История открытия



Смонтированный скелет гигантского короткомордого медведя (
Arctodus simus) с ранчо Ла-Бреа в штате Калифорния, США. На заднем плане — смонтированный скелет западной лошади (
Equus occidentalis) оттуда же. Музей Пейджа, Лос-Анджелес.
Таксон Arctotherium simum был установлен в 1879 г. Эдвардом Дринкером Копом по почти полному черепу, найденному в 1878 г. Дж. А. Ричардсоном в позднеплейстоценовых отложениях пещеры Поттер-Крик на территории округа Шаста, штат Калифорния, США. Его видовой эпитет переводится как "курносый", намекая на короткую морду зверя. В 1902 г. Оливер Пэрри Хэй определяет этот вид в род Arctodus. В дальнейшем было описано несколько синонимических таксонов. В 1911 г. Джон Кэмпбелл Мерриам по костям кисти из позднего плейстоцена ранчо Ла-Бреа в Калифорнии устанавливает Arctotherium californicum. В том же году Лоуренс Моррис Ламб по позднеплейстоценовому черепу из Голд-Ран-Крик на Юконе в Канаде описывает Arctotherium yukonense. В 1916 г. Эрвин Хинкли Барбур по левой плечевой кости из Касса в штате Небраска, устанавливает Dinarctotherium merriami. Наиболее полно сохранившийся скелет гигантского короткомордого медведя был обнаружен в округе Фултон, штат Индиана.

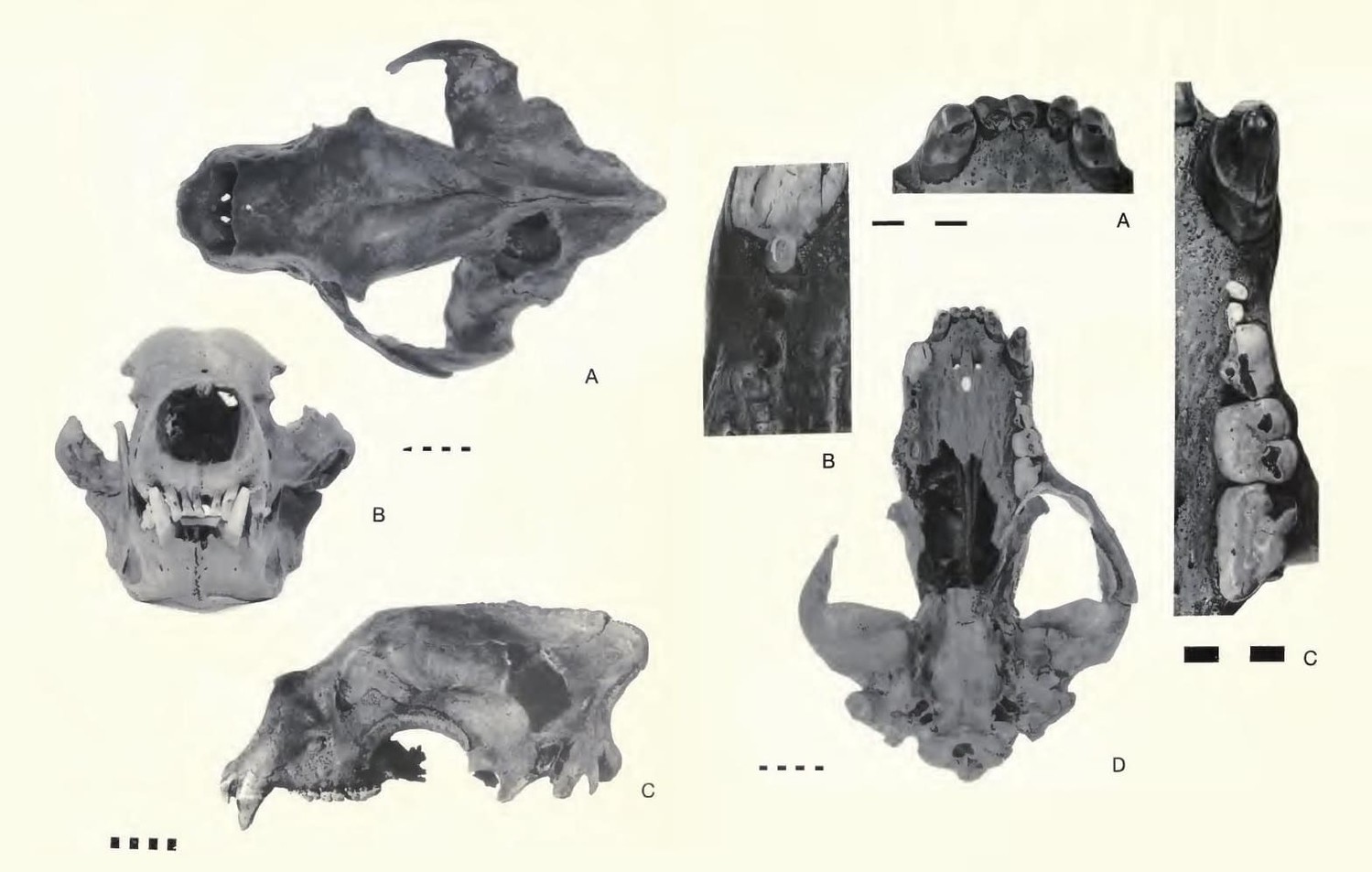
Череп с нижней челюстью гигантского короткомордого медведя (Arctodus simus) и детали его строения. Масштабные линейки в сантиметрах.
Строение и функции
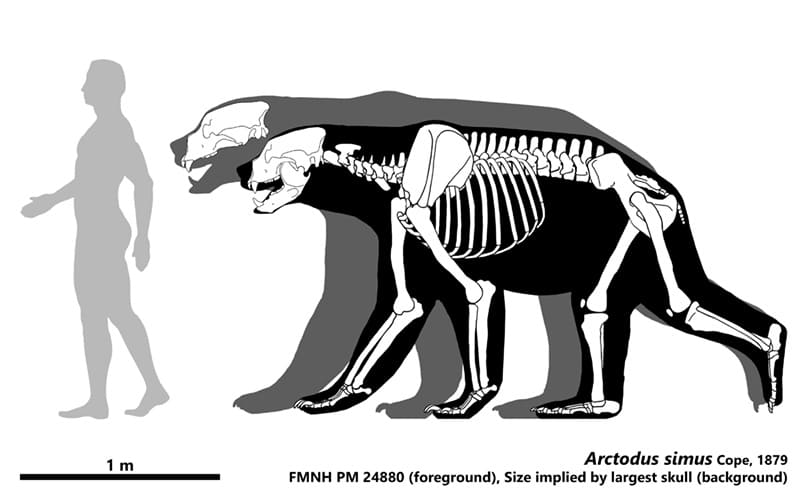
Гигантский короткомордый медведь был самым большим наземным млекопитающим хищником, когда-либо живших в Северной Америке. Судя по размерам найденных остатков, крупнейшие его особи достигали около 1,6 м в холке, а когда стояли на задних лапах, их рост составлял порядка 3,4 м. Однако в пещере Риверблафф в штате Миссури серия следов когтей, предположительно принадлежащих этому зверю, расположена вдоль стены на высоте до 4,6 м, что указывает на еще больший его рост — порядка 3,7 м. Масса тела взрослых животных, рассчитанная исходя из размеров элементов скелета, обычно варьировала в пределах от 300 до 800 кг, достигая порой 1000 кг. Как и у других медведей, ярко выражен половой диморфизм: самцы в среднем весили порядка 600 кг, тогда как самки — до 400 кг. В целом, как показывают расчеты, самцы по меньшей мере на 15 % превосходили самок линейно и были на 25 % тяжелее (примечательно, что у очкового медведя (Tremarctos ornatus) последний показатель еще выше, достигая 40 %).

Реконструкция внешнего облика гигантского короткомордого медведя (Arctodus simus). Автор — К. Бьюэлл.
Помимо половой принадлежности, средние размеры варьируют в зависимости от временно́го и географического факторов. Так, крупнейшие особи (вероятно, в основном взрослые самцы, поскольку оттуда же известны особи примерно на 1/4 мельче) из битумных ям ранчо Ла-Бреа в Калифорнии (ранчолабреаний) и ряда других районов заметно уступают по величине большинству экземпляров с Аляски, Юкона, Небраски, Юты и Калифорнии (ирвингтоний). На основании этого указывалось на присутствие экоморфологических различий и даже предлагалось выделение двух подвидов — крупного
Arctodus simus yukonensis, занимавшего северную и центральную части ареала, и более мелкого
Arctodus simus simus, встречавшегося на его западе и юге. Однако разделение по размерному критерию является недостаточно обоснованным, поскольку известны как достаточно крупные медведи из Флориды, так и относительно мелкие с Юкона, а также наблюдается широкий вариационный размах размеров в одном местонахождении. Несостоятельность выделения
Arctodus simus simus и
Arctodus simus yukonensis подтвердило генетическое исследование, не обнаружившее никакой разницы между ними. Судя по всему, приписываемые им размерные различия на самом деле были проявлением полового диморфизма.
Прорисовка скелета крупной особи гигантского короткомордого медведя (Arctodus simus), вписанного в телесные очертания (на переднем плане), и телесный силуэт, реконструированный на основе крупнейшего известного черепа данного животного (на заднем плане).
Согласно современным данным, в среднем плейстоцене (ирвингтоний) на территории западной и центральной частей США встречались крупные короткомордые медведи. В позднем плейстоцене (ранчолабреаний) звери южных популяций стали мельчать, хотя среди них еще попадались крупные особи. На Аляске и у кромки Канадского ледникового щита, напротив, преобладали крупные короткомордые медведи. Сохранению крупных размеров, вероятно, способствовали северное распространение (т. н. правило Бергмана о уменьшении млекопитающих в направлении с севера на юг) и конкуренция со стороны встречавшихся здесь бурых медведей.
Из других представителей семейства медвежьих гигантского короткомордого медведя по размерам тела превосходит только арктотерией (Arctotherium angustidens) из раннего плейстоцена Южной Америки. Его расчетная масса колеблется в пределах 400-1200 кг, а для одного исключительно крупного самца составила порядка 1600 кг. Однако большинство особей указанных двух видов вполне сопоставимы по величине друг с другом. Помимо крупных размеров тела, представляющих собой пример эволюционного параллелизма, между этими медведями имеются заметные морфологические различия.
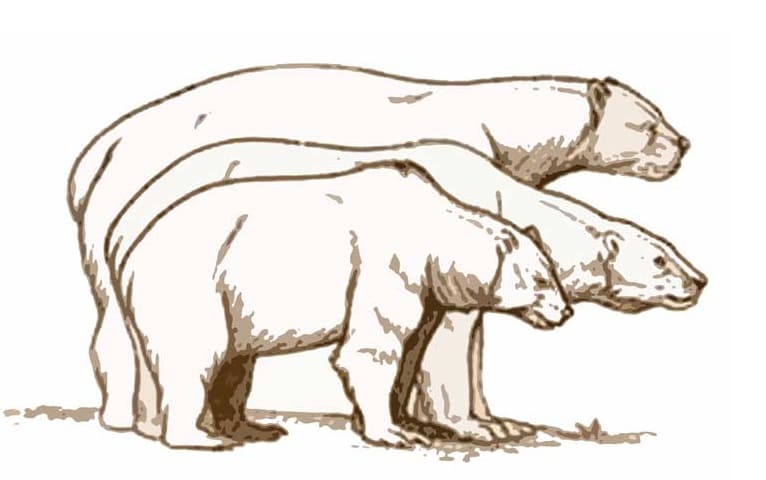
Относительные размеры, спереди назад: бурый медведь (Ursus arctos), белый медведь (Ursus maritimus), крупная особь гигантского короткомордого медведя (Arctodus simus).
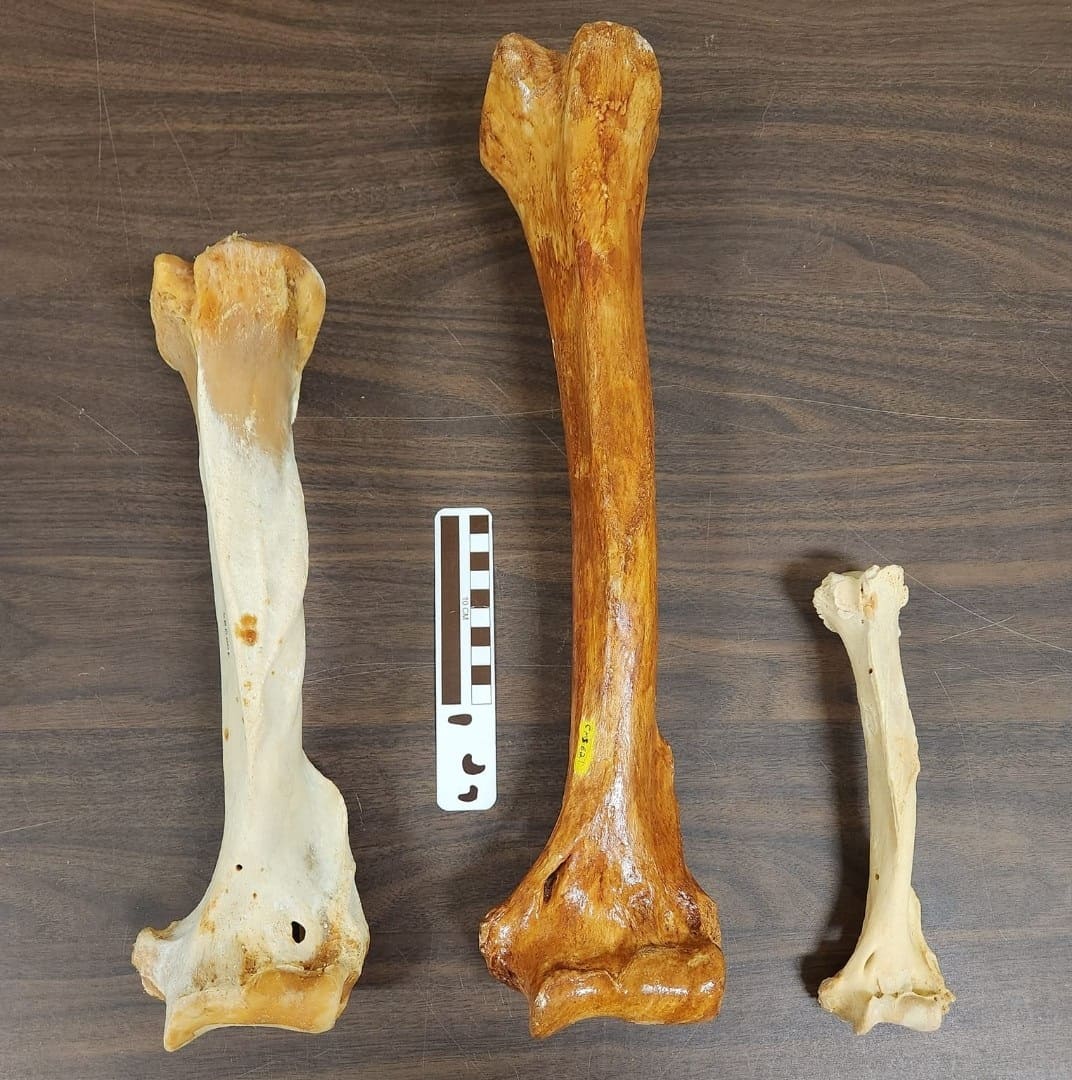
Самый большой из ныне живущих белых медведей (Ursus maritimus) весил порядка 800 кг. При этом средняя масса самца этого вида составляет 400-450 кг, самки — 350-380 кг. Крупные самцы имеют рост в холке 130-150 см. Примерно таких же габаритов достигают крупные самцы бурого медведя (Ursus arctos). Вариационный размах длины костей конечностей гигантского короткомордого и бурого медведя составляют соответственно: плечевая кость — 49-58 см и 25-42 см, бедренная кость — 56,5-66 см и 30-50 см, таранная кость — 6,8-9,1 см и 3,8-6,2 см; при этом следует учитывать различия в телесных пропорциях обоих видов.
Таким образом, как по максимальным, так и по средним размерно-весовым показателям гигантские короткомордые медведи несколько превосходят своих современных родственников. Впрочем такая ситуация наблюдается не всегда: например, гигантские короткомордые медведи из Ла-Бреа (в среднем весившие около 370 кг) были более мелкими, чем жившие там же бурые медведи (в среднем массой примерно 450 кг). Согласно одному из предположений, остатки бурого медведя из упомянутого местонахождения могут датироваться более поздним периодом, чем остатки гигансткого короткомордого.
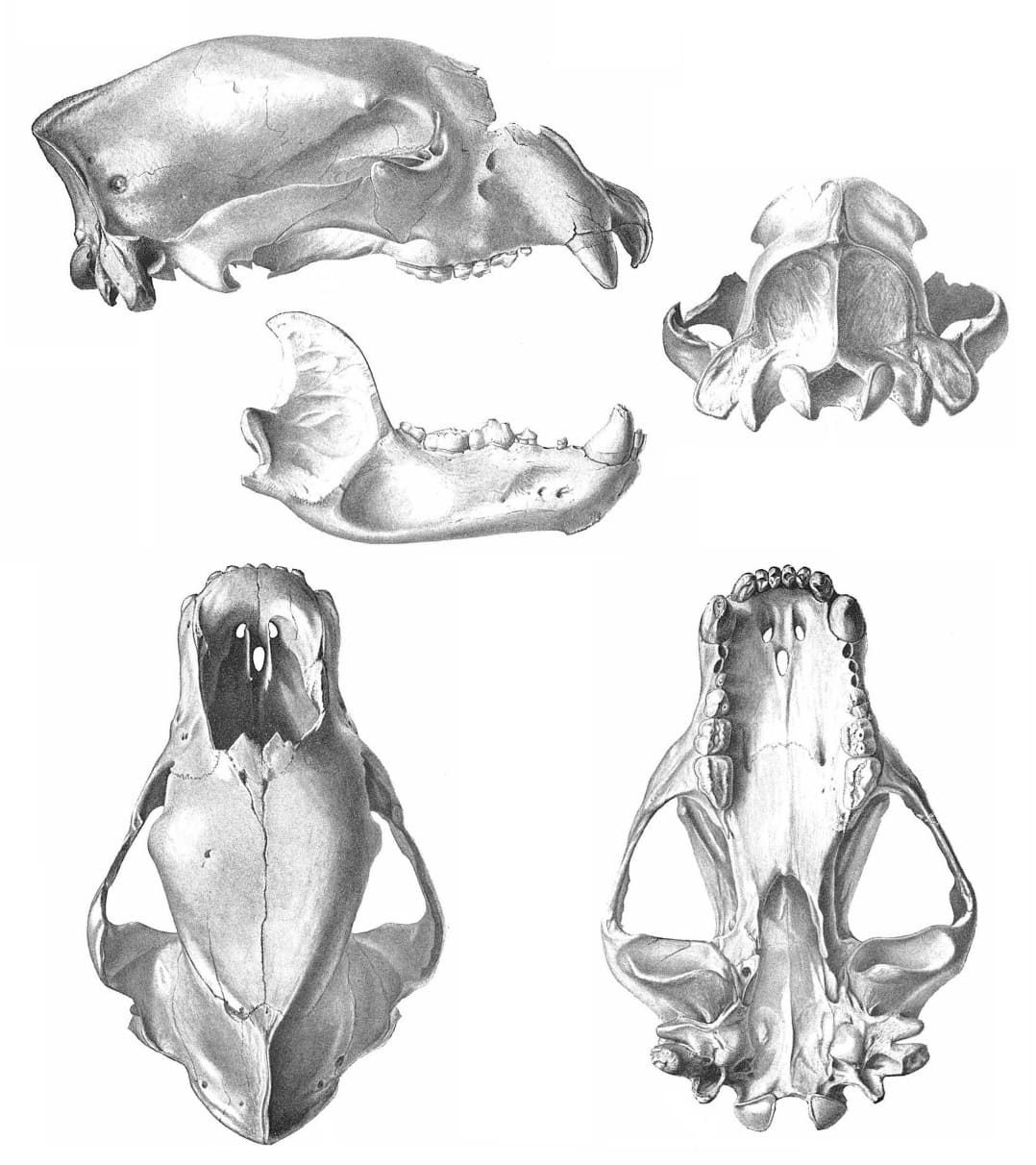
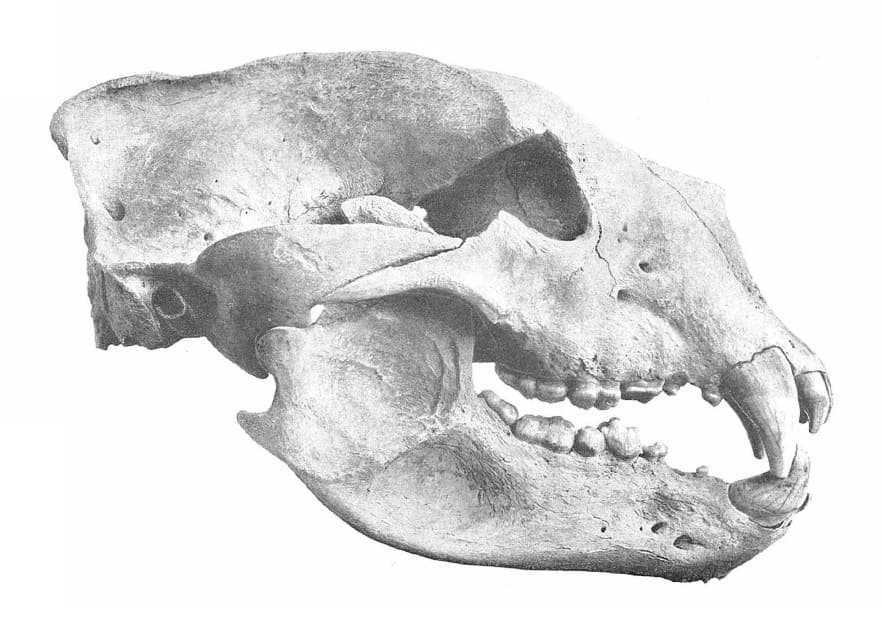
Прорисовка черепа с нижней челюстью гигантского короткомордого медведя (Arctodus simus, = Arctotherium simum) из пещеры Поттер-Крик в штате Калифорния, США.
Основная длина черепа составляет 33-44,2 см. По общим пропорциям он похож на череп
Tremarctos, а также крупных кошачьих. Лицевой отдел короткий и широкий. Носовая апертура большая, носовые кости короткие. Обонятельная зона по площади значительно превосходит таковую соразмерного
Ursus. Передний край глазницы лежит над передней частью М2. Глазницы сравнительно небольшие, широко расставленные, направлены латеральнее, чем в роде
Ursus. Такое строение глазниц характерно и для других Tremarctinae.
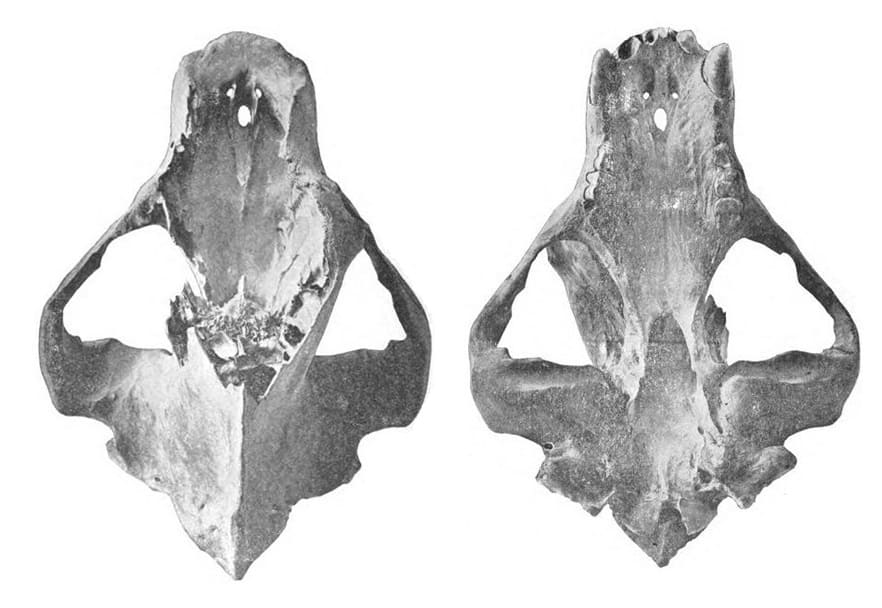
Череп с нижней челюстью гигантского короткомордого медведя (
Arctodus simus, =
Arctotherium simum) с ранчо Ла-Бреа в штате Калифорния, США.
Подглазничное отверстие двойное, расположено над серединой коронки M1. Слезное отверстие одиночное. Заглазничные отростки тупые, заглазничное сжатие резко выраженное. Скуловые дуги высокие, широко разнесенные. Лоб пологий, очень широкий. Сагиттальный гребень высокий у самцов, развит умеренно или отсутствует у самок. Костное небо широкое, в том числе в области хоан, что указывает на сильное дыхание и выносливость зверя. Слуховой барабан уплощен; нет бугра на основной затылочной кости, проходящего вдоль ее контакта с барабаном, как это наблюдается у некоторых видов Ursus. Околозатылочный и сосцевидный отростки спускаются почти до одного уровня.
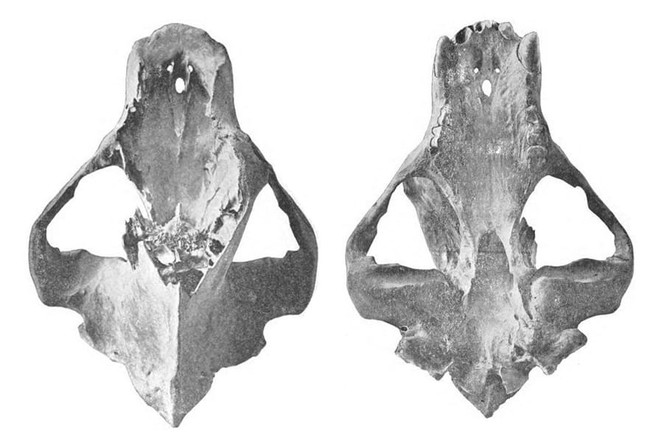
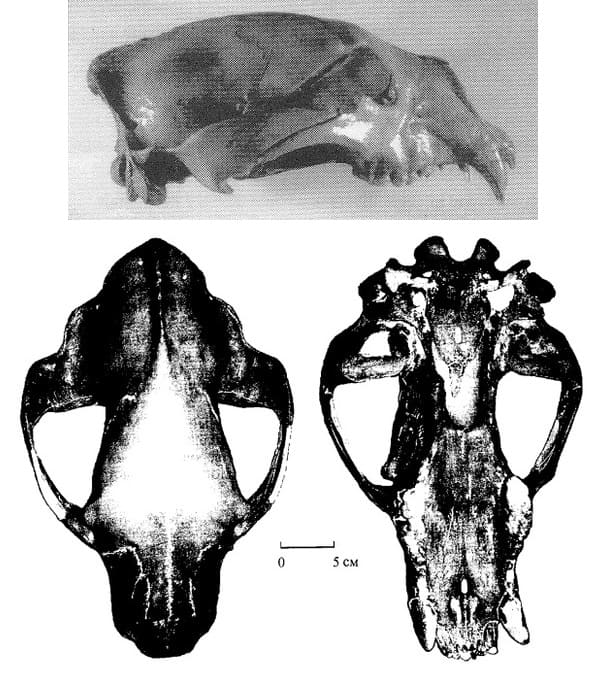
Крупнейший найденный череп самца гигантского короткомордого медведя (Arctodus simus, = Arctotherium yukonense) из Голд-Ран-Крик на территории Юкон, Канада. Полная длина данного образца составляет 52,1 см, ширина в скуловых дугах — 36,4 см. Таким образом, соотношение ширины черепа к его длине достигает почти 80 %, тогда как у бурого (Ursus arctos), черного (Ursus americanus) и пещерного (Ursus spelaeus) медведей оно никогда не превышает 70 %. Возраст черепа составляет около 26 тыс. лет.
Нижнечелюстная кость массивная, высота ее тела постепенно уменьшается от m3 к симфизному отделу. Присутствует хорошо выраженный "подбородок". Нижний край челюсти выпуклый, сочленовный отросток расположен немного выше уровня жевательной поверхности щечных зубов. Венечный отросток высокий, круто поднимающийся спереди; нижнечелюстная вырезка слабая. Имеются два больших подбородочных отверстия, лежащие впереди уровня р4, и два маленьких задних; число и расположение последних варьируют. Гребень, отделяющий премассетерную ямку от массетерной, высокий. Широкие скуловые дуги, хорошо развитые сагиттальный гребень на черепе и разделительный гребень на нижней челюсти указывают на наличие чрезвычайно мощной височной и жевательной мускулатуры. Укорочение челюстей увеличивало силу их сжатия.
Череп самки гигантского короткомордого медведя (Arctodus simus) из Офир-Крик на территории Юкон, Канада. Возраст образца составляет порядка 20 тыс. лет.
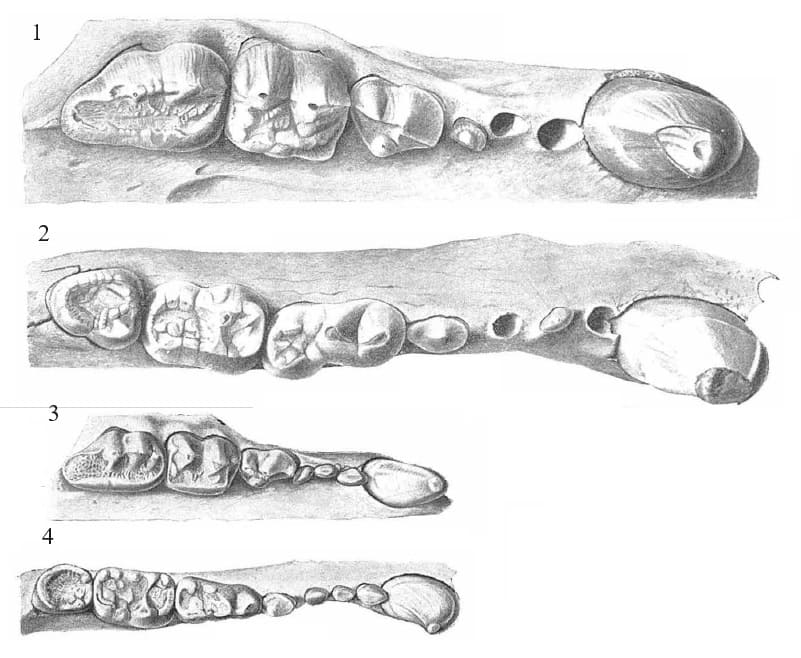
Клыки большие, высота коронки верхнего клыка около 50 мм. Передние предкоренные Р2-3/р2-3 маленькие, однокорневые. Коронка Р4 треугольных очертаний, протокон лежит на уровне хищнической щели, метакон в виде вытянутого спереди назад бугорка. Коренные зубы массивные, имеют вид широких давящих платформ. Коронка M1 квадратная, длина ее почти равна ширине. Паракон и метакон сходных размеров. Присутствуют большой парастиль и маленький метастиль. В лингвальном ряду бугорков различим четкий мезокон. Срединная долинка широкая, поверхность ее слабоскладчатая. М2 (длина 33,9-41,7 мм для 7 особей) с небольшим талоном; зубная коронка расширена в области паракона и резко сужается назад. Основания лабиальных и лингвальных бугорков разобщены продольной внутренней долинкой.
Прорисовки жевательной поверхности правых верхних и нижних зубов гигантского короткомордого медведя (Arctodus simus) из позднего плейстоцена штата Калифорния, США (1, 2), и современного очкового медведя (Tremarctos ornatus) (3, 4).
р4 уменьшен, простого строения, с двумя корнями. Нижний хищнический зуб m1 (длина 29,6-35,3 мм для 32 особей) больше, чем m2. Параконид, протоконид и метаконид выстроены почти на одной линии. Протоконид очень высокий, метаконид маленький, лежит у задней стенки протоконида. Талонид относительно короткий, но широкий, с неглубоким бассейном. m2 и m3 более простого строения, чем в роде
Ursus. Коронка m2 с длинным талонидом, который почти не уступает по ширине тригониду. Бассейн тригонида неглубокий. Бугорки талонида слабо дифференцированы. Коронка m3 небольшая, жевательная поверхность треугольной формы и слабо наклонена впереди.
Реконструированный скелет гигантского короткомордого медведя (
Arctodus simus).
Туловище сравнительно укорочено. Конечности высокие и относительно стройные. Передняя и задняя их пары примерно одинаковой длины, но благодаря сильно развитому плечевому поясу холка находится значительно выше крестца. Весь характер телосложения позволяет предположить, что гигантский короткомордый медведь был способен неутомимо преодолевать значительные расстояния. В отличии от рода Ursus, плечевая кость с внутренним надмыщелковым отверстием. Как у Ailuropoda и Arctotherium, медиальный надмыщелок плеча расширен, что указывает на определенную свободу движений в локтевом суставе.
Реконструированный скелет гигантского короткомордого медведя (Arctodus simus).
Необходимо отметить, что конечности гигантского короткомордого медведя не являлись пропорционально более длинными, чем у таких ныне живущих медведей, как белый и бурый. Ноги этих зверей также достаточно длинные, что не кажется очевидным вследствие их массивности и сильно развитого мехового покрова. Длинноногость гигантского короткомордого медведя в значительной степени является иллюзией, созданной его относительно более короткой и покатой спиной. Судя по всему, такое изменение пропорций было вызвано необходимостью уравновесить длину тела этого животного с нго значительной массой.
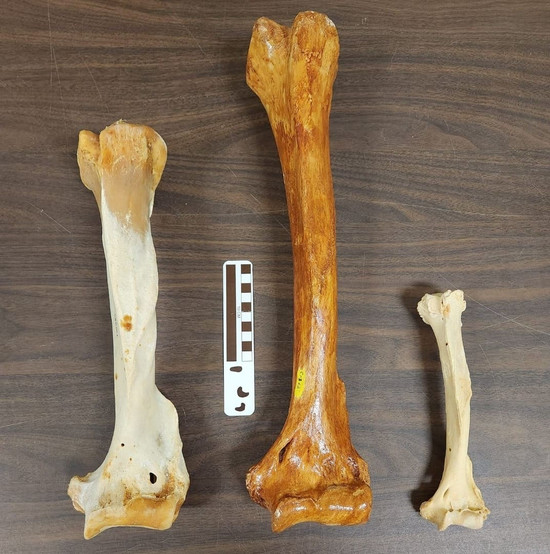
Плечевые кости, слева направо: бурый медведь (
Ursus arctos), гигантский короткомордый медведь (
Arctodus simus), очковый медведь (
Tremarctos ornatus).
По сравнению с представителями Ursinae, метаподии и фаланги кисти и стопы гигантского короткомордого медведя более длинные и тонкие, особенно в III-х и IV-х лучах. Таким образом, его лапы более симметричны, чем у урсин, оси ступней которых совмещены с наружным (V-м) пальцем. Кроме того, I-й палец расположен ближе и параллельно остальным четырем пальцам, т. е. пальцы кисти и стопы имеют меньшее латеральное разведение и в большей степени направлены прямо вперед. Все это предполагает менее косолапую походку, а значит возможность более быстрого передвижения.

Эскизы к реконструкции внешнего облика гигантского короткомордого медведя (Arctodus simus). Автор — М. Антон.
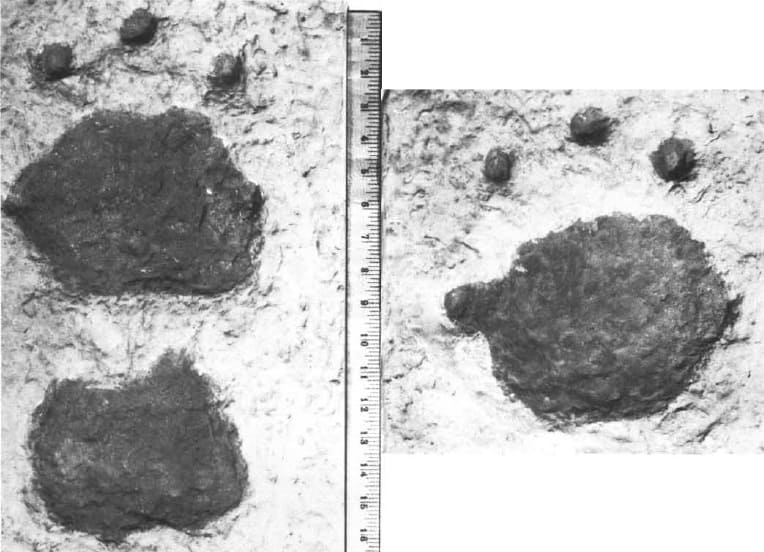
Последнему утверждению потенциально противоречит серия больших пятипалых следов, сохранившихся в уплотнившихся селевых отложениях предположительно плейстоценового возраста неподалеку от Лейквью в штате Орегон. Данные следовые отпечатки, соответствующие размерам лап гигантского короткомордого медведя, демонстрируют сильную степень разведения: II-й, III-й и IV-й пальцы равномерно выровнены по центру и направлены вперед, а I-й и V-й пальцы почти перпендикулярно (80 °) отклонены от оси стопы в каждую сторону. Также эти следы демонстрируют, что на подошвах имелись неразделенные подушечки овальной формы (отпечатки пальцевых подушечек не сохранились), пальцы несли длинные когти, передние лапы были несколько больше задних, а задняя ступня при ходьбе выходила за переднюю, как у современных медведей. Для сравнения: все пять пальцев кисти очкового медведя (Tremarctos ornatus) расположены неглубокой дугой, а их когти довольно длинные и простираются далеко вперед. Цапапины от когтей в пещере Риверблафф, также приписываемые гигантскому короткомордому медведю, свидетельствуют о лапах шириной почти 20 см.
Следовые отпечатки, предположительно принадлежащие гигантскому короткомордому медведю (Arctodus simus), из окрестностей Лейквью в штате Орегон, США.
От малого короткомордого медведя (
Arctodus pristinus) гигантский отличается более крупными размерами, его морда относительно короче, а конечности длинее. Нижняя челюсть с более глубокой горизонтальной ветвью. Щечные зубы крупнее, с теснее посаженными передними предкоренными. Однако в целом морфологических различий относительно немного, в результате чего отличить гигантского короткомордого медведя от малого бывает сложно, поскольку крупные особи второго могут перекрываться по размеру с мелкими особями первого. Гигантские короткомордые медведи, хотя обычно и превосходили по величине бурых медведей (
Ursus arctos), не являлись настолько массивными. Их конечности, особенно задние, относительно более длинные и тонкие, а туловище бочкообразно. Еще в большей степени гигантский короткомордый медведь уступает в массивности арктотерию (
Arctotherium angustidens).
Следы когтей, предположительно принадлежащие гигантскому короткомордому медведю (
Arctodus simus).
Генетика
Исследование митохондриальной ДНК, секвенированной из позднеплейстоценовых (средний возраст около 44 тыс. лет) образцов гигантского короткомордого медведя с Аляски, Юкона, Альберты и Огайо, позволяет предположить чрезвычайно низкий уровень их генетического разнообразия: для 23 изученных экземпляров установлено лишь 7 гаплотипов, к тому же образующих монофилетическую кладу. Это сопоставимо с генетическим разнообразием современных видов, находящихся под угрозой исчезновения и может объясняться прохождением через "бутылочное горлышко". Помимо гигантского короткомордого медведя, утрата генетического разнообразия перед наступлением последнего ледникового максимума отмечена для целого ряда представителей евразийской и американской мегафауны, таких как львы, бизоны, верблюды, лошади и мамонты. Это обусловлено снижением численности их популяций с последующим их восстановлением, либо вымиранием в начале голоцена.
Кроме того, снижение генетического разнообразия характерно для видов, ведущих преимущественно одиночный образ жизни, занимающих обширный ареал и имеющих относительно небольшую по численности популяцию. В настоящее время подобное отсутствие генетического разнообразия на значительных пространствах наблюдается в Африке у полосатой (Hyaena hyaena) и бурой (Parahyaena brunnea) гиен. Небольшие размеры популяции также могут быть характерны для тремарктиновых медведей: очковый медведь (Tremarctos ornatus), хотя и имеет низкий уровень генетического разнообразия, не демонстрирует признаков недавнего прохождения через "бутылочное горлышко".
Бурый медведь (Ursus arctos), наряду со многими недавно иммигрировавшими в Америку таксонами, обладали разнообразными, симпатрическими исходными популяциями в Евразии, что позволяло им осуществлять новые вселения на Американский континент. Однако если бы гигантский короткомордый медведь столкнулся с прохождением через "бутылочное горлышко" или локальным вымиранием до наступления последнего ледникового максимума, он не смог бы пополнить свое снизившиеся генетическое разнообразие за счет новых мигрантов, как это мог сделать бурый медведь, что сделало бы его уязвимым для вымирания.
Тем не менее вышеприведенные факты не исключает полностью генетическое разнообразие гигантского короткомордого медведя: его образцы из пещеры Чикиуите в Мексика, имеющие возраст порядка 14 тыс. лет, указывают на глубокое генетическое расхождение с ранее изученными образцами.
Происхождение
Как полагают, гигантский короткомордый медведь произошел от малого короткомордого медведя (Arctodus pristinus) в конце раннего плейстоцена (около 1,1 млн. лет назад, ранний ирвингтоний) и сосуществовал с ним по меньшей мере до конца среднего плейстоцена (примерно 0,3 млн. лет назад, поздний ирвингтоний). Предковый вид имел более мелкие размеры, потреблял больше растительной пищи и был привязан к лесным местообитаниям в субтропическом и тропическом поясах преимущественно на востоке и юге Северной Америки. Об экологических взаимоотношениях обоих видов ничего не известно.
Древнейшие известные остатки гигантского короткомордого медведя происходят из местонахождения Ирвингтон в Калифорнии. Появлению, а также широкому распространению в Северной Америке гигантского короткомордого медведя могло способствовать исчезновение в раннем плейстоцене псовых из подсемейства Borophaginae, до того занимавших на открытых пространствах континента ниши доминирующих хищников.
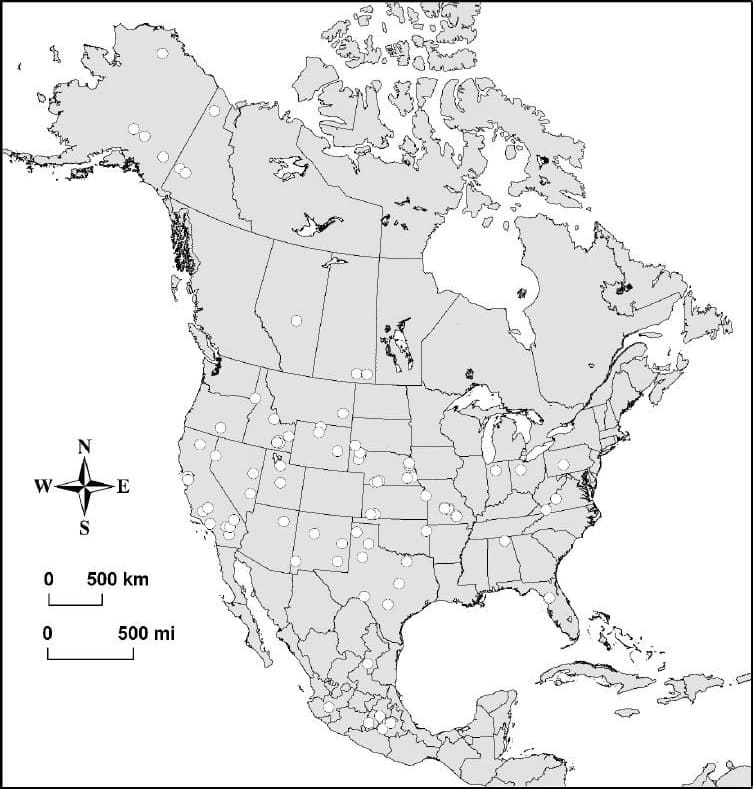
В ирвингтонии (конец раннего - конец среднего плейстоцена) гигантский короткомордый медведь обитал в центральных и западных штатах США (Техас, Канзас, Небраска, Монтана и, особенно, Калифорния), а в ранчолабреании (начало - конец позднего плейстоцена) его ареал простиралился от арктического склона хребта Брукса на севере Аляски (69 ° с. ш.) через Юкон, Британскую Колумбию, Альберту, Саскачеван и большую часть территории США от Флориды и Пенсильвании на востоке до Калифорнии и Орегона на западе (не известны на северо-востоке, редки на юго-востоке) до центральной части Мексики. Большая часть находок ископаемых остатков сосредоточена на западе США.
Основные местонахождения ископаемых остатков гигантского короткомордого медведя (Arctodus simus).
Таким образом, гигантский короткомордый медведь являлся наиболее распространенным представителем подсемейства Tremarctinae в Северной Америке. Однако, несмотря на длительный период существования и обширный географический ареал, его ископаемые остатки встречаются сравнительно редко: по состоянию на 2010 г. они были известны приблизительно из 110 местонахождений, подавляющее большинство из которых имеет позднеплейстоценовый возраст. В целом, пик разнообразия медведей на Североамериканском континенте пришелся именно на поздний плейстоцен: помимо гигантского короткомордого, здесь в это время встечались белый (Ursus maritimus), бурый (Ursus americanus), черный (Ursus americanus), флоридский пещерный (Tremarctos floridanus), арктотерий (Arctotherium wingei) и, возможно, малый короткомордый (Arctodus pristinus) медведи.