростное передвижение требует больших затрат кислорода, что влечет за собой увеличение носовой полости у животных.
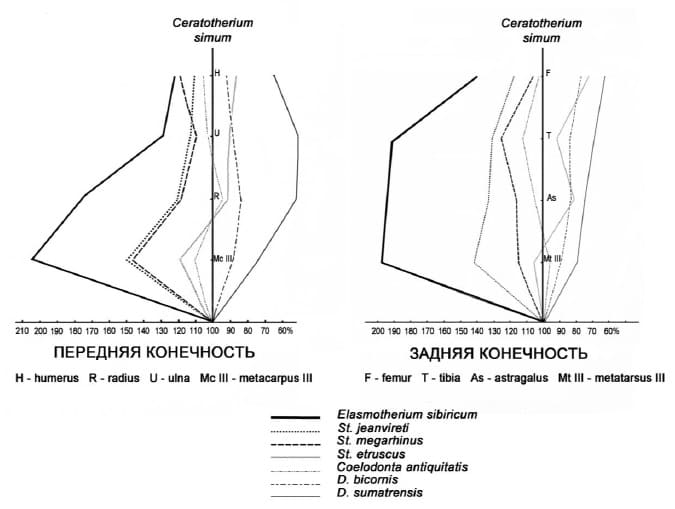
Соотношение длины сегментов конечностей сибирского эласмотерия (Elasmotherium sibiricum) и других носорогов. Размеры костей носорогов, кроме сибирского эласмотерия, взяты из работы К. Гирена (1980 г.). Длина костей конечностей белого носорога принята за 100 %.
Судя по большой глубине носовой вырезки, задний край которой располагается на уровне заднего края альвеолы М1, у эласмотериев носовая полость была развита хорошо. Глубина носовой вырезки интерпретируется как экологический показатель. Среди копытных животных обладатели глубокой вырезки, как правило, являются обитателями открытых ландшафтов (лошади, сайгак и др.). Судя по развитию носовой вырезки, эласмотерии относятся к их числу.
Локомоторные способности животных определяются совокупностью признаков строения конечностей, позвоночника и отчасти черепа. Для определения способа и формы бега сибирского эласмотерия А.К. Швыревой (2016 г.) применена методика, разработанная П.П. Гамбаряном (1972 г.). Этому носорогу, подобно слонам, был присущ дилокомоторный дорзостабильный способ бега, когда животное добивается ускорения тела толчком о землю и передних, и задних конечностей, причем спина на всем протяжении цикла движения остается жесткой.
Передние конечности являются активно опорными. Они первыми принимают на себя нагрузку при опоре о землю и тянут вперед тело животного. После опоры наступает фаза подготовки нового толчка. Сила толчка передних ног частично достигалась за счет работы кистей эласмотерия по принципу заведенной спирали. В статичном состоянии кисть слегка развернута наружу (супинация), при движении в фазе подготовки толчка происходит поворот ее вовнутрь (пронация), конечность как бы ввинчивается в субстрат и, испытывая его противодействие, согласно третьему закону динамики, обеспечивает мощь толчка. Далее вынесенная вперед конечность за счет работы мускулов-сгибателей подтягивает вперед тело животного. О значительном сгибании в запястно-предплечевом суставе в фазе переноса конечности свидетельствует сильное развитие гороховидной кости. Она обладает мощным бугром (индекс гороховидной кости 128,6), сплошь покрытым шероховатостями: местами крепления локтевых сгибателей и разгибателей запястья. Бугор гороховидной кости исполняет роль рычага силы.
В процессе бега локомоторную роль активно играют задние конечности. Они толкают тело животного вперед, обеспечивая ускорение. Задние конечности, судя по величине углов, под которыми сочленяются их сегменты, действовали как сжатые пружины, обладающие запасом потенциальной энергии, которая в момент толчка преобразуется в кинетическую, при этом стопа животного обеспечивает размах движений конечности и перемещение тела. О силе сгибательно-разгибательных движений в голеностопном суставе эласмотерия свидетельствует строение пяточной кости. Ее пяточный бугор мощный, но короткий. Вершина его шишковидно вздута и очень шероховата. Укороченность пяточного бугра, который является рычагом приложения силы разгибателей голеностопного сустава, свидетельствует о большой мощи этой мускулатуры. Это подтверждается не только сильным развитием шероховатостей пяточного бугра, но и интенсивными шероховатостями наружной и внутренней стенок тела пяточной кости. В связи с мощным развитием мускулатуры, разгибания в голеностопном суставе происходят резко, что обеспечивает скорость передвижения животного.
В процессе эволюции конечности эласмотериев приобретают ряд приспособлений амортизационного характера. Проксимальным амортизатором является эластичный пояс мышц и фасций этих мышц переднего и заднего поясов конечностей. Амортизационное действие передается через плечевой и коленный суставы. При этом в плечевом суставе, на что впервые обратил внимание В.К. Грегори (1912 г.), достигается экономия мышечной энергии за счет явно спиральной формы плечевой кости.
Следующий амортизационный узел располагается в запястье и заплюсне с их внутрисвязочным и синовиальным аппаратами. К пружинным приспособлениям принадлежат многочисленные мелкие внутрисуставные поверхности между костями запястья и заплюсны. Поскольку эти фасетки поставлены под различными углами, они способствуют разложению на множество составляющих сил тяжести и противоудара, возникающих в момент соприкосновения конечности с землей в фазе опоры.

Кисть (слева) и стопа (справа) сибирского эласмотерия (Elasmotherium sibiricum), а также схемы их строения.
Проксимальные концы боковых плюсневых костей возвышаются над средней плюсневой костью. Такое расположение, видимо, тоже выполняло функцию амортизатора, когда при толчке III-го пальца о субстрат они могли смещаться дистально, смягчая удар при передаче его силы на вышележащие отделы конечностей. Роль боковых лучей как пясти, так и плюсны, смещенных относительно среднего назад, очевидно, сводилась к ограничению погружения дистальных отделов конечностей в грунт и для принятия на себя части давления тела при экстремальных нагрузках обременения среднего луча. Они как бы подключались к опоре, амортизируя удар о субстрат. Эласмотерии — копытоходящие животные. Поэтому универсальным рессорным механизмом в обеих конечностях является копыто.
К амортизационным устройствам конечностей эласмотериев следует отнести и сезамовидные кости. Функционально они являются флексорными тормозами и блоками переброски сухожилий сгибателей. В момент наибольшего сгибания пальцев в фазе переноса конечностей они сдвигались проксимально, выходя верхними концами за пределы своих суставных поверхностей на пястях и плюснах. При этом их верхние концы сближались, а нижние расходились и упирались в первые фаланги. В момент опоры они обеспечивали более жесткую фиксацию пястно- и плюсно-фаланговых суставов. Находясь в крайнем нижнем положении, когда их проксимальные концы не выходили за уровень суставных поверхностей на прилежащих костях, сезамовидные кости исполняли роль амортизаторов удара о землю.
Учитывая длинные конечности эласмотериев, можно предположить, что им, несмотря на значительный вес, была свойственна полутяжелая форма бега типа галопа (ныне живущие носороги переходят на галоп лишь в исключительных случаях). Современный белый носорог бегает со средней скоростью 30 км/ч, развивая максимально 40-45 км/ч. Поскольку эласмотерии достигали гораздо более крупных размеров, скорость их передвижения явно была существенно меньше.
Обобщая вышесказанное, подчеркнем, что особенности строения кисти и стопы эласмотериев указывают, что их развитие шло в сторону выработки системы приспособлений к передвижению по жестким субстратам. Перечисленные выше приспособления в фазе опоры (момент удара о субстрат) препятствуют раздвижению костей и обеспечивают тугой замок в суставе. Это было залогом успешного освоения ландшафтов с твердыми грунтами.