Особенности строения позвоночного столба
Синхронно с выработкой специфических приспособлений органов локомоции эласмотерии приобрели черты строения позвоночника, постоянно обеспечивающие жесткость спины. В целом позвоночный столб этих носорогов имеет арочную конструкцию. При взгляде на их скелет в профиль обращает на себя внимание высокая холка в области первых грудных позвонков. В.Я. Бровар (1935, 1940 гг.) рассматривает холку как особый орган жесткости позвоночника, работающий против его деформации посредством взаимодействия остистых отростков и связочного аппарата. При идеальной холке (термин Бровара) остистые отростки, образующие ее, должны быть максимально длинными, чтобы обеспечить экономичность работы всего аппарата жесткости.
При раскопках скелета сибирского эласмотерия в г. Зеленокумске производился замер названных отростков. Наибольшая длина их составила 51-53 см. Холку у эласмотериев образуют остистые отростки I-VI-го грудных позвонков. Шероховатость и грибообразное расширение спереди назад их верхушек указывают на значительное развитие надостистой связки. Начинается она от III-IV-го грудных позвонков и тянется по вершинам всех грудных и поясничных позвонков. Она является как бы продолжением парной выйной связки. Глубокая выйная впадина на затылочной кости черепа говорит о наличии сильной парной выйной связки, которая по существу является антагонистом надостистой связки.
Столь сильный аппарат жесткости грудного отдела позвоночного столба выработался у эласмотериев для противодействия сгибанию позвоночника (флексии) в результате воздействия на него реактивной силы передних конечностей в момент опоры на субстрат в процессе движения животного. Развитию холки способствовало также наличие тяжелой головы и выполнение зверем трудоемкой работы по добыче корма из-под земли.
К особенностям строения позвоночного столба эласмотериев следует отнести необычайно крупный атлант. Экстраординарные размеры атланта, совершенно фантастическое развитие его крыльев, сильное развитие мускульных шероховатостей и бугров, а также могучие, отклоненные назад затылочные гребни черепа дают основание предполагать наличие мощной шейной мускулатуры, выполняющей изгибающее действие. В шейном отделе позвоночника жесткость обеспечивается сильным развитием парных суставных и поперечных отростков с их связочным аппаратом. Экстензивным тормозом, помимо отростков позвонков, очевидно, выступают и затылочные гребни черепа.
Арочная конструкция грудного и поясничного отделов позвоночника обусловлена особым строением тел позвонков. Они характеризуются очень выпуклой головкой впереди и столь же глубокой ямкой сзади. Тело имеет скос спереди назад, в результате чего суставные поверхности оказываются на разновысоких уровнях и при сочленении тел позвонков обеспечивают форму широкой пологой арки. Вместе со связочным аппаратом она имеет жесткую конструкцию, противостоящую разгибающему действию позвоночника (экстензия).
Об объеме грудной и брюшной полостей эласмотериев мы можем судить по остаткам грудной клетки сибирского эласмотерия, полученным в результате раскопок в ст. Гаевской. По форме она была бочковидной, а по размерам — внушительной, что характерно для крупных травоядных животных.
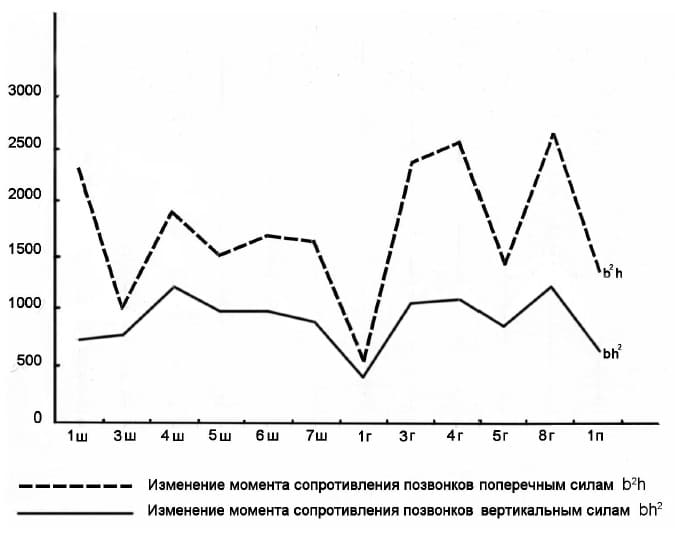
Для вычисления моментов сопротивления сил, действующих на сгибание позвоночника в вертикальной и поперечной плоскостях, А.К. Швыревой (2016 г.) использована методика Е. Слайпера (1946 г.). Материалом для этого послужили позвонки, в основном полученные во время раскопок скелета сибирского эласмотерия в г. Зеленокумске.
Расчет был произведен для каждого отдельно взятого позвонка: момент вертикальных сил по формуле W = bh2/1000 (куб. мм), где b — ширина задней суставной поверхности тела позвонка, h — ее высота; момент поперечных сил по формуле W = b2h/1000 (куб. мм). Рассчитанные моменты сопротивления сведены в график. Анализ кривой показал, что в шейном отделе максимум момента сопротивления на сгибание позвоночника в вертикальной плоскости приходится на IV-V-е позвонки. Минимум момента вертикального сопротивления приходится на конец шейного отдела позвоночного столба.
Изменения моментов сопротивления позвонков силам, действующим вдоль позвоночника сибирского эласмотерия (Elasmotherium sibiricum).
В грудном отделе прослеживается 2 пика момента вертикального сопротивления. В пределах III-IV-х позвонков обозначается первый пик, что, очевидно, обусловлено наибольшей длиной остистых отростков позвонков, служащих точками приложения силы очень развитой надостистой связки, оказывающей разгибающее действие на позвоночник. Второй пик приходится на участок VIII-IX-х позвонков, где наблюдается наибольшая высота позвоночной арки. В конце грудного отдела сопротивление постепенно падает.
На данном этапе исследования не представляется возможным делать заключение о моменте вертикального сопротивления в области поясничных позвонков, так как из этого отдела позвоночника известен всего один позвонок. Тем не менее можно сказать, что момент сопротивления в вертикальной плоскости этого отдела позвоночного столба был выше, чем в конце шейного отдела. Вероятно, это можно объяснить особенностями локомоции эласмотериев как обитателя открытых ландшафтов.
Кривая моментов сопротивления поперечным силам, действующим на позвоночник, повторяет в общем кривую моментов вертикального сопротивления. Это свидетельствует в пользу синхронного усиления жесткости позвоночника как в вертикальном, так и в горизонтальном направлениях. Исключение составляет участок I-го шейного позвонка — атланта, где момент поперечного сопротивления более чем в 2,5 раза превышает момент вертикального сопротивления. Безусловно, это связано с невероятным развитием крыльев атланта.
Приведенное выше исследование особенностей строения позвоночного столба эласмотериев позволяет высказать мнение, что такая жесткая арочная конструкция позвоночника могла принадлежать животным, которые передвигались по твердым грунтам. Не противоречат этому и особенности строения конечностей эласмотериев.
Тафономические данные
Наконец обратимся к тафономическим данным, пригодным для аутэкологической интерпретации, так как они дополняют рассуждения об условиях существования и образе жизни эласмотериев, построенные на функциональном и экологическом анализе особенностей их черепа, конечностей и позвоночника.
Оба скелета сибирского эласмотерия, обнаруженные на Ставрополье, залегали in situ в палево-желтых суглинках. Как известно, палевый цвет пород является экологическим показателем формирования осадконакопления в условиях засушливых степей. Отложения, вмещающие скелет эласмотерия в ст. Гаевской, представлены не только палево-желтыми суглинками, но и серыми песчанистыми глинами с пятнами ожелезнения.
Положение скелета сибирского эласмотерия (Elasmotherium sibiricum) в раскопе у ст. Гаевской, Ставропольский край, Россия (справа — грудная клетка и правая передняя конечность).
Положение скелета в этом раскопе было необычным: животное как бы стояло на ногах, кости которых были в анатомическом порядке. При этом туловище находилось в наклонном состоянии, голова располагалась ниже таза. Задние конечности были сильно согнуты, передние выдвинуты вперед, дистальные концы пальцев раздвинуты в стороны. Создается впечатление, что эласмотерий, упираясь, соскальзывал по склону. Вероятно, подобная ситуация могла возникнуть, когда животное оказалось у водоема и неосторожно ступило на подмытый водой берег, который не выдержал тяжести массивного тела зверя и неожиданно обрушился. Погребение завершили песчано-глинистые отложения дна, из объятий которых эласмотерий уже не смог вырваться.
Особенности захоронения гаевского скелета наводят на размышления о том, что, по всей вероятности, эласмотериям, как и современным носорогам, в составе ландшафта необходимы были водоемы, где они могли после утомительных скитаний в поисках пищи не только утолить жажду, но и принять водные и грязевые процедуры. Кроме того, интразональные околоводные растительные сообщества расширяли пространство трофических ресурсов эласмотериев. Зачастую околоводные пространства с их топкими и илистыми грунтами становились тафономическими ловушками для эласмотериев, конечности которых не были приспособлены для передвижения по мягким, а тем более топким субстратам.
Реконструкция момента гибели сибирского эласмотерия (Elasmotherium sibiricum) из ст. Гаевской. Автор — С. Пучков, 2003 г.
Хорошая сохранность коллагена в некоторых образцах сибирского эласмотерия позволила П.А. Косинцеву с коллегами (2018 г.) сравнить содержание в них стабильных изотопов углерода и азота с таковым в остатках других животных плейстоцена. Высокое содержание изотопов 13C и 15N, обнаруженное у эласмотерия, характерно для млекопитающих, обитающих в сухих степях или пустынях. Повышенный уровень изотопа 13С также может указывать на питание растениями подсемейства лебедовые (накапливающих изотоп 13С из-за особого типа фотосинтеза). Полученные значения выходят за пределы их вариативности у других носорогов (в частности, шерстистого) и сопоставимы со значениями у некоторых популяций современного сайгака (
Saiga tatarica). Это также указывает на то, что сибирский эласмотерий мог питаться корнями, так как в нефотосинтезирующих частях растений накапливается 13С.

Остатки сибирского эласмотерия (
Elasmotherium sibiricum) из местонахождения Иргиз 1, Саратовская область, Россия: правая половина верхней челюсти (образец RMZ 02/269), щечная (A) и жевательная (B) поверхности; левый III-й верхний коренной зуб (образец PR 10), жевательная (C), язычная (D) и щечная (E) поверхности; левый I-й верхний коренной зуб (образец PR 8), жевательная (F), язычная (G) и щечная (H) поверхности; левый верхний III-й или IV-й предкоренной зуб (образец RMZ 02/75), жевательная (I), язычная (J) и щечная (K) поверхности; левый III-й нижний коренной зуб (образец PR 11), жевательная (L), язычная (M) и щечная (N) поверхности; правый II-й нижний коренной зуб (образец PR 12), жевательная (O), язычная (P) и щечная (Q) поверхности; левая ветвь нижней челюсти (образец RMZ 02/11), щечная (R) и жевательная (S) поверхности.
Дополнительную ясность в вопрос о рационе эласмотерия вносит изучение Ф. Ривалзом с коллегами (2017 г.) материала из местонахождения Иргиз 1, расположенного на берегу р. Большой Иргиз в Саратовской области, Россия. Здесь, помимо лругих представителей фауны начала позднего плейстоцена, были обнаружены остатки по меньшей мере 7 особей Elasmotherium sibiricum всех возрастов — от совсем молодых до старых. Геологический и палеонтологический контекст позволяет предположить внезапную гибель животных и быстрое захоронение их трупов. Общие показатели износа зубов эласмотериев оказались аналогичными таковым современного белого носорога, питающегося низкой травянистой растительностью. Однако анализ следов поверхностного микроизноса показал, что на последнем этапе жизни звери перешли с обычного травяного рациона на питание мягкой растительностью. Ученые объясняют необычное изменение рациона зимней бескормицей (дзудом) вследствие значительного накопления снега и/или ледяного покрова. Возникшая в результате этого недоступность травянистой растительности вынудила эласмотериев прибегнуть к кормлению листьями и ветками кустарников и деревьев, что привело к недоеданию и, в конечном итоге, к их гибели.

Схема резкого изменения рациона питания сибирского эласмотерия (Elasmotherium sibiricum) из местонахождения Иргиз 1, Саратовская область, Россия.
А.В. Шпанский (2021 г.) выдвигает против питания подземными частями растений следующие возражения. Во-первых, в условиях холодных открытых ландшафтов, в которых жил эласмотерий, добывать их было практически невозможно. Во-вторых, строение передней части морды и ног животного таково, что они не приспособлены для рытья почвы. В-третьих, длина передних конечностей и шеи позволяли голове эласмотерия в своем нижнем положении лишь с трудом достовать до поерхности земли. Все это говорит о том, что эласмотерий физически не мог питаться подземными частями растений. Исследователь полагает, что его основной пищей являлись травянистые растения.
Обобщая сказанное, на основе морфо-функционального и экологического анализа приходим к выводу, что эласмотерии — это узкоспециализированные животные, которые в процессе естественного отбора, судя по строению черепа, позвоночника и локомоторного аппарата, приобретают адаптации к освоению междуречных открытых пространств. Тип ландшафта, в котором обитали эласмотерии, — саванны, переходящие в высокотравные степи. Пространство трофических ресурсов было расширено за счет интразональных околоводных растительных сообществ. Поскольку одно пастбище не могло долгое время кормить столь крупное животное , эласмотерию неминуемо приходилось совершать дальние переходы.
Почему у столь специализированных животных развились приспособления для специфического способа расширения кормовой базы? Главная причина кроется в том, что в конце неогена в Евразии широко распространились многочисленные группы копытных (лошади, антилопы, быки и др.), плотно заполнившие экологическую нишу, обитателями которой были и эласмотерии. Столь крупным носорогам требовалось порядка 150 кг растительной массы в день, так что их конкурентная борьба за выживание обострилась. Расширение пространства ресурсов за счет использования подземных частей растений могло предоставить эласмотериям некоторое преимущество в питании перед другими травоядными животными, трофическая база которых оставалась традиционной.
Что же касается ранних предположений о полуводности эласмотериев, то они в свете всех полученных данных явно несостоятельны. Против такого образа существования свидетельствуют пропорции тела в целом и длинные ноги в частности, явно приспособленные для жизни в открытых ландшафтах.