Эволюция
В рамках подсемейства Rhinocerotinae триба Elasmotheriini является сестринской по отношению к трибе Rhinocerotini, включающей многих вымерших и всех ныне живущих носорогов. Некоторые исследователи поднимают ранг этих триб до подсемейств. Обе эволюционные линии носорогов имеют долгую независимую историю развития. По молекулярно-генетическим данным, их разделение произошло уже в палеогене. В зависимости от исследования, определенный период варьируется от среднего эоцена (около 47 млн. лет назад) до раннего олигоцена (порядка 31 млн. лет назад).
Палеонтологическая история трибы Elasmotheriini надежно документирована лишь начиная с раннего миоцена. Однако в это время она предстает как уже радиировавшая группа, что подтверждает ее палеогеновое происхождение, когда семейства риноцеротоидов окончательно размежевались в пространствах обитания и ресурсов. Болотные носороги Amynodontidae обосновались в нижней части катены, в прибрежных стациях интразональных растительных сообществ; гигантские Indricotheriidae освоили трофические ресурсы древесного яруса, а мало специализированные по сравнению с этими семействами Hyracodontidae и Rhinoceratidae разделили ресурсы нижнего яруса зональных сообществ.
Предки Elasmotheriini, вероятно, занимали стации в верхней части катены, настолько удаленные от тафономических ловушек местных базисов эрозии, что их шансы попасть в геологическую летопись были ничтожны. Можно предполагать, что они напоминали таких примитивных носорогов, как Aceratherium и Ronzotherium из олигоцена Евразии — достаточно крупных, длинноногих, с длинным клиновидным черепом. Более точно судить о предках и ближайших родственниках Elasmotheriini на сегодняшний день не представляется возможным ввиду сохраняющегося дефицита сведений о наиболее древних их представителях.
В раннем миоцене границы областей обитания радиировавших Elasmotheriini спустились по катене, где шло активное осадконакопление. Вероятность включения этих носорогов в ориктоценозы существенно возросла и в летопись попали сразу по меньшей мере 4 их рода: Gobitherium и Begertherium восточной Палеарктики и Beljaevina и Hyspanotherium — западной (впрочем, одна находка последнего есть и на севере Китая). По скудным тафономическим данным, пригодным для аутэкологической интерпретации, можно предположить, что весьма редкий в ориктоценозах Gobitherium занимал стации, приближенные к водоразделам, а Begertherium, остатки которого встречаются практически во всех местонахождениях фауны раннего и среднего миоцена Центральной Азии, обитал вблизи мест формирования русловых отложений, однако и те и другие — в сухих стациях.
Судя по геохронологии и сравнительной морфологии ранних Elasmotheriini, а также по данным палеогеографической ситуации в целом, областью происхождения трибы была Центральная Азия. Широтная унификация ландшафтов семиаридной зоны во время аридных климатических эпизодов раннего миоцена позволила Elasmotheriini распространиться в западные области Палеарктики, включая Пиренейский полуостров и север Африки. Наибольшего разнообразия Elasmotheriini достигают в среднем миоцене. В это время появляются примитивные Procoelodonta, Huaqingtherium, Кeniatherium. Сохраняют признаки примитивности среднемиоценовые Caementodon, Hispanotherium. В позднем миоцене, когда разнообразие непарнопалых повсеместно сократилось на фоне экспансии жвачных и в связи с ней, разнообразие Elasmotheriini упало незначительно и пополняется в восточной части Палеарктики более продвинутыми родами Iranotherium, Раrеlasmotherium, Sinotherium и собственно Elasmotherium, а в Северной Африке представлено реликтовым Hyspanotherium.
Носороги рода Sinotherium считались непосредственными предками Elasmotherium, пока не стал известен позднемиоценовый Elasmotherium primigenium. Таким образом, разделение родов Elasmotherium и Sinotherium могло произойти не позже начала позднего миоцена. Однако морфологически данный род остается переходным между более древними представителями трибы и Elasmotherium. Sinotherium достиг эволюционного успеха, освоив трофические ресурсы более влажных стаций, чем ранние Elasmotheriini, сохранив и усилив при этом адаптацию к высокоабразивным кормам, равно как и локомоторную специфику обитателей открытых пространств. К началу плиоцена этот род расселился по всей Центральной Азии, от северо-восточного Китая до Зайсанской впадины и Северного Кавказа, уступая по площади ареала только амфибиотным носорогам Chilotherium из подсемейства Aceratheriinae, с которыми он отчасти делил пространство обитания.
В отличие от всех других центральноазиатских носорогов, Elasmotheriini благополучно пережили глобальный кризис целостности континентальной экосистемы в финале миоцена, не испытав при этом существенных морфологических преобразований. После этого кризиса, повсеместно приведшего к вымиранию больше чем половины родов миоценовых млекопитающих (в Центральной Азии — до 3/4 родов) и нескомпенсированному сокращению биоразнообразия, Elasmotheriini на 2,5 млн. лет практически исчезли из геологической летописи, а затем, представленные только родом Elasmotherium, начали быстро осваивать территорию от Хуанхэ до Карпат, в областях преимущественного распространения степей и лесостепей, распространяясь с юга по влажным биотопам до 38 ° с. ш. и проникая по широким речным долинам до 58 ° с. ш.
Современный уровень изученности рода Elasmotherium позволяет предложить его представителей в качестве руководящих форм в биохронологии по крупным млекопитающим Восточной Европы, поскольку эволюционные уровни эласмотериев несут четкие диагностические признаки. Они жили относительно короткий промежуток времени, каждый вид соответствует только одной биозоне слонов мамонтовой линии. Для азиатской части части ареала такая работа пока остается не проделанной.
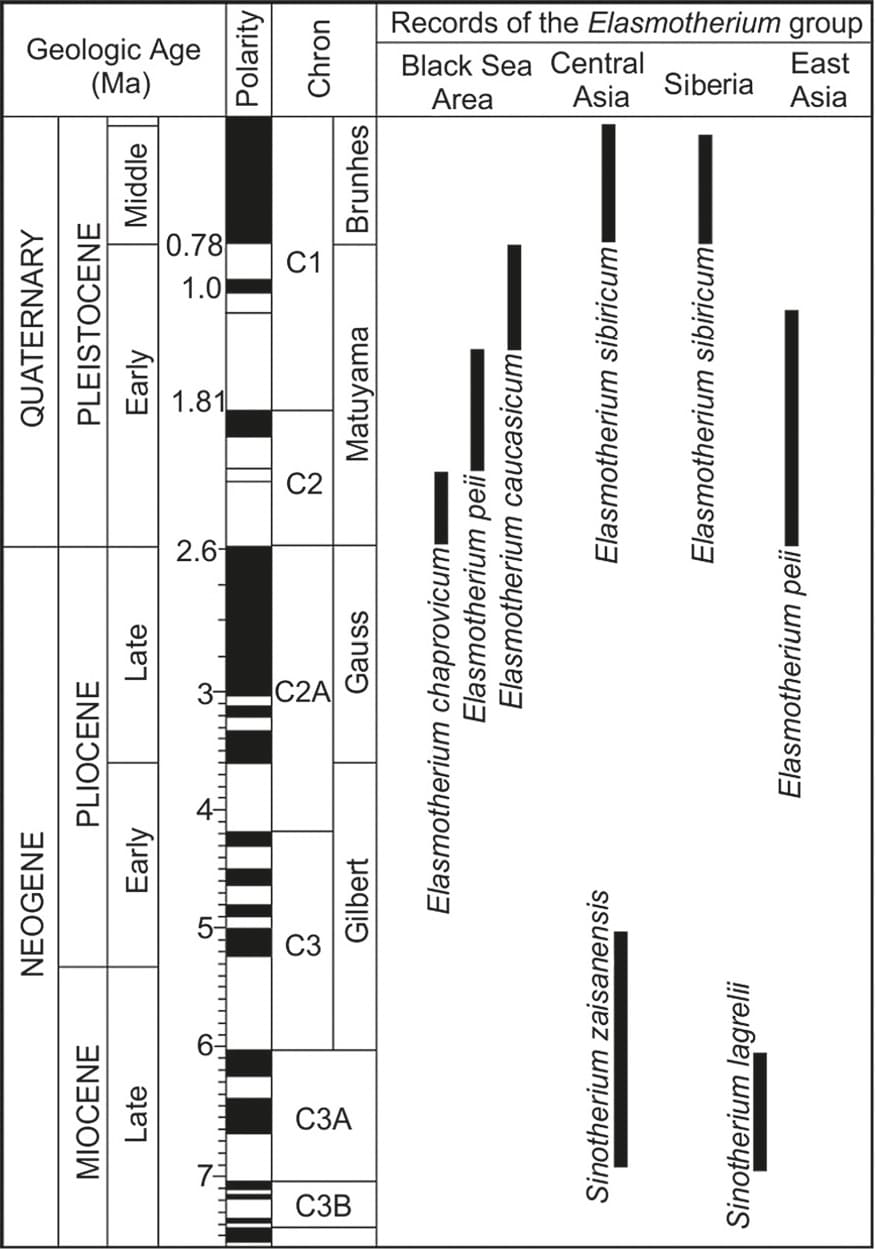
Стратиграфическое положение представителей родов Elasmotherium и Sinotherium (по Тонгу с соавт., 2018).
При сопоставлении таксономических и стратиграфических данных в процессе плейстоценовой эволюции представителей рода Elasmotherium отчетливо прослеживаются 4 этапа их расселения на Восточно-Европейской равнине. Три из них приурочены к раннему плейстоцену, и еще один обозначается в среднем плейстоцене. Каждый этап представлен собственным видом эласмотерия.
I-й этап распространения относится ко второй половине среднего акчагыла. Это время характеризуется теплым, относительно сухим климатом и распространением лесостепных ландшафтов. Эласмотерии этого времени характеризуются относительной массивностью, имеют низкий астрагал с узким блоком и широким дистальным отделом, массивные метаподии и зубы с грубой эмалью. Это архаичный Elasmotherium chaprovicum из хапровского комплекса млекопитающих. На севере Казахстана эласмотерий входил в состав одновозрастную подпуск-лебяжьинскую фауну. Отсутствие хорошо сохранившихся зубов не позволяют точно установить его видовую принадлежность, но отмечается сходство остатков с типичным Elasmotherium sibiricum.
II-й этап расселения приурочен к концу акчагыла - началу апшерона, когда проявляется тенденция аридизации и похолодания климата. Лесостепи постепенно преобразуются в открытые пространства саванного типа. Наступает эпоха Elasmotherium peii, который появился на северо-востоке Китая в начале плейстоцена и в указанное время расширил свой ареал до Причерноморья. Этот эласмотерий имеет зубы, обладающие ранним замыканием корней, длительным существованием незамкнутой задней долинки, наличием воротничка на задней стенке зуба, сильной оттянутостью назад лингвальных концов поперечных лофов, обозначением дифференциации коронки и корней.
III-й этап определяется концом раннего плейстоцена - среднеапшеронским временем, когда появляется крупнейший в роду Elasmotherium caucasicum. Аридизация и похолодание климата прогрессируют, преобладающим ландшафтом становятся открытые ландшафты типа саванн, лесостепи отступают на второй план. Для зубной системы Elasmotherium caucasicum типично наличие 3 верхних и 3 нижних предкоренных зубов. По сравнению с более древними формами зубы этого эласмотерия теряют воротничок, имеют более выраженную призмовидную форму, позднее замыкание корней и задней долинки, беспорядочное нарастание и бо́льшую извитость эмали. Судя по размерам зубов и астрагалов, от своих предков кавказский эласмотерий унаследовал крупные размеры.

Ранний плейстоцен Таманского полуострова. Кавказский эласмотерий (Elasmotherium caucasicum) бредет на водопой. Неподалеку, чтоб обезопасить себя от хищников, шествуют стеноидные лошади. Автор — Р. Евсеев
IV-й этап расселения эласмотериев прослеживается в начале среднего плейстоцена, на границе апшерона и баку, после резкого глобального похолодания. Формируется все более континентальный климат, заметнее становится сезонность. Саванноподобные ландшафты начинают преобразовываться в степные. Распространение получает Elasmotherium sibiricum, расширивший свой ареал в пределы Закавказья, Средней Азии и юга Сибири вплоть до Забайкалья. В данном случае, в противоположность прежним, азиацентричным сценариям прохореза, центр происхождения таксона, по-видимому, находился в Восточной Европе. Во всяком случае, плотность населения Elasmotherium sibiricum в западной части ареала была наиболее высокой, и время его существования здесь длилось больше, чем на востоке.
Судя по находке скелета в ст. Гаевской Ставропольского края, Elasmotherium sibiricum сохранил крупные размеры, но по организации зубной системы перешагнувший на следующий эволюционный уровень. Этот эласмотерий обнаруживает сокращение зубной формулы за счет уменьшения количества предкоренных зубов. Его коренные зубы, несмотря на достаточную стертость, имеют открытые корни, утонченную эмаль с сильной складчатостью как на верхних, так и на нижних зубах. Характерно для данного вида раннее закрытие и исчезновение задней долинки. Все зубы имеют форму слегка изогнутой призмы. В бакинское время наблюдается некоторое затухание эласмотериевой ветви. Большинство исследователей связывают этот процесс с изменением природных условий в сторону увлажнения климата.
В среднем плейстоцене, в период лихвинского межледниковья, сибирский эласмотерий вновь становится многочисленным, но поздние представители этого вида становятся более мелкими. Очевидно, аридизация среды вызвала остепнение ландшафтов. Это привело не только к постепенному сужению пространства обитания, но и сокращению пространства ресурсов. Ухудшение кормовой базы могло стать фактором, повлиявшим на изменение размеров животных. Возможно, со временем более мелкие представители плейстоценовых эласмотериев будут выделены в самостоятельный подвид.
Среднеплейстоценовые эласмотерии характеризуются усложнением рисунка эмали щечных зубов. Они приобретают еще большую глубину и хаотичность нарастания складок эмали, особенно нижних зубов. Это прослеживается, например, на находках Elasmotherium sibiricum из района г. Райгород в Поволжье, из береговых отложений р. Большой Караман на территории Саратовской области и района г. Зеленокумска Ставропольского края. Беспорядочное нарастание эмали на зубах этих носорогов, по мнению Т. Рингстрема (1924 г.), свидетельствует о деградации вида. Несмотря на значительную степень специализации животных, в хазарское время эласмотериевая ветвь постепенно угасает. Следы эласмотериев встречаются все реже и реже. К моменту развития валдайского оледенения, эта группа животных, не оставив потомков, заканчивает свое существование.

Поздний плейстоцен юга Западной Сибири. Сибирский эласмотерий (Elasmotherium sibiricum) преодолевает водораздел в поисках подходящей для пропитания растительности. Автор — Х. Ласерда, 2022 г.
На протяжении всей истории Elasmotheriini их эволюция шла в направлении развития адаптации зубной системы и органов локомоции применительно к изменяющимся условиям среды обитания. В связи с этим считаю необходимым обратить внимание на конвергентное формирование признаков в разных подотрядах непарнопалых: у Нiрроmоrрhа — Equidae, у Сеrаtоmоrрha — Elasmotheriini. Вероятно, поэтому исследователи прошлого века, пытаясь определить систематическое положение эласмотериев, ставили этих животных в непосредственном соседстве с лошадьми.
Как известно, эволюция лошадей шла в направлении их приспособления к быстрому бегу в открытых пространствах и питанию жесткой травянистой растительностью. Тенденция к монодактилизации прослеживается уже в палеогене, а в позднем плиоцене конечности лошади стали постоянно однопалыми. Зубы предков лошадей в среднем эоцене становятся лофодонтными; верхние имеют наружный гребень эктолоф и 2 поперечных — прото- и металоф; нижние строятся из 2 слабо соединенных простых полулуний. В последующее время идет дальнейшее усложнение строения зубов лошадей. Предки Elasmotheriini по меньшей мере в среднем олигоцене тоже начинают развиваться в сторону выработки монодактилизации. Их зубы также приобретают лофодонтность и в дальнейшем совершенствуются по линии усиления гипсодонтии, что получило наибольшее выражение у представителей рода Elasmotherium. По аналогии с лошадями, характер устройства жевательной поверхности зубов и высокая степень их гипсодонтии интерпретируется как ландшафтный показатель.
Таким образом, эволюция Elasmotheriini, как и лошадей, шла по пути приспособления к питанию жесткими растительными кормами и усовершенствования локомоции применительно к твердым грунтам обширных степных пространств с обильной травянистой растительностью, получающих все большее распространение в связи с усиливающейся аридизацией климата. Переход к питанию жесткой растительной пищей выразился в увеличении гипсодонтии и приспособлении к почти постоянному росту зубов с одновременным усложнением эмалевой складчастости и изменением общей формы щечных зубов. Помимо этого, совершенствование зубной системы выражается в редукции резцов и клыков, уменьшении количества предкоренных с одновременной их моляризацией. Параллельно с изменениями в зубной системе наблюдается преобразование аппарата захвата пищевого пучка и возникновение приспособлений для питания подземными частями растений. Прогресс локомоции проявился в общем удлинении конечностей в основном за счет автоподиев и среднего пальца, в редукции боковых пальцев, в отрыве I-х и II-х фаланг от субстрата (переход к копытохождению), образовании тугого замка в момент опоры в суставах кисти и стопы.
По всей видимости, своим эволюционным успехом Elasmotheriini обязаны прежде всего чрезвычайно удачному ключевому ароморфозу — снятию генетических ограничений на рост гипсодонтии, коррелятивно связанной с усилением складчатости эмали. Это обеспечило возможность постоянного расширения трофической базы за счет включения в нее все более абразивных кормов. Следующим удачным приобретением было преобразование аппарата захвата корма, сопровождавшееся полной утратой резцов и ороговением ростральной области. Такой аппарат оказался более универсальным, чем у других носорогов, хотя с точки зрения морфологии, представляется весьма специализированным. Существенным для эволюционного успеха было и сохранение архаичного (инадаптивного) локомоторного аппарата, не накладывавшего заметных ограничений на выбор пространства обитания.
Характерные для Elasmotheriini морфологические особенности, такие как очень мощное телосложение, утрата передних зубов и высококоронковые щечные зубы со сложными эмалевыми петлями, еще не проявлялись вполне у ранних представителей трибы и окончательно выкристаллизовались лишь в позднем миоцене и начале плиоцена. Сходная картина наблюдается в процессе развития плейстоценовых родов Stephanorhinus и Coelodonta из трибы Rhinocerotini. И в этом случае расширение открытых пространств привело к изменению рациона питания и, таким образом, к усилению щечных зубов, что достигает пика своего выражения у Coelodonta и, в особенности, у Elasmotherium.
Вполне возможно, что именно столь выраженная специализация поспособствовала сравнительно раннему исчезновения Elasmotherium, поскольку доступные на данный момент изотопные анализы указывают на его менее разнообразный рацион. Соответственно, животные, вероятно, были способны лишь умеренно реагировать на все усиливающиеся климатические колебания ледниковой эпохи в сочетании с изменением количества кормовых растений. Процесс вымирания был усилен низкой плотностью популяции и высокой продолжительностью смены поколений, характерной для носорогов. Нельзя также исключать, что последнюю точку в истории эласмотериев поставил человек.
Гипертрофированная реализация в эласмотериях многих характерных для носорогов эволюционных трендов, в сочетании с архаикой, создают у наблюдателя впечатление о дисгармоничности эласмотериев. Однако, эта "дисгармоничная" эволюционная стратегия оказалась действенной и спасовала только перед ландшафтно-климатическим стрессом экосистемы, вызванным рисским оледенением.
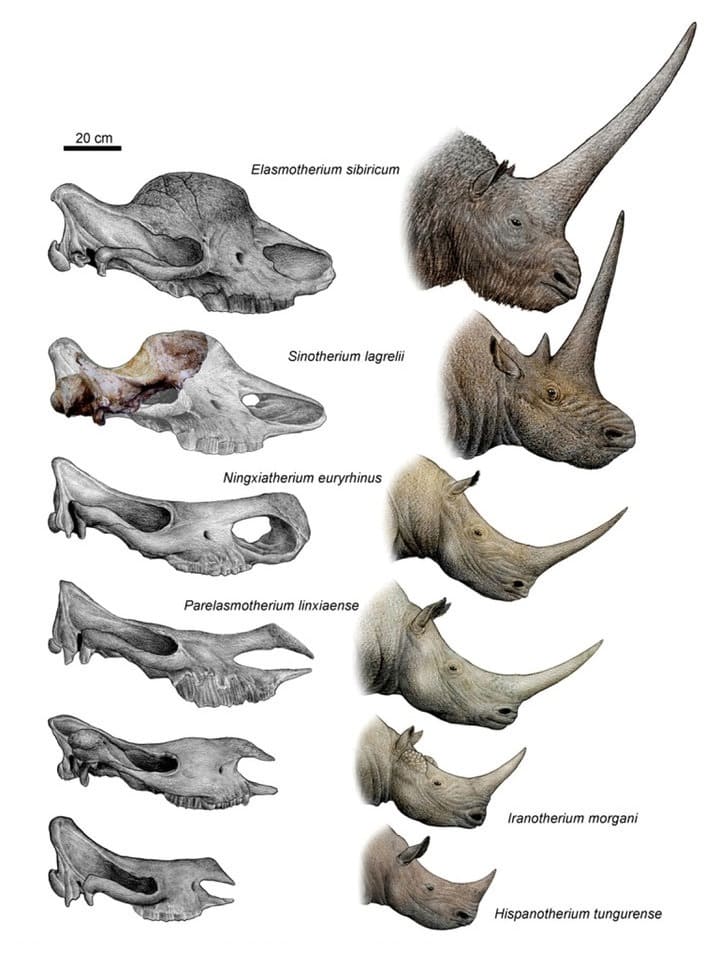
Прорисовки черепов и портретные реконструкции 6 представителей трибы Elasmotheriini, демонстрирующие изменения в строении черепов и развитие рога на лобных костях от среднего миоцена до позднего плейстоцена. Автор — Чен Ю.
Elasmotherium представляет собой заключительную стадию развития трибы Elasmotheriini и характеризуется большим куполообразным возвышением лобных костей. Долгое время оставалось неясным, как эволюционировала и выглядела при жизни эта структура и находящийся на ней рог. Большинство филогенетических предшественников Elasmotherium обладали единственным рогом на носу и прямой линией лба. Лишь у Sinotherium строение лобной области не было известно ввиду фрагментарной сохранности его черепа, но по анатомическим особенностям строения данный род являлся одним из ближайших к Elasmotherium. Благодаря открытию частичного черепа в котловине Линься в 2012 г. краниальная морфология Sinotherium стала в значительной степени известна, что облегчает понимание развития куполообразного возвышения внутри этой группы носорогов. Оно началось у сравнительно мелких ее представителей с небольшим рогом на кончике носа — таких, как Hispanotherium — в конце раннего миоцена (около 18 млн. лет назад). Окаменелости демонстрируют постепенное увеличение размеров тела животных, а также их рога посредством расширения шероховатости на месте его крепления.
В ходе эволюционного развития Elasmotheriini происходит также небольшое перемещение переднего рога, который располагался уже не непосредственно на кончике носа, а немного дальше назад, на носовых костях. Такая картина особенно характерна для Parelasmotherium, существовавшего в позднем миоцене (примерно от 10 до 5 млн. лет назад). У Sinotherium основание рога находится на месте перехода от носовой кости к лобной. Прямо за ним и немного обособленно лежит дополнительное, значительно меньшее по величине основание второго рога. Однако оба они имеют явно куполообразную структуру, как это также известно у Elasmotherium из позднего миоцена - плейстоцена. В последнем случае основание рога возвышается прямо на лбу. В ходе эволюции Elasmotherium, должно быть, произошло слияние этих двух куполообразных возвышений. На основании рога Elasmotherium можно увидеть небольшой костный шов, происхождение которого еще не объяснено, но которое, вероятно, образовалось в результате слияния двух отдельных возвышений синотериевого типа.
Параллельно смещению рога назад у Elasmotheriini происходит изменение формы носовой области. Elasmotherium имеет полностью окостеневшую носовую перегородку, как и Sinotherium, что практически неизвестно у более древних представителей трибы. Лишь позднемиоценовый Ningxiatherium имеет окостеневшую в передней трети перегородку. Неизвестно, связаны ли эти спайки с изменением положения рога. У Sinotherium обладал лобные и носовые кости стабилизировались за счет нагрузки массивного рога на многочисленные поперечные костные ребра в области неба. Помимо поздних Elasmotheriini, окостеневшие носовые перегородки встречаются только у носорогов эволюционных линий Coelodonta и Stephanorhinus, что особенно заметно у последних их представителей — Coelodonta antiquitatis и Stephanorhinus kirchbergensis.