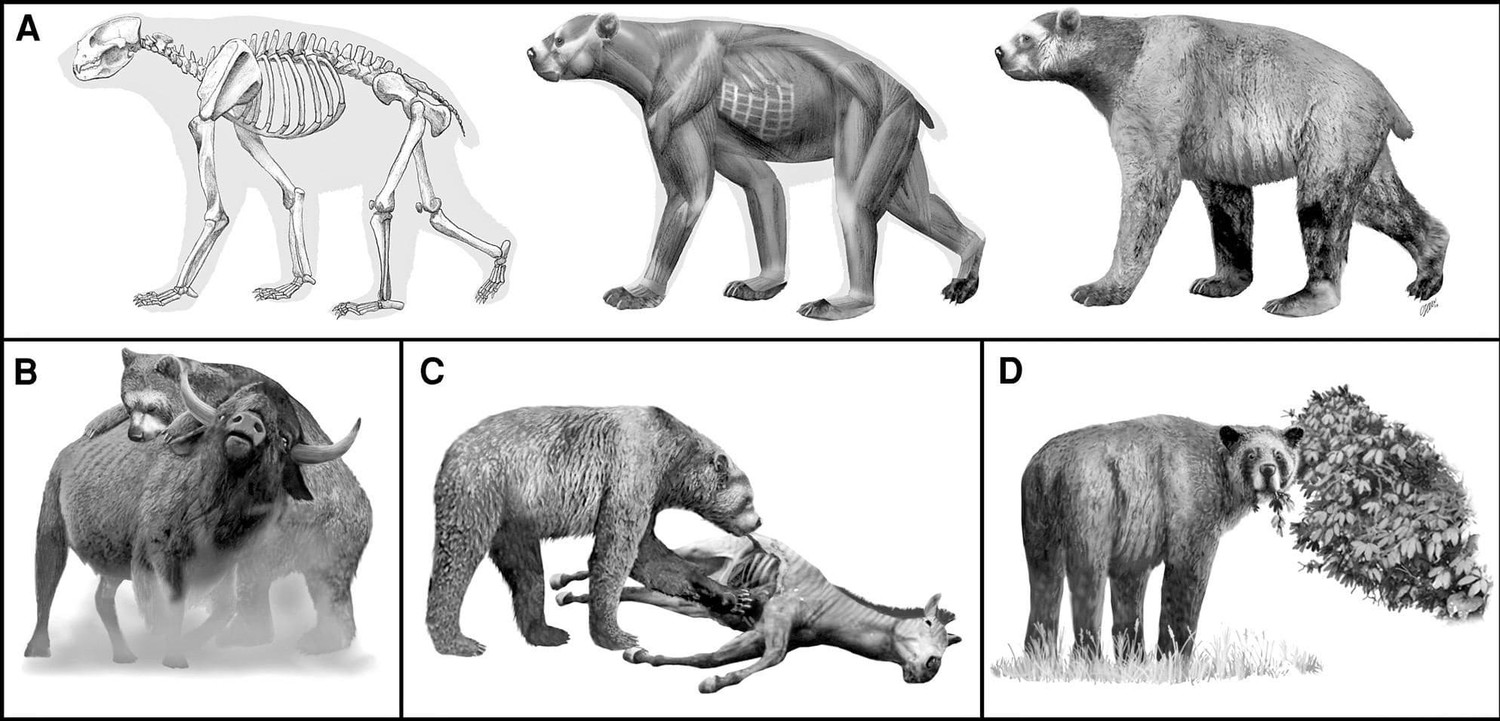
Образ жизни
Благодаря широкому распространению на североамериканском материке, гигантский короткомордый медведь населял большинство его климатических зон и сред обитания (кроме пространств, занятых Кордильерским и Лаврентийским ледниковыми щитами): тундростепи и бореальные леса Берингии, прерии и высокогорные лесаов внутреннего запада США, субтропические лесные массивы юго-востока США, смешанные саванны юго-запада США и Мексиканского нагорья, сосново-дубовые леса Транс-Мексиканского вулканического пояса на южной границе Неарктической области.
Судя по палеогеографическим условиям распространения находок ископаемых остатков, данный вид предпочитал хорошо дренированные травянистые пространства, реже встречался в разреженных лесах, посещал долины рек, берега низинных озер и болота. Судя по всему, это был хорошо приспособляемый вид, использующий множество сред обитания и возможностей для добывания пищи. Учитывая относительную редкость находок остатков гигантского короткомордого медведя по сравнению с другими крупными плейстоценовыми хищниками, предполагается, что плотность его популяции была низкой.
Имеется несколько точек зрения на образ жизни и питание гигантского короткомордого медведя. Наиболее распространенная была высказана Б. Куртеном (1967 г.). Согласно его представлениям, это был могучий, быстро бегающий хищник, способный охотиться на стадных копытных размером с бизона. Об этом свидетельствуют кошкоподобные признаки в очертаниях черепа (в частности, укороченный лицевой отдел, большая ширина в клыках, расположение скуловых дуг и др.), крупные клыки, высокие щечные зубы (особенно m2), возможный хищнический сдвиг Р4 и длинные конечности с относительно тонкими пястными и плюсневыми костями. Сходных взглядов придерживались и другие иссдедователи (Вурхиз и Корнер, 1986 г.; Шоу и Кокс, 1994 г.), полагавшие, что гигантский короткомордый медведь был более хищным, чем современные медведи, за исключением белого (Ursus maritimus).
Мнение Куртена было подвержено обстоятельной критике С. Эмсли и Н. Чаплевски (1985 г.). Они полагали, что кошкоподобные пропорции черепа гигантского короткомордого медведя объясняются редукцией предкоренных зубов и не демонстируют явной хищнической специализации, а близки к таковым ближайшего ныне живущего родственника — очкового медведя (Tremarctos ornatus) из Андского региона Южной Америки. Последний питается в основном мягкими частями растений — плодами, ягодами, листьями, корой деревьев, мякотью кактусов. При случае также поедает падаль и ловит мелких животных, лишь изредка охотясь на крупную добычу. В целом, очковый медведь имеет преимущественно растительный рацион с потенциалом оппортунистического хищничества, но конкретный его состав зависит от региона и сезонной доступности кормов. Вместе с тем нельзя не отметить, что флора поросших лесом склонов Анд гораздо богаче флоры открытых берингийских ландшафтов.
Реконструкция внешнего облика гигантского короткомордого медведя (Arctodus simus). Автор — К. Бьюэлл.
Поскольку у растительноядных хищных млекопитающих нет эффективного пищеварительного тракта для расщепления растительной массы посредством микробного воздействия, им приходится достигать этого путем интенсивного измельчения в ротовой полости и, таким образом, обладать свойствами, обеспечивающими высокое механическое преимущество жевательного аппарата. Глубокая нижняя челюсть и большая премассетерная ямка для прикрепления мышц у гигантского короткомордого и очкового медведей свидетельствуют об усилении давящей функции нижнечелюстной кости. Возможно, указанные черты представляют собой предковое для тремарктин состояние.
В отличие от специализированных плотоядных, клыки гигантского короткомордого медведя не сжаты с боков. Щечные зубы, действительно обладающие высокими коронками, несут уплощенные бугорки с небольшими фасетками срезания, что наблюдается у особей разных возрастных групп. Подобное строение жевательных поверхностей сходно с таковым у современных всеядных медведей (в особенности, очкового медведя), использующих свои зубы в качестве раздавливающих платформ, в отличие от хищников, у которых вместо этого имеются режущие лезвия. (Хотя у единственного современного сверхплотоядного урсида, белого медведя, на щечных зубах тоже отсутствуют режущие лезвия, питание этого вида главным образом жиром, а не более грубой плотью, может свести на нет необходимость в развитии у него зубной системы, специализирующейся на переработке мяса.) Горизонтальные фасетки износа бугорков щечных зубов, стертые у старых особей до почти плоских площадок, также свидетельствуют о том, что зубы гигантского короткомордого медведя использовались преимущественно для перетирания твердой и волокнистой растительности, а не для разрезания мяса, хотя вполне могли дробить и кости.
Наконец, очень крупные размеры скорее свойственны всеядным или растительноядным наземным хищным, чем специализированным мясоедам. Самый крупный из существующих медведей, кадьяк (Ursus arctos middendorfi), имеет среднюю массу около 300 кг, хотя отдельные особи могут достигать и 700 кг. Справедливости ради следует отметить, что большая панда (Ailuropoda melanoleuca), почти полный вегетарианец, весит всего до 182 кг. Соответственно, растительный рацион не ведет с неизбежностью к увеличению размеров тела. Но для сверххищника гигантский короткомордый медведь, согласно этой точке зрения, был все же слишком велик.
Кроме того, авторы гипотезы гигантского короткомордого медведя-вегетарианца усмотрели противоречие в выводах относительно гигантского короткомордого медведя-сверххищника с одной стороны, и его возможном исчезновении в результате конкуренции с бурым медведем, — с другой. Действительно, если принять интерпретацию гигантского короткомордого медведя как сверххищника, то затруднительно объяснить его проигрыш в конкурентной борьбе с более мелким и менее сильным бурым медведем.

Пластическая реконструкция внешнего облика гигантского короткомордого медведя (
Arctodus simus).
Согласно несколько модифицированной гипотезе образа жизни и пищевой стратегии гигантского короткомордого медведя, он, как и большинство современных медведей, был всеядным. Кормясь главным образом растительностью, зверь периодически также выступал в роли хищника или падальщика, предпочитая при этом крупную добычу. Основаниями для такого утверждения являются особенности строения черепа, конечностей и размеры тела. Предполагается, что высокий рост, боковая постановка небольших глаз и мощное телосложение не способствуют активной охоте. Чисто растительным рационом невозможно объяснить обитание гигантского короткомордого медведя далеко на севере, в Берингии. Кроме того, его щечные зубы не имеют специальных адаптаций для перетирания растительного корма, хотя потребность в нем у такого большого животного должна быть велика. Наконец, по величине головного мозга относительно размеров тела гигантский короткомордый медведь занимает промежуточное положение между предположительно сверхплотоядным Cephalogale и почти полностью растительноядным европейским пещерным медведем (Ursus spelaeus), что также свидетельствует в пользу его всеядности.
Г.Ф. Барышниковым с коллегами (1994 г.) была предложена палеоэкологическая реконструкция гигантского короткомордого медведя в качестве всеядного зверя, который специализировался как падальщик ("сверхгиена"), обходивший в поисках трупов крупных млекопитающих огромные территории. П. Матеус (1995, 2003 гг.) обосновал ее детальным морфофункциональным и палеоэкологическим анализом. Анализ стабильных изотопов коллагена из костей гигантского короткомордого медведя, собранных на Аляске и Юконе, также показал, что он потреблял больше мясной пищи, чем современный бурый медведь (Ursus arctos). Плейстоценовые ландшафты Северной Америки изобиловали крупными млекопитающими: мамонтами, мастодонтами, наземными ленивцами, верблюдами, бизонами, овцебыками, оленями, лошадями и другими стадными копытными, гибель которых обеспечивала существование падальщиков разной величины и различных пищевых предпочтений. (Важно отметить, что изотопные данные не позволяют отличить сверхплотоядных животных от всеядных, также потребляющих значительное количество животных веществ.)
Высоко расположенные глаза позволяли гигантскому короткомордому медведю осматривать большие площади безлесных равнин, отыскивая падаль не только с помощью острого обоняния, но и ориентируясь на поведение птиц-падальщиков. С другой стороны, близкое к латеральному положение небольших глазниц позволяет предположить, что стереоскопическое зрение не являлось для гигантского короткомордого медведя приоритетным и подтверждает малую степень его специализации к активному преследованию жертв.
Признанию гигантского короткомордого медведя в качестве "сверхгиены" отвечают его широкие скуловые дуги и короткие массивные челюсти с высокими щечными зубами, способными разрывать шкуру и отгрызать куски мяса, а также дробить большие кости для получения питательного костного мозга. Небольшая длина морды способствовала увеличению силы височных и жевательных мышц, закрывающих челюсти, а значительная ширина челюстей между клыками могла способствовать более надежному захвату добычи. Также широкая морда могла быть связана с сильно развитым обонянием, на что также указывают крупные обонятельные доли головного мозга. Схожая ситуация наблюдается и у белого медведя (Ursus maritimus), самого плотоядного ныне живущего представителя семейства. Кроме того, широкая морда могла содержать широкий горловой проход для захвата крупных кусков мяса, как у пятнистой гиены (Crocuta crocuta); впрочем, последнее характерно и для преимущественно растительноядных очкового (Tremarctos ornatus) и малайского (Helarctos malayanus) медведей.

Портретная реконструкция гигантского короткомордого медведя (Arctodus simus). Автор — leviatan_666.
Примечательно, что частота встречаемости повреждений зубов у ужасных волков, смилодонов и американских львов с ранчо Ла-Бреа в Калифорнии примерно втрое превышает этот показатель у современных крупных хищников. Это указывает на то, что конкуренция между хищниками в позднем плейстоцене была более интенсивной и позволяет предположить, что плотоядные активнее питались падалью и более полно утилизировали туши. Специализированные падальщики, такие как гиены, демонстрируют характерные закономерности повреждения щечных зубов в результате разгрызания костей. На основании отсутствия соответствующих следов микроизноса на зубах гигантского короткомордого медведя из Ла-Бреа и ряда других местонахождений делается вывод, что этот зверь не являлся специализированным падальщиком. Наоборот, у южноамериканского арктотерия (Arctotherium angustidens) весьма обычны тяжелые повреждения зубных коронок и альвеолярные инфекции. Это интерпретируется как свидетельство питания твердыми материалами (например, костями), что может указывать на регулярное поедание этим медведем туш копытных.
Однако, хотя в Ла-Бреа были найдены остатки большего числа особей гигантского короткомордого медведя, чем в любом другом месте (предположительно, в связи с благоприятными условиями захоронения), они составляют лишь 1 % от числа всех хищников в битумных ямах, что аналогично данному показателю у бурого и черного медведей — всеядных с уклоном в растительноядность. На этом основании высказано предположение, что в Ла-Бреа гигантский короткомордый медведь не соперничал активно за туши с другими хищниками. В пещере Нэтурал-Трап в Вайоминге, представлявшей собой естественную ловушку для хищников и растительноядных, были найдены только 2 экземпляра гигантского короткомордого медведя. Это составляет примерно 0,9 % от общей численности представителей мегафауны в данном местонахождении, что соответствует относительной численности гигантского короткомордого медведя в Ла-Бреа. Выявленные зубные патологии, например стертые резцы и наддесневой зубной камень у молодого животного, а также кариес, вызванный потреблением углеводов у других особей из Ла-Бреа, также предполагает наличие достаточно большого количества растительности в их рационе.
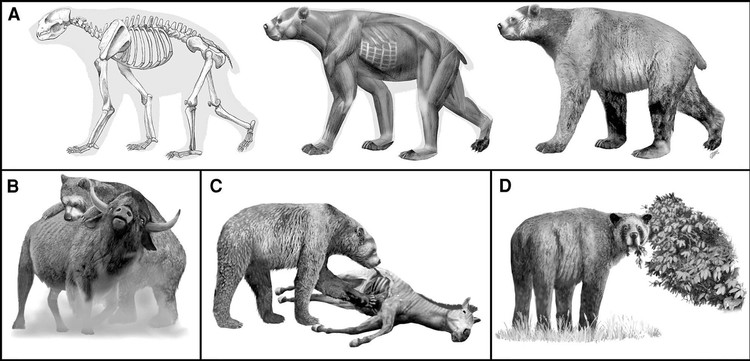
Этапы реконструкции внешнего облика гигантского короткомордого медведя (Arctodus simus) и различные типы его питания.
В ряде случаев остатки гигантского короткомордого медведя обнаружены в ассоциации с остатками хоботных — например, костями молодого американского мастодонта (Mammut americanum) неподалеку от Франкстауна в Пенсильвании, костями мамонта Колумба (Mammuthus columbi) в пещере возле водохранилища Хантингтон в Юте и в массовом захоронении мамонтов Колумба в Хот-Спрингс, Южная Дакота. Обычно остается неясным, указывают ли эти находки на хищнические или падальнические отношения, либо же остатки оказались в одном местонахождении случайно. С другой стороны, иногда на элементах скелета хоботных сохраняются следы зубов крупных хищников, которые вполне могут принадлежать гигантскому короткомордому медведю. Таковы, в частности, пяточная кость шерстистого мамонта (Mammuthus primigenius) из Солтвилля в Вирджинии, плечевая кость американского мастодонта из Сноумастодон в Колорадо, а также несколько костей мамонтов Колумба из вышеупомянутых местонахождений Хантингтон в Юте и Хот-Спрингс в Южной Дакоте.
Важно отметить, что резцы и клыки гигантского короткомордого медведя соразмерны с таковыми американского (Panthera atrox) и пещерного (Panthera spelaea) львов, что усложняет идентификацию следов зубов. Однако это не дискредитирует все зубные следы, приписываемые гигантскому короткомордому медведю. Так, находка поврежденных длинных костей в пещере Рампарт на Аляске позволяют предположить, что медведь притаскивал в находящееся здесь логово части тел млекопитающих и поедал их, в то время как львы имели в Берингии ограниченное распространение. Кроме того, считается, что в пасти гигантского короткомордого медведя побывала прокушенная подвздошная кость пекари из пещеры Шериден в Огайо. Следы зубов на черепном фрагменте (и, возможно, плечевой кости) гигантского короткомордого медведя из пещеры на острове Ванкувер приписываются другой его особи на том основании, что этот зверь является здесь единственным установленным крупным наземным хищником.

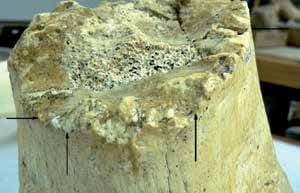
Слева: остатки гигантского короткомордого медведя (Arctodus simus) из Хот-Спрингс в штате Южная Дакота, США. Во времена позднего плейстоцена (около 25 тыс. лет назад) здесь находилась карстовая воронка с водоемом, представлявшая собой природную ловушку для мамонтов Колумба (Mammuthus columbi). Они приходили сюда на водопой и для отдыха, а когда когда уровень воды внезапно поднимался, не успевали спастись и погибали. Воронка с разлагающимися трупами мамонтов должна была привлекать гигантских короткомордых медведей. Часть скелета, в том числе череп и нижняя челюсть одного из них сохранились в более мелкой части воронки. По его слабо стертым зубам, размеру черепа и нижней челюсти можно заключить, что это молодая особь, не старше 6 лет. В переводе на возраст современных медведей — это особь, живущая самостоятельно в течение 2-3 лет. В этой же части водоема найдены несколько ребер и часть плечевой кости еще одной, более молодой особи. Основываясь на особенностях поведения современных медведей и немногих данных о возможных особенностях экологии и этологии гигантского короткомордого медведя можно предполагать, что между этими плотоядными происходили схватки за право первым подобраться к трупу мамонта, и для некоторых из них такие столкновения заканчивались гибелью. Кости молодых медведей захоронены в мелководной части водоема, с пологим склоном, откуда можно было легко выйти на берег — это подтверждает, что они, скорее всего, не утонули в воронке-ловушке подобно мамонтам, а были убиты или смертельно ранены своими сородичами. В ассоциации с костными остатками гигантского короткомордого медведя были найдены и его копролиты. Примечательно, что помимо костных фрагментов они содержат семена можжевельника.
Справа: фрагмент дистальной части большой берцовой кости мамонта Колумба (Mammuthus columbi) из Хот-Спрингс. На поперечном сломе кости видны следы скусывания, оставленные крупным хищником. По размеру выемок от передних зубов можно определить, что его размерный класс соответствует гигантскому короткомордому медведю (Arctodus simus) или американскому льву (Panthera atrox). Кости последнего в воронке-ловушке пока не обнаружены, кроме того, современные львы редко едят кости слонов и носорогов. Разгрызание диафизов костей характерно именно для медведя. У мамонтов и слонов нет костномозговой полости внутри длинных костей конечностей. Эти кости полностью состоят из губчатой ткани, ячейки которой заполнены костным мозгом, на 80 % состоящим из жира. Вот почему хищник поедал внутреннюю часть костей, разгрызая толстые плотные наружные стенки.
Необходимо учитывать, что туши крупных млекопитающих с большой продолжительностью жизни и низкой плодовитостью не могли составлять основу рациона гигантского короткомордого медведя ввиду их относительной редкости и большого числа других хищников, при случае также охотно поедавших падаль. Таким образом, охотиться ему приходилось бы главным образом на крупную живую добычу, а питание падалью было скорее спорадическим. Однако вследствие своих крупных размеров гигантский короткомордый медведь был не в состоянии долго преследовать быструю добычу вроде копытных. Поэтому кажется вполне вероятным, что он специализировался как клептопаразит, отнимающий животных, убитых другими хищниками — смилодоном (Smilodon fatalis), американским львом (Panthera atrox), ужасным волком (Aenocyon dirus), серым волком (Canis lupus), более мелкими медведями. По всей видимости, естественный отбор способствовал увеличения размеров тела гигантского короткомордого медведя, чтобы он мог добывать и защищать добытые им туши от других крупных хищников, часть из которых была стайной, либо отбирать их собственную добычу.
Радиохимический анализ костей гигантского короткомордого медведя с Аляски показал высокий уровень содержания в них азота-15 — стабильного изотопа азота, накапливаемого плотоядными. Хотя известно небольшое количество образцов, в настоящее время нет свидететельств наличия у них зубных патологий, связанных с потреблением значительного объема углеводов, которые характерны для южных популяций гигантского короткомордого медведя. На основании этих данных было высказано предположение, что гигантский короткомордый медведь был более плотоядным в Берингии, чем в остальной части Северной Америки. При этом предпочтение отдавалось фитофагам, потребляющим С3-растительность и вследствие этого накапливающим углерод-13 — в частности, северному оленю (Rangifer tarandus) и овцебыку (Ovibos moschatus). Вместе с тем, результаты изотопного анализ берингийских образцов гигантского короткомордого медведя указывают на низкий уровень потребления мяса лошадей и мамонтов, несмотря на то, что они составляли около половины доступной биомассы в регионе.

Гигантский короткомордый медведь (Arctodus simus) кормится трупом шерстистого мамонта (Mammuthus primigenius). Автор — Б. Зайкен.
Увеличение роли хищничества может быть связано с меньшим числом конкурентов и, вероятно, с меньшей доступностью богатых углеводами продуктов питания в условиях Крайнего Севера. Согласно расчетам, в Берингии гигантский короткомордый медведь весом 700 кг потреблял около 5,9 т мяса в год, что эквивалентно 12 бизонам, 45 кабаллоидным лошадям или 2 шерстистым мамонтам (с поправкой на несъедобные части тела). Следовательно, гигантскому короткомордому медведю пришлось бы добывать 100 кг плоти или съедобной падали каждые 6,25 дней (16 кг в день). Тем не менее, его локальное вымирание в Берингии около 23 тыс. лет назад (значительно раньше, чем в других частях ареала) поднимает вопрос о том, почему гигантский короткомордый медведь не проник в регион повторно, когда свободный ото льда коридор вновь открылся на юге и Берингия снова была заселена пещерным львом и бурым медведем.
Помимо этого, судя по значительному количеству изотопа углерода-13 в образцах из Аляски, Калифорнии и Мексики, гигантский короткомордый медведь потреблял много C3-растительности, когда она была доступна — листьев, стеблей, цветов, ягод, плодов, коры деревьев, кустарников и трав. Таким образом, это было оппортунистическое всеядное животное с достаточно гибким, адаптированным к местным условиям рационом, напоминающим таковой бурого медведя.
Относительно стиля локомоции гигантского короткомордого медведя мнения также расходятся. Согласно одному из них, этот зверь передвигался с умеренной скоростью походкой более специализированной, чем современные медведи. Большие размеры тела, высоко расположенный плечевой пояс, короткая и покатая спина и длинные конечности повышали эффективность передвижения, поскольку эти черты ведут к возрастанию количества полезной энергии упругого напряжения в сухожилиях и увеличивают длину шага. Все это указывает на выносливость гигантского короткомордого медведя и его бо́льшую приспособленность к длительному, чем к высокоскоростному передвижению. Основанные на гиеноподобных пропорциях расчеты показали, что животное от спокойного передвижения шагом со скоростью 8,5 км/ч могло переходить к бегу скоростью 18,5 км/ч, будучи способным развить максимальную скорость 40-45 км/ч. Если судить по другим млекопитающим, оптимальная для гигантского короткомордого медведя скорость передвижения составляла 13,7 км/ч (для сравнения: современные гиены преодолевают местность со скоростью около 10 км/ч).

Гигантский короткомордый медведь (Arctodus simus) преследует молодого бизона. Автор — Р. Евсеев.
В посткраниальном скелете гигантского короткомордого медведя отсутствуют высокоподвижные суставы, которые позволили бы ему совершать быстрые повороты — способность, необходимая любому хищнику, преследующему маневренную добычу. Кроме того, особенности строения поясничных позвонков ограничивало возможность ускорения, как и у бурого медведя. Массивные позвонки имеют прямоугольную форму и лишены рычагов, позволяющих межпозвоночным мышцам сгибать позвоночный столб. Вероятно, его ограниченная способность к сгибанию и разгибанию в сагиттальной плоскости вела к снижению максимальной скорости бега.
Значения индексов, определяющих отношение длины лучевой кости к плечевой, у гигантского короткомордого медведя равняется 78,2-87,6. У бурого медведя среднее значение соответствующего индекса равно 88,4 (вариационный размах 83-95), а у черного медведя — 90,3 (вариационный размах 85,5-97,9). Значение индексов, определяющих отношение длины большой берцовой кости к бедренной, у гигантского короткомордого медведя составляет 71,4-74,8. У бурого медведя среднее значение этого индекса – 73,5 (вариационный размах 70,4-75,1), а у черного медведя — 77 (вариационный размах 74,8-78,4). Очевидно, что пропорции задних конечностей у гигантского короткомордого медведя практически не отличаются от таковых у бурого медведя. Что же касается передних конечностей, то 2 показателя из 3 находятся в границах вариационного размаха бурого медведя. Учитывая абсолютно большую длину конечностей гигантского короткомордого медведя, его меньшую косолапость и пропорции конечностей, почти совпадающие с пропорциями бурого медведя (во всяком случае, относительно длинных костей), то предположение о несколько более выраженных курсорных тенденциях гигантского короткомордого медведя в сравнении с бурым кажется вполне обоснованным.
Однако необходимо учитывать, что адаптацией к быстрому бегу является увеличение длины дистальных отделов конечностей, а не проксимальных, как это наблюдается у гигантского короткомордого медведя. Удлинение проксимальных отделов конечностей (т. е. увеличение длины плечевой и бедренной костей) — характерная черта зверей, которым часто приходится рыть землю. Это увеличение как раз и свойственно многим медведям. Гигантский короткомордый медведь, однако, утратил другие характеристики животных, которым часто приходится рыть землю, — в частности, длинный локтевой отросток и большие когти. К тому же, как и другие медведи, он продолжал оставаться стопоходящим животным, что также не указывает на хорошую приспособленность к быстрому бегу с безопорной фазой. Соответственно, длинные стройные конечности не позволяли зверю делать скоростные рывки, необходимые для того, чтобы настигать добычу.

Гигантский короткомордый медведь (Arctodus simus) отгоняет от туши американского лося (Alces americanus) пару американских львов (Panthera atrox). Автор — Р. Евсеев.
Современный гривистый волк (Chrysocyon brachyurus) имеет экстремально длинные и тонкие конечности, бегая при этом не особенно быстро и довольно неуклюже. Предполагается, что длинные конечности нужны были гигантскому короткомордому медведю, как и гривистому волку, для увеличения обзора, особенно на высокотравных равнинах. Помимо этого, длинные конечности позволяли доставать и пригибать ветви небольших деревьев и кустарников при питании листьями, плодами или корой. Вместе с тем, пронация предплечья, сгибание запястья и пальцев, а также мускулатура относительно тонких передних конечностей, которые имеют решающее значение для захвата крупной добычи передними лапами, судя по всему, были развиты в меньшей степени, чем у бурого медведя или крупных кошек из рода Panthera.
Хотя известно более сотни местонахождений остатков гигантского короткомордого медведя, лишь на одном из них был обнаружен его бакулюм (кость полового члена). Судя по всему, это отображает как тафономические, так и поведенческие особенности данного животного. Практически все скелетные остатки, принадлежащие по-настоящему крупным особям (предположительно, самцам), происходят из открытых местонахождений, где обычно было найдено лишь несколько элементов. Напротив, пещерные отложения дали многочисленные примеры находок относительно небольших, но более полно сохранившихся особей, вместе с которыми, вероятно, были бы найдены и бакулюмы, если бы они имелись. В общей сложности, в пещерах было обнаружено порядка 70 % из числа наиболее мелких образцов. Например, кости всех 8 особей из пещеры Поттер-Крик в Калифорнии судя по всему принадлежат самкам. Небольшой размер обнаруженных элементов скелета и отсутствие бакулюмов позволяют предположить, что самки гигантского короткомордого медведя устраивали в пещерных ходах логова для выведения потомства (примечательно, что подобная тенденция отмечена и для Arctotherium angustidens из Южной Америки). Впрочем, судя по имеющимся данным, в некоторых пещерах были найдены остатки животных обоих полов. В общей сложности, из пещерных местонахождений происходит около 1/3 всех обнаруженных остатков гигантского короткомордого медведя.
Следы медвежьих когтей в пещере Риверблафф, штат Миссури, наиболее многочисленны в камерах и ведущих туда проходах, что указывает на их тесную связь с логовами. Другие найденные здесь отпечатки включают следы когтей крупной кошки (Panthera atrox или Smilodon fatalis) и следы плоскоголового пекари Platygonus. В пещере Биг-Биар, также на территории Миссури, в ассоциации с остатками гигантского короткомордого медведя обнаружены фоссилизированные волосы.
Самка и детеныш гигантского короткомордого медведя (Arctodus simus) в берингийском ландшафте. Автор — Х. Ласерда.
Исследование остатков молодой особи из пещеры Озарк в центральной части США позволяет предположить, что гигантский короткомордый медведь, как и другие урсиды, достигал половой зрелости раньше остеологической. Сравнение с известными последовательностями слияния эпифизов у ныне живущего черного медведя (Ursus americanus) показало, что, хотя на момент смерти зверь не являлся остеологически зрелым, поскольку его многочисленные эпифизы не срослись. Стадия сращения эпифизарных пластин длинных костей указывает на то, что животное практически достигло взрослых размеров, таким образом сексуально созрев задолго до того, как слияние эпифизов завершится. Если бы гигантский короткомордый медведь по срокам созревания напоминал черного медведя, тогда указанный экземпляр был бы уже половозрелым с возрастом 4-6 лет для самки, либо 6-8 лет для самца. Кроме того, характер износа зубов данной особи аналогичен 4-6-летнему черному медведю. Для сравнения: самки очкового медведя (Tremarctos ornatus) достигают половой зрелости примерно в 4 года, самки черного медведя (Ursus americanus) становятся половозрелыми в возрасте 2-4 лет, а самки бурого медведя (Ursus arctos) начинают размножаться в некоторых частях своего ареала примерно в 3 года. Для определения взрослости также используются стадии сращения межкостных швов и прорезывания зубов.
Анализ костей гигантских короткомордых медведей показывает, что иногда они страдали от таких заболеваний, как остеомиелит, туберкулезоподобные и, возможно, сифилисподобные инфекции. Так, наиболее полно сохранившийся скелет из Индианы демонстрирует целый ряд патологических нарушений: поражения позвонков и ребер, деформирующие разрастания дельтовидного и грудного гребней на правой плечевой кости, абсцессы на обеих локтевых костях, а также между m1 и m2 обеих зубных костей нижней челюсти. Перечень предполагаемых диагнозов данной особи включает продолжительный сифилис, туберкулез, остеомиелит, артрит и грибковое инфицирование раны отдельно или в сочетании с другими причинами.
Эпифиз плечевой кости американского мастодонта (Mammut americanum) из местонахождения Сноумастодон в штате Колорадо, США, несущий погрызы крупного хищника, предположительно гигантского короткомордого медведя (Arctodus simus).