в оглавление
назад
Энергообмен (продолжение)
Многие млекопитающие защищаются от перегрева посредством испарения воды с поверхности кожи (процесс потоотделения) и слизистых. При переходе из жидкого состояния в газообразное (пар) происходит поглощение энергии. Для испарения 1 г воды требуется около 600 кал тепловой энергии. Интенсивно выделяясь и испаряясь, пот охлаждает кожу с обширной сетью кровеносных сосудов. У человека функционирование потовых желез на ладонях находится под контролем эмоций, а на остальной поверхности тела они в норме реагируют на терморегуляцию.
Помимо кожи, излишки тепла могут отводиться через дыхательные пути. Это способ особенно характерен для млекопитающих со слаборазвитыми потовыми железами, например, для хищных. При превышении температуры окружающей среды до значений, превышающих верхний предел зоны температурного комфорта, дыхание млекопитающих становится частым и неглубоким — развивается т. н. термическая одышка, или полипноэ. Она значительно увеличивает испарение влаги с поверхности верхних дыхательных путей и, в меньшей мере, со слизистой ротовой полости и языка. При этом вдох животное производит преимущественно через нос, а выдох через рот. У собак (Canidae) теплообмен в носовых полостях в сравнении с ротовой полостью проходит более эффективно. Воду слизистой их носа поставляет особая структура — носовая железа. Все же выдох при одышке осуществляется через рот. Это связано с тем, что хотя носовые полости эффективно охлаждают воздух как при вдохе, так и при выдохе, но выдыхаемый через рот воздух выносит из организма больше тепла.
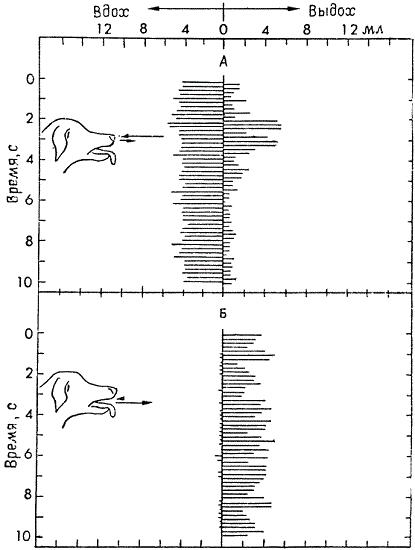
Пути прохождения воздуха при тепловой одышке у собаки (Canis lupus familiaris):
А. Поток воздуха через нос. Горизонтальными линиями, отходящими влево от вертикальной средней линии, показан вдыхаемый воздух, а отходящими вправо — выдыхаемый. Средние объемы вдыхаемого и выдыхаемого воздуха обозначены стрелками у носа собаки.
Б. Поток воздуха через рот. Обозначения те же. Вдыхание через рот практически равно нулю, но почти весь воздух, вдыхаемый через нос, выдыхается через рот.
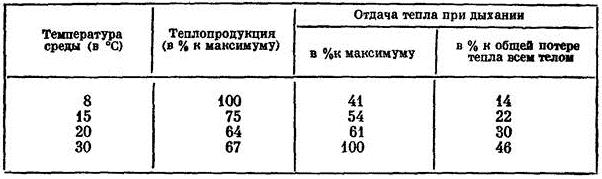
Интересно также, что при тепловой одышке собака (Canis lupus familiaris) дышит с постоянной частотой. Если в комфортных температурных условиях она совершает 15-30 дыхательных циклов в минуту, то при тепловой одышке — 300-400. Промежуточных вариантов не бывает. Периоды одышки собаки чередуют с периодами нормального дыхания. Вместе с числом совершаемых дыханий увеличивается и количество тепла, выводимого из организма. Отношение отдачи тепла при дыхании к общей его потере у собаки при температуре воздуха 8 °С составляет 14 %, при 15 °С — 22 %, при 30 °С — 46 %.
Потеря тепла у собаки (Canis lupus familiaris) при тепловой одышке (по Наумову, 1965)
При тепловых нагрузках испарение влаги через слизистые оболочки верхних дыхательных путей значительно возрастает и у жвачных жаркого климатического пояса. У коров, овец и коз тепловая одышка может иметь затяжной характер (весь жаркий день), но частота дыхания у них при этом не превышает 200 за минуту. У ежей (Erinaceus) частота дыхания достигает 240 за минуту, у мышей (Mus) она еще больше. У человека хорошо развиты потовые железы, поэтому частота дыхания даже при температуре окружающей среды 70-80 °С (температура сауны) у него составляет порядка 50-60 за 1 минуту.
Частота дыхания в зависимости от температуры среды у различных млекопитающих (по Наумову, 1989)

Оценивая значение тепловой одышки для увеличения теплоотдачи, следует учитывать, что в этом случае в первую очередь увеличивается не легочная вентиляция, а вентиляция дыхательных путей. Например, у домашнего быка (Bos primigenius taurus) при общем росте вентиляции с 50 л/мин до 300 альвеолярная вентиляция возрастает с 25 л/мин до 75, т.е. в 3 раза, а вентиляция мертвого пространства — с 25 л/мин до 225, т. е. в 9 раз. Прослеживаются и экологические особенности. Так, пустынные грызуны — кенгуровые прыгуны (Dipodomys) — теряют с дыханием 50-57 мг воды на 1 мл потребленного кислорода, а лабораторные мыши (Mus) и крысы (Rattus) — 85-94 мг. Длительная тепловая одышка приводит к акапнии — снижению содержания углекислого газа в крови и к изменению кислотно-щелочного равновесия организма (алкалозу). Существует тесная обратная связь между тепловой одышкой и потоотделением. Кроме того, изменения частоты дыхания сопровождаются изменением частоты сердечных сокращений.
У северных животных при резком понижении температуры среды дыхание становится редким, медленным, но глубоким. За счет такого изменения дыхания сокращается теплоотдача организма. При кратковременном воздействии низких температур на животных, для которых холод не является привычным фактором (песчанки, крысы, мыши), наоборот, частота дыхания повышается вследствие усиления обмена веществ и роста теплопродукции. Но при длительном пребывании в условиях низких температур (совместимых с жизнью) у этих животных, как и у северных аборигенов, дыхание со временем замедляется.
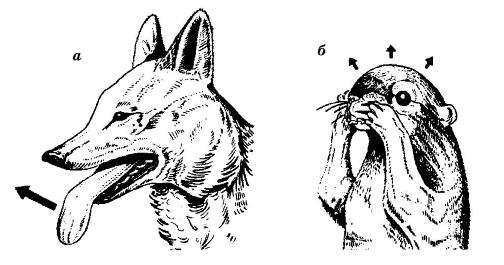
Некоторые этологические приемы использования воды для охлаждения тела: а — высунутый язык койота (Canis latrans); б — натирание головы слюной у антилопового суслика (Ammospermophilus leucurus).
Кошки (Felis), крысы (Rattus) и антилоповые суслики (Ammospermophilus) усиливают охлаждающее испарение, увлажняя поверхность своего тела слюной. У самцов отдельных видов млекопитающих дополнительно имеет место испарение влаги со слизистой полового члена — тепловая эрекция у лошадей (Equus), верблюдов (Camelus), слонов (Elephantidae). У ластоногих (Pinnipedia) механизмом увеличения теплоотдачи служит помахивание ластами. Слоны удаляют избыток тепла, помахивая увеличенными ушными раковинами с развитой сетью кровеносных сосудов и охлаждая воздухом протекающую в них кровь. Также при возможности они засасывают воду в хобот и обрызгивают себя из него. У кенгуру (Macropodidae) пот перестает выделяться практически сразу после того, как прекращается интенсивная физическая активность. Однако частота дыхания животного при этом увеличивается до 300 вдохов в минуту — в результате его разгоряченное тело остывает, не теряя слишком много влаги. Верблюды также экономят влагу, поэтому начинают потеть лишь при достижении температурой тела критического уровня. Накопленный днем избыток тепла они теряют холодной ночью.

Ватусси — порода домашнего быка (Bos primigenius taurus), выведенная в Африке. Ее отличительной особенностью являются очень длинные рога, которые пронизаны системой кровеносных сосудов и используются для терморегуляции. Кровь, циркулирующая по рогам, охлаждается потоками воздуха, и затем возвращается в тело и понижает его температуру.
Теплорегуляция также может осуществляться за счет изменения положения тела в зависимости от погоды. На поверхностную теплоизоляцию влияют ветер и влажность. Влажная шерсть повышает теплопотери за счет повышенной проводимости. Когда млекопитающее лежит или сидит на земле, его шерсть уплотняется и содержит меньше воздуха. Тепло в этом случае уходит в почву, особенно если она влажная. Часто можно видеть, как перед дождем коровы опускаются на землю для отдыха, сохраняя при этом тепло, которое было бы потеряно через некоторое время — когда земля станет влажной. Эффективную поверхность тела также можно уменьшить, свернувшись клубком.
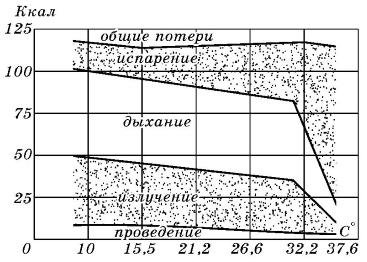
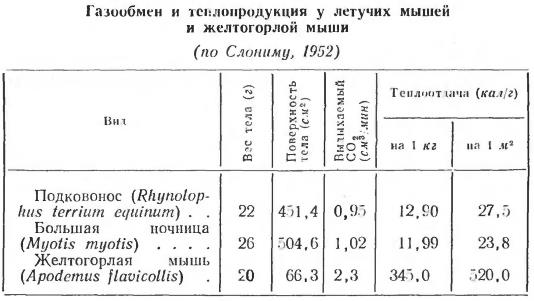
Пути выведения тепла из организма домашней свиньи (Sus scrofa domesticus) при разных температурах окружающей среды.
Весьма эффективным приемом сохранения теплового гомеостаза является простое уклонение от неблагоприятного фактора среды. С этой целью животные используют как индивидуальные локомоции, так и преимущества группового поведения. Например, свинья в жару при возможности валяется в грязи, корова уходит в тень или воду, разгоряченная собака ложится на снег, отдавая тем самым излишки своего тепла более холодной внешней среде. Стадные животные (например, овцы) в отсутствие тени, навеса собираются в группы и прячут голову в тени, которую создают соседи. Луговые собачки (Cynomys), суслики (Spermophilus), сурки (Marmota) и другие роющие животные в жаркую погоду периодически прячутся в своих прохладных норах, где перегретое тело может охлаждаться путем теплоизлучения. Сбивание в кучу с другими особями собственного вида снижает теплопотери. Овцы в снежную бурю образуют плотные скопления и таким образом защищаются от ветра и снега.
Более или менее значительно снижается температура тела у млекопитающих, впадающих в сезонную спячку или глубокое оцепенение. Для животных это выгодно, поскольку пропорционально уменьшаются и энергетические затраты организма. Сходное явление может наблюдаться и при суточном снижении активности, например, у тенреков (Tenrecidae) и рукокрылых во время дневного сна. Значительная поверхность крыльев последних обусловливает существенные потери тепла, а поскольку они не способны поддерживать достаточную температуру тела во время отдыха, то снижают ее. Нередко эти зверьки так сильно охлаждаются, что после пробуждения должны некоторое время делать своего рода разминку, прежде чем отправиться в полет. Впрочем, даже у бодрствующих рукокрылых температура большей части тела относительно низка и почти совпадает с температурой окружающей среды. Также температура тела млекопитающих регулируется путем перемены местообитаний и совершения миграций.
Для подавляющего большинства видов животных в условиях температурного комфорта основным местом испарения воды при терморегуляции является все-таки кожа. По имеющимся данным, лактирующая корова живой массой от 300 до 800 кг испаряет через кожу от 6 до 16 л воды. На испарение через кожу у лошади приходится 5-8 л, у взрослой свиньи — 2,0-2,5 л, у стриженной овцы — около 2,5 л воды. Таким образом, суточная теплоотдача за счет испарения воды через кожу у коровы доходит до 9600 ккал, у лошади — до 4800 ккал, а у свиньи и овцы она составляет от 1200 до 1500 ккал.
Очевидно, что путь теплоотдачи, который используется организмом животного, определяется силой теплового фактора. О влиянии температуры среды на отведение тепла из организма домашней свиньи приводятся следующие данные. До температуры воздуха 30 °С основным способом теплоотдачи у свиньи является дыхание и излучение. Когда разница между температурой тела животного и температурой окружающей среды становится минимальной или вовсе пропадает, излучение тепла прекращается. Главным способом избавления от избыточного тепла становится испарение. У свиней важную роль в терморегуляции играют конечности, уши и хвост. Интересно, что при температуре окружающей среды 5 °С температура выступающих частей тела у животного существенно ниже, чем при температуре 25 °С. Так, температура ушей при 5 °С составляет всего 15 °С, при температуре воздуха 15 °С температура ушей поднималась до 27 °С, при 25 °С уши нагреваются до температуры 36 °С. Аналогичные изменения происходят и с температурой хвоста свиньи. В целом за счет изменения кровоснабжения разных участков кожи у свиньи суммарные теплопотери с поверхности тела в неблагоприятных условиях изменяются в пределах 70 %.
Изменения температуры поверхности разных частей тела домашней свиньи (Sus scrofa domesticus) при разных значениях температуры окружающей среды.
Приспосабливаясь к температурным условиям среды, животные подпадают под ряд экогеографических закономерностей.
Согласно правилу Бергмана, в пределах одного вида гомойотермных животных более крупными являются формы, обитающие в условиях более холодного климата — в высоких широтах или в горах. Кроме того, если близкие виды существенно не отличаются по характеру питания и образу жизни, то более крупные из них также встречаются в условиях более холодного климата. Данное правило основывается на предположении, что общая теплопродукция у эндотермных видов зависит от объема тела, а скорость теплоотдачи — от площади его поверхности. Действительно, выделение тепла через поверхность тела составляет приблизительно 2/3 тепловых потерь организма, и чем поверхность тела больше относительно его объема, тем больше потеря тепла. В свою очередь, объем тела при увеличении общего размера растет быстрее, чем поверхность тела. Это значит, что более крупные животные теряют меньше тепла и относительно (но не абсолютно) экономнее в использовании энергии, чем более мелкие. Несомненно поэтому, что крупные размеры выгодны для животных, обитающих в суровых климатических условиях, где большие потери собственного тепла восполняются труднее, чем в условиях теплого климата. Наоборот, в умеренном и жарком климате мелкие и средние животные имеют преимущества перед своими более крупными собратьями.
Правило Бергмана часто выполняется. Так, подвиды тигра из островной части Индонезии — суматранский (Panthera tigris sumatrae), яванский (Panthera tigris sondaica), балийский (Panthera tigris balica) — весят всего 65-140 кг, тогда как масса бенгальского (Panthera tigris tigris) и амурского (Panthera tigris altaica) тигров составляет 100-225 кг. Европейский бурый медведь (Ursus arctos arctos) в среднем весит от 150 до 300 кг, камчатский медведь (Ursus arctos beringianus) — от 200 до 500 кг, а медведи Аляски (Ursus arctos gyas) и острова Кадьяк (Ursus arctos middendorffi) имеют массу от 200 до 600 кг. (Во всех приведенных примерах нижний предел массы относится к более мелким самкам, а некоторые особенно крупные самцы далеко выходят за верхнюю границу.)
Далее, южные подвиды волка (Canis lupus) в среднем также мельче, чем северные. На территории Евразии рыжая лисица (Vulpes vulpes), заяц-беляк (Lepus timidus) и сибирская косуля (Capreolus pygargus) мельчают в юго-западном направлении и, напротив, увеличиваются в размерах к северу и востоку, где зимы суровее. Дикие кабаны (Sus scrofa) из Южной Испании имеют черепа длиной в среднем 32 см, в Польше — около 41 см, в Беларуси — 46, а в Сибири встречаются очень крупные особи с длиной черепа 56 см. Закономерность работает и в отношении домашних животных. К примеру, беспородные собаки в Краснодарском крае, крупный рогатый скот местной селекции отличаются меньшей живой массой по сравнению с представителями этих видов, скажем, Архангельска.

Относительные размеры различных подвидов тигра (Panthera tigris).
Если две относительно близко расположенные территории имеют сходные фауны, но различаются средними показателями температуры, т. е. одна из них холоднее, то именно на ней средние размеры млекопитающих окажутся больше. К примеру, на южном побережье Австралии среднегодовая температура составляет 16 °C, а на побережье Тасмании — 11 °C. Этого достаточно, чтобы тасманийские ехидна (Tachyglossus aculeatus), утконос (Ornithorhynchus anatinus) и кенгуру были крупнее своих австралийских сородичей.
В то же время данная закономерность не может иметь общего характера, поскольку помимо температуры на размеры животных влияют многие другие факторы. Кроме того, адаптации к суровому климату на популяционном и видовом уровне часто происходят не за счет изменений размеров тела, а за счет изменений размеров внутренних органов (увеличение размера сердца и легких) или за счет биохимических адаптаций. Поэтому необходимо подчеркнуть, что правило Бергмана носит статистический характер и отчетливо проявляет свое действие только при прочих равных условиях.
Действительно, из правила Бергмана известно немало исключений, которые можно понять и объяснить в каждом конкретном случае. Так, наиболее мелкая форма шерстистого мамонта, Mammuthus primigenius vrangeliensis, известна с заполярного острова Врангеля. Ее измельчание, несомненно, объясняется скудностью кормовой базы на этом клочке северной суши. Многие лесные подвиды волка крупнее, нежели тундровые — например, исчезнувший Canis lupus alces с полуострова Кенай на юге Аляски. Предполагается, что крупные размеры могли давать этим волкам преимущество при охоте на крупных лосей, населяющих полуостров. Дальневосточный подвид леопарда (Panthera pardus orientalis) существенно меньше, чем африканский (Panthera pardus pardus). По-видимому, это связано с тем, что копытных, служащих основной пищей этой кошке, в тайге гораздо меньше, чем в саванне.
Другой пример представляют мелкие млекопитающие арктического и субарктического регионов. Здешние полевки, мыши и землеройки, большую часть времени проводящие в специфическом микроклимате нор, более или менее стабильном и нередко более мягком, чем климат окружающей местности. Активные зимой под слоем снега, они находятся в условиях, существенно отличающихся от тех, которые царят над заснеженной равниной, поскольку снег оказывает значительный теплоизоляционный эффект. В центральной части Аляски было исследовано распределение температуры над снегом и под ним. Снежный покров при этом был относительно тонок — 60 см. Термометр показывал -50 °C, а под слоем снега на поверхности почвы мороз не достигал и -7 °C. В этих условиях серые полевки (Microtus) ведут активную жизнь и свободно передвигаются в своих подснежных ходах, хотя их меховой покров тонок, а ступни на лапах вообще не покрыты шерстью. В то же самое время северные олени с большими трудностями переживают эти тяжелые холода. Таким образом, можно сказать, что эти млекопитающие, находясь в одной и той же географической точке, существуют в совершенно различных климатических условиях.

Сравнение размера ушных раковин шерстистого мамонта (Mammuthus primigenius), саванного (Loxodonta africana) и азиатского (Elephas maximus) слонов. Автор — З. Буриан.
Следующая экогеографическая закономерность — правило Аллена — гласит, что среди родственных форм гомойотермных животных, ведущих сходный образ жизни, те, которые обитают в более холодном климате, имеют относительно меньшие выступающие части тела: уши, морду, конечности, хвост и т. д. Это приводит к уменьшению относительной поверхности тела и, соответственно, способствует экономии тепла. Кроме того, у северных и высокогорных животных указанные части тела покрыты густой шерстью, вследствие чего кажутся еще более короткими. Наоборот, населяющие жаркий пояс животные склонны обладать особенно большой относительной поверхностью тела, рассеивающей избыток тепла. Примечательно, что данная закономерность распространяется не только на диких, но и на домашних животных.
В качестве типичного примера действия правила Аллена приведем отношение длины хвоста к длине тела у темной полевки (Microtus agrestis) из различных районов Европы: Швеция — 29 %, Центральная Европа — 33 %, Португалия — 39 %. Закономерность подтверждается и при межвидовых сравнениях. Так, у арктического беляка (Lepus arcticus) из Гренландии длина ушей достигает только 96 % длины головы, тогда как у белобокого зайца (Lepus callotis) в Мексике — целых 189 %.
Большие уши, недопустимые в условиях низких температур, возникли как приспособление к жизни в жарком поясе. В пустынях южной части североамериканского континента живет антилоповый заяц (Lepus alleni), на голые ушные раковины которого приходится 25 % поверхности тела. Их развитая кровеносная система с уникальной сосудодвигательной способностью обслуживает терморегуляцию. За счет усиления и ограничения кровотока через ушные раковины животное меняет теплоотдачу на 200-300 %. Помимо того, что уши антилопового зайца выполняют функции поддержания теплового гомеостаза и экономии воды, они очень чувствительны к звукам.
Почти столь же показательны ушные раковины саванного слона (Loxodonta africana), площадь которых составляет 1/6 часть поверхности всего тела животного. Как и уши антилопового зайца, они голые, насыщенны термочувствительными нервными окончаниями и сильно васкуляризованны. Благодаря сосудодвигательным реакциям в жаркую погоду через кровеносную систему ушных раковин слона проходит примерно 1/3 часть всей циркулирующей крови. В результате усиленного кровотока во внешнюю среду отдается излишняя тепловая энергия. Наоборот, у шерстистого мамонта (Mammuthus primigenius) уши имели небольшую площадь и к тому же были покрыты густой шерстью. По сравнению с современными слонами, у мамонта были относительно короткие конечности и хобот.
Не менее наглядно сравнение песца (Vulpes lagopus), рыжей лисицы (Vulpes vulpes) и фенека (Vulpes zerda). Маленькие уши песца сохраняют тепло, а огромные уши фенека рассеивают его избыток. Морда песца короткая, а у фенека она узкая и вытянутая. Песец отличается приземистой осанкой и компактной формой тела, а фенек относительно выше и изящнее. Лисица по степени развития указанных признаков занимает между этими двумя видами промежуточное положение.
Уменьшение длины ушей и морды у лисиц при продвижении на север: 1 — песец (Vulpes lagopus); 2 — рыжая лисица (Vulpes vulpes); 3 — фенек (Vulpes zerda).
Длинные конечности невыгодны в условиях низких температур, так как с их поверхности теряется слишком много тепловой энергии. Но в условиях жаркого климата длинные конечности являются полезной адаптацией. При сравнении северного оленя (Rangifer tarandus) с благородным (Cervus elaphus) ясно видно, что уши у первого короче и он более коротконог. Весьма длинны конечности и шея у жирафа (Giraffa camelopardalis) и геренука (Litocranius walleri), которые обитают в саваннах Африки. В пустынных условиях верблюды, козы, лошади местной селекции, а также овцы, кошки, как правило, тоже длинноноги. Рассматриваемая закономерность касается и рукокрылых. Относительно более крупные крыланы (Megachiroptera) имеют огромную поверхность крыльев и распространены только в тропической зоне, тогда как мелкие летучие мыши (Microchiroptera) смогли расселиться вплоть до умеренных широт.
Лабораторные опыты также подтверждают оба правила. Домовые мыши (Mus musculus), содержавшиеся с самого раннего возраста при пониженной температуре среды, равной 6 °C, вырастали значительно крупнее тех, которые в тот же период времени находились в условиях средней температуры в 26 °C. Кроме того, у мышей, выращенных в холоде, уши и ступни оказались короче, а у выращенных при повышенной температуре — длиннее нормальных. Известно также, что в промышленных холодильниках живут наиболее короткохвостые мыши. Укорачиваются хвосты и у особей, которым в условиях эксперимента регулярно давали препараты, снижающие температуру тела. Значительная вариативность морфологических показателей, обусловленная влиянием факторов внешней среды, свидетельствует о том, что не все признаки, подчиняющиеся климатическим правилам, изменяются под действием отбора. Зачастую наблюдаются лишь ненаследственные изменения — модификации.
Постоянная температура тела поддерживается за счет окисления органических соединений, поступающих в организм с кормом. Следовательно, при большей энергетической отдаче необходимо увеличивать объем принимаемой пищи. Величина энергообмена у крупного копытного (при массе приблизительно 500 кг) составляют 50 кДж на 1 кг живого веса, но у обезьяны (масса 4,2 кг) — уже 206 кДж/кг, у мыши (0,021 кг) — 714 кДж/кг, а у бурозубки (0,0035 кг) — даже 3486 кДж/кг. Поэтому неудивительно, что дневной рацион тигра составляет 3-4 % массы его тела, небольших грызунов — около 100 %, а наиболее мелких землероек — даже до 200 % их массы. Насытившись, тигр (Panthera tigris) подолгу отдыхает, а этрусская землеройка (Suncus etruscus) вынуждена почти безостановочно питаться, чтобы не погибнуть от голода. Сохранение энергии очень важно и для морских млекопитающих. Для того чтобы максимально использовать преимущества соотношения поверхности и объема тела, многие из них достигают просто грандиозных размеров. Так, у синего кита (Balaenoptera musculus) соотношение между поверхностью тела и его объемом в 10 раз выгоднее, чем у небольшой морской свиньи (Phocoena phocoena). Это, в сочетании с толстым слоем подкожного жира, дает полосатику 100-кратное преимущество в отношении теплопотерь в холодной воде.
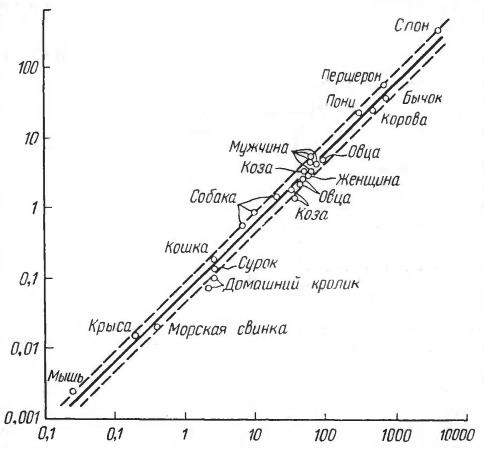
Соотношения между пульсом, уровнем обмена и величиной тела у млекопитающих. По оси ординат — отношение потребления кислорода (мл) к частоте пульса; по оси абсцисс — вес тела (кг).
Итак, у мелких животных соотношение между объемом тела и его теплоотдающей поверхностью в целом менее выгодно, чем у крупных. С другой стороны, критическим результатом увеличения площади поверхности (и, следовательно, теплоотдачи) относительно массы (и объема) по мере уменьшения тела является резкое возрастание потребления энергии. Судя по всему, самые маленькие бурозубки находятся на нижней размерной границе млекопитающих. Еще более мелкие зверьки, вероятнее всего, просто погибли бы, так как оказались бы не в состоянии восполнять огромные энергетические потери достаточным количеством пищи.
При прочих равных условиях энергозатратный обмен веществ мелких млекопитающих вынуждает их потреблять больше пищи и быстрее ее переваривать в сравнении с более крупными зверями. Однако условия редко бывают равными, поскольку пища различается по качеству и содержанию энергии. Животная ткань, ягоды, орехи и клубни очень богаты легко усвояемой энергией, в отличие от большинства растительных тканей, где содержимое каждой клетки заключено в толстую оболочку неперевариваемой (в отсутствие специальных микроорганизмов) клетчатки. Кусок мяса содержит не только больше энергии в сравнении с листом, вдобавок он еще и легче переваривается. Поэтому хищная ласка (Mustela nivalis) в 26 раз эффективнее в извлечении энергии из пищи, чем ее растительноядная добыча — обыкновенная полевка (Microtus arvalis). Более мелкие представители любого отряда млекопитающих обычно поддерживают свои энергетические потребности за счет пищи, более богатой питательными веществами, в сравнении с крупными животными. Так, весящий 4 кг голубой дукер (Philantomba monticola) выбирает почки и молодые ростки, тогда как имеющая массу до 1 т канна (Taurotragus oryx) способна выжить па пастбище с редкой травянистой растительностью; мелкий сенегальский галаго (Galago senegalensis) поедает плоды, а очень крупная восточная горилла (Gorilla beringei) — листья.
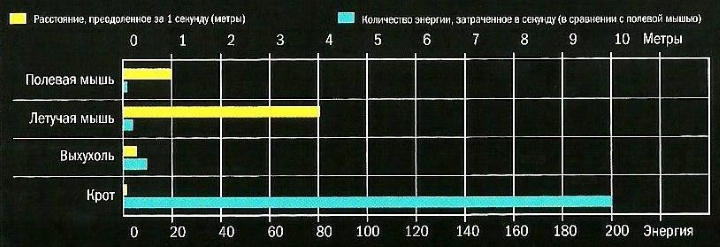
Эффективность использования энергии четырьмя млекопитающими сходной массы (около 50 г): бегущей по земле полевой мыши, летящей летучей мыши, плывущей в воде выхухоли и движущегося в почве крота. Полосы диаграммы демонстрируют, как далеко каждый из них продвинулся за 1 секунду и сколько энергии потратил (энергия полевой мыши принята за единицу). Летучая мышь использует энергию наиболее эффективно, а крот наименее.
У животных, потребляющих малопитательные растительные корма или сталкивающихся с сезонным минимумом пищевых ресурсов, обмен веществ зачастую понижен. Например, энергетический бюджет орангутана (Pongo pygmaeus) составляет 70 % от ожидаемого значения энергозатрат для млекопитающего с соответствующей массой тела, коала (Phascolarctos cinereus) — 69 %, кошачьего лемура (Lemur catta) — 52 %, большой панды (Ailuropoda melanoleuca) — 38 %, трехпалого ленивца (Bradypus tridactylus) — 36 %. Установлено также, что звери с пониженным метаболизмом тратят меньше ресурсов и на поддержание самых дорогостоящих в энергетическом плане органов — мозга, печени и почек. Эти органы у них меньше, чем можно было бы ожидать исходя из их массы тела и на основании данных для других плацентарных. Например, масса головного мозга большой панды в среднем составляет 82,5 % от ожидаемого, масса печени — 62,8%, почек — 74,5%. Кроме этого, панда экономит энергию, двигаясь меньше и медленнее, чем другие медведи. По сравнению с млекопитающими сходных размеров, мышечная масса ленивцев уменьшена наполовину. Большую часть времени они спят, передвигаются крайне медленно, а съеденная ими пища может перевариваться в течение недели.
Различные виды пищи различаются своей доступностью. «Высококачественные» компоненты не столь обильны, как «низкокачественные». В целом обилие пищи, доступной для представителей какого-либо вида, зависит от того, какое место в пищевой цепочке они занимают: в каждом фрагменте этой цепочки часть энергии теряется и она становится все менее доступной для животных, находящихся в конце цепочки. Вот почему суммарный вес (биомасса) хищников меньше, чем их жертв, а масса растительноядных животных меньше, чем растений, которые являются первичными переработчиками энергии солнца в потребляемую животными форму.
Таким образом, более мелкие животные требуют больше энергии и поэтому потребляют лучше усвояемую пищу, тогда как крупные могут продержаться и на менее калорийных, но более доступных кормах. Многие виды игнорируют это правило благодаря освоению особого рациона: например, любители муравьев и термитов, древесные листоеды или летающие пожиратели насекомых характеризуются менее интенсивным метаболизмом, чем другие млекопитающие сходных с ними размеров. С другой стороны, у небольших дельфинов, ластоногих и выдр, ведущих очень энергозатратный образ жизни, скорость обмена веществ даже выше, чем это следует из их размеров.

У одной группы японских макак (Macaca fuscata) сформировалась привычка принимать ванны в термальных источниках, расположенных на их территориальном участке. Когда температура воздуха опускается ниже 15 °C, обезьяны укрываются от холода в теплой воде.