Возраст и распространение
Возможно, горгоноповые — самая обособленная группа диноморф. Их вероятные близкие родственники происходят из средней перми Приуралья — это фтинозухиды, ранее включавшиеся в состав сборной группы "эотериодонтов". Ископаемые остатки собственно горгоноповых обнаружены в средне-позднепермских отложениях Южной и Восточной Африки (большинство находок), Восточной Европы (преимущественно север Европейской части России). Неописанные остатки известны также из Южной Америки, Нигера и Китая. Отпечатки следов найдены в Италии.
В Южной Африке первые настоящие горгоноповые известны из биостратиграфической зоны Tapinocephalus. В Восточной Европе горгоноповые исчезли незадолго до конца пермского периода — в терминальных пермских слоях (Вязниковский фаунистический комплекс) они не обнаружены. В Южной Африке они дожили до пермо-триасовой границы, но из терминальных слоев известны лишь рубиджеины.
Образ жизни
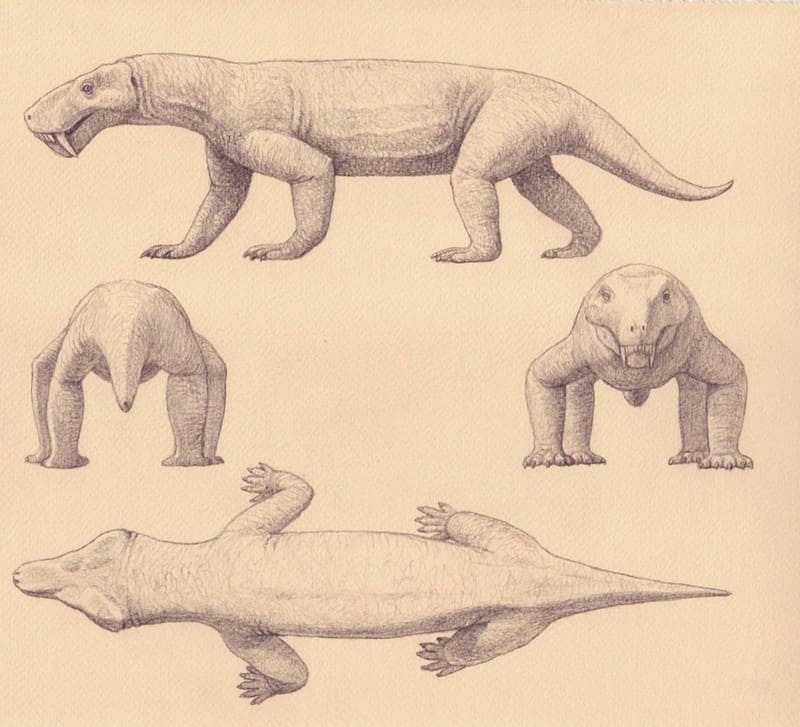
Реконструкция внешнего облика рубиджеи (Rubidgea atrox). Автор — М. Антон.
Горгоноповые — исключительно плотоядные формы, часто весьма крупные. Обычно они были доминирующими хищниками в своих биотопах, сменив в этой роли хищных диноцефалов. Некоторыми деталями строения и адаптациями они напоминают саблезубых кошек. Так, у обеих групп наблюдается увеличение верхних клыков и развитие для них своеобразных "полуножен" на нижней челюсти, сокращение числа и размера заклыковых зубов и целый ряд анатомических особенностей, позволяющих широко раскрывать пасть для укуса. Большие когти на лапах тоже помогали при охоте. Вероятно, среди горгоноповых наблюдалось значительное экологическое разнообразие — часть их могла являться полуводными хищниками, возможно, даже охотниками за водной добычей. О последнем, в частности, может свидетельствовать предполагаемое наличие электрорецепторов, которые среди современных млекопитающих имеются у утконоса. Судя по строению глазных косточек, горгоноповые охотились по ночам и в сумерках. Пищей им служили прежде всего дицинодонты и парейазавры.

Иностранцевии (Inostrancevia sp.) атакуют скутозавра (Scutosaurus sp.). Автор — М. Антон.
Вымирание
Высказано предположение, что в конце перми синапсидное сообщество могло развалиться из-за горгоноповых. Они были тесно связаны с доминирующими фитофагами дицинодонтами и составляли с ними пары хищник - жертва. Резкие перепады температуры в конце пермского периода, в том числе серьезное похолодание, привели горгоноповых к гибели: их примитивная терморегуляция не справилась с температурными катаклизмами. В результате освободилась большая и перспективная ниша охотников за дицинодонтами, которые сами, скорее всего, были теплокровными и спокойно перенесли похолодание. Нишу крупных хищников попробовали занять сразу несколько групп животных, окончательно разваливших наземное природное сообщество пермского периода.

Рубиджеи (Rubidgea atrox) и элурогнаты (Aelurognathus tigriceps), разрывающие дицинодонта. Автор — М. Антон.
Систематика
Впервые горгоноповых описал Р. Оуэн. Примечательно, что из-за неполноты первого найденного черепа их долгое время считали анапсидами.
Классификация горгоноповых основывается на различиях пропорций черепа: степени длинномордости; форме сечения через морду, которая может быть прямоугольной или квадратной; наличии предклыковой "ступеньки" в крае верхней челюсти; длине теменного отдела; широком или узком выступании наружу скуловых дуг. Самая дробная классификация была предложена Д. Уотсоном и А. Ромером (1956 г.), разделившими "типичных" горгонопов на 15 самостоятельных семейств.
На основании широкой ревизии южноафриканских горгоноповых Д. Сигоньо (1970 г.), опираясь на сходство пропорций черепа, свела в синонимы почти половину ранее выделенных родов. Типичных южноафриканских горгонопов Д. Сигоньо объединила в единственное подсемейство горгонопин — Gorgonopinae. В другое подсемейство — Rubidgeinae — Д. Сигоньо, помимо собственно рубиджеин (с типовым родом Rubidgea), включает также род Sycosaurus (по Д. Уотсону и А. Ромеру (1956 г.) — семейство Sycosauridae). Единственное семейство Gorgonopidae для всех прогрессивных горгонопов принимает и Т. Кемп (1982 г.).
В общей сводке по горгонопам Д. Сигоньо-Расселл (1989 г.) также принимает всех прогрессивных горгонопов в объеме семейства Gorgonopidae, разделенного на 3 подсемейства: Gorgonopinae, Rubidgeinae и Inostranceviinae. К высшим горгонопам автор также относит североамериканское семейство Watongiidae.
Компромиссную упрощенную схему предложил Л.П. Татаринов (1974 г.), сохранив на основе африканских форм всего 7 подсемейств: Gorgonopinae, Broomisaurinae, Aelurosaurinae, Sycosaurinae и Cynariopinae, а также новые Cyonosaurinae и Arctopinae, выделенные на основе комбинации африканских родов. В данном варианте сохранилось, конечно, и семейство северодвинских горгоноповых Inostranceviidae. Эту схему принял А. Бринк (1986, 1988 гг.).
М.Ф. Ивахненко (2000, 2002, 2003, 2008 гг.) упорно работал над совершенствованием классификации горгонопов. Он разделил высших горгонопов, принимаемых им в ранге инфраотряда Gorgonopida, на 2 надсемейства — Rubidgeoidea и Gorgonopioidea.
Более примитивных рубиджеоидей М.Ф. Ивахненко (2000, 2001, 2008 гг.) характеризует отсутствием или небольшой величиной межтеменной кости, расширенной заглазничной дугой, опущенной под глазницу скуловой костью, резко расширенной в сквамозной области, так что височные окна открываются косо вверх, и субпараллельностью наружных краев теменного щита. Помимо типового семейства Rubidgeidae, М.Ф. Ивахненко (2008 г.) относит к рубиджеоидеям и Inostranceviidae, а также Phthinosuchidae, но в весьма суженном по сравнению со схемой Л.П. Татаринова объеме.
По М.Ф. Ивахненко (2000, 2001, 2008 гг.), собственно фтинозухиды отличаются от других рубиджеоидей умеренно удлиненным черепом, очень крупными глазницами, слабо расширенной заглазничной дугой, отсутствием гребня на ангулярном лепестке и большим числом (не менее 7) заклыковых зубов.
К семейству Inostranceviidae М.Ф. Ивахненко (2008 г.) относит только род Inostrancevia — общеизвестного представителя Северодвинской фауны. Другой род северодвинских горгонопсов — Pravoslavlevia — М.Ф. Ивахненко вывел из числа иностранцевиид. Иностранцевии характеризуются крупными размерами, относительно узким черепом, удлиненными височными окнами с оттянутыми назад верхнезадними углами, отсутствием межтеменной кости, расхождением кзади латеральных углов теменного щита, средней величины глазницами, широкой скуловой дугой и умеренно расширенной заглазничной дугой. Интересны отсутствие небных бугров, редукция небных зубов, гипертрофия удлиненных клыков и полная утрата заклыковых зубов на нижней челюсти. Весьма вероятно родство иностранцевий с крупными рубеджеидами.
Собственно рубиджеид М.Ф. Ивахненко (2001 г.) характеризует укороченным черепом, небольшими глазницами, сильно расширенной за-глазничной дугой, узостью небно-птеригоидных бугров с малым числом мелких зубов и уменьшением числа заклыковых зубов максимум до 6. Апоморфии рубеджеид в строении сквамозного края височной ямы, крайне расширенного у этих горгоноповых, делают их, несмотря на слабое развитие предтеменной кости, более специализироваными, чем хотя бы гиппозавриды, одно время включаемые М.Ф. Ивахненко в число горгонопиоидей. В состав рубиджеид этот автор включал до 3 подсемейств: Sycosaurinae, Rubidgeinae и Broomocephalinae. Их представители известны преимущественно из поздней перми Южной Африки.
Горгонопиоидеи, согласно М.Ф. Ивахненко (2000, 2001, 2008 гг.), отличаются крупными размерами межтеменной кости, не расширенностью заглазничной дуги, узостью скуловой дуги, полого изогнутой вниз, практически латеральной плоскостью височного окна. По М.Ф. Ивахненко (2008 г.), группа включает 3 семейства: Galesuchidae, Cyonosauridae и Gorgonopidae. Более примитивные галезухиды обладают сравнительно низким и широким черепом, крупными глазницами и сравнительно мало расширенными кзади височными окнами. К ним принадлежат около 5 родов только из зоны Tapinocephalus Южной Африки.
В составе собственно горгоноповых М.Ф. Ивахненко (2001 г.) первоначально принимал 4 подсемейства — Aelurosaurinae, Cyonosaurinae, Gorgonopinae и Scylacopinae. В своей более поздней ревизии (2008 г.) М.Ф. Ивахненко оставил лишь 2 подсемейства — Gorgonopinae и Scylacopinae. Они известны из Южной и, реже, Восточной Африки, но единичные формы описаны также из верхней перми Восточной Европы.
На сегодняшний день классификация горгонопов остается недостаточно разработанной. Положение в системе семейств Phthinosuchidae, Burnetiidae, Ictidorhinidae и Hipposauridae (Сигоньо-Рассел (1989 г.)), а также Watongiidae (Ивахненко (2001 г.)), определено лишь в самых общих чертах. Сближение горгоноповых с бурнетиевыми, иктидориновыми и гиппозавровыми, не обладающими выраженным венечным отростком, приводит, если принять систему М.Ф. Ивахненко, к выводу, что венечный отросток развился независимо у рубиджеид и иностранцевиид, что, во всяком случае, не доказано. Только у типичных горгоноповых задний край наружного ангулярного лепестка далеко отстоит от области нижнечелюстного сустава, тогда как у бурнетиид, иктидоринид, гиппозаврид, фтинозухид и ропалодонтид (=эстемменозухид), как и у диноцефалов и сфенакодонтных пеликозавров, ангулярный лепесток почти достигает челюстного сустава. Значение этих различий еще не установлено.
Однако в обсуждаемых вариантах системы горгоноповых внимание излишне акцентируется на пропорциях черепа (Сигоньо (1970 г.), Сигоньо-Рассел (1989 г.)). М.Ф. Ивахненко (2001, 2008 гг.) добавил к этому строение височной области черепа, скуловой дуги и небных бугров. Но некоторые краниологические различия, по существу, по-прежнему игнорируются. Л.П. Татаринов (2009 г.) указывает хотя бы на особенности в ушной области, базисфеноиде, нижней челюсти (засуставный отросток и др.). Он отмечает также, в частности, на то, что отнесенные М.Ф. Ивахненко к горгоноповым одного подсемейства Scylacopinae роды Scylacops, Sauroctonus и Suchogorgon столь резко различаются по строению стремени (у Scylacops оно толстое, с массивным верхним отростком, у Suchogorgon — грацильное, у Sauroctonus — лишь с миниатюрным стапедиальным отверстием, но с массивным дорсальным отростком), что невольно возникают сомнения в их близком родстве.
Suchogorgon резко отличается от Sauroctonus наличием очень длинного засуставного отростка нижней челюсти, практически не выраженного у Sauroctonus. Короткий засуставный отросток указан и для Gorgonops, у Arctops он массивный, но прямой, а у Arctognathus отчетливо длинный, загнутый на конце вперед, как у сухогоргона. У последних 2 родов его конструкция может служить модельной формой для развития среднего уха млекопитающих. Все это указывает на недостаточную убедительность существующих объемов подсемейств горгоноповых в понимании М.Ф. Ивахненко и на необходимость дальнейшей работы в построении их системы.
Количество родов горгоноповых в старых исследованиях достигало 30, ревизия группы Е. Гебауэр (2007 г.) сократила их число до 10-15. Дело в том, что многие южноафриканские формы были описаны как различные роды по остаткам молодых особей либо особей разного размера.
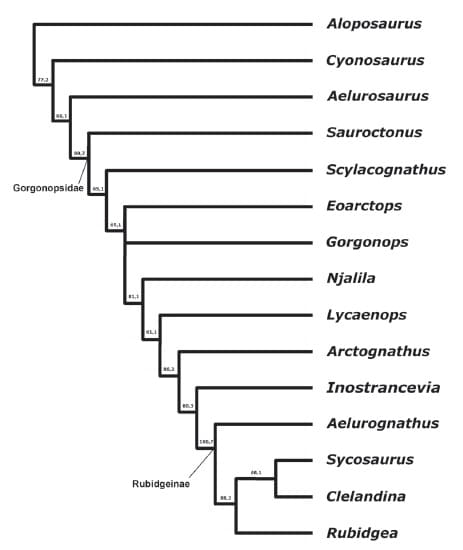
Кладограмма, иллюстрирующая родственные связи горгоноповых (по Гебауэр, 2007 г.).
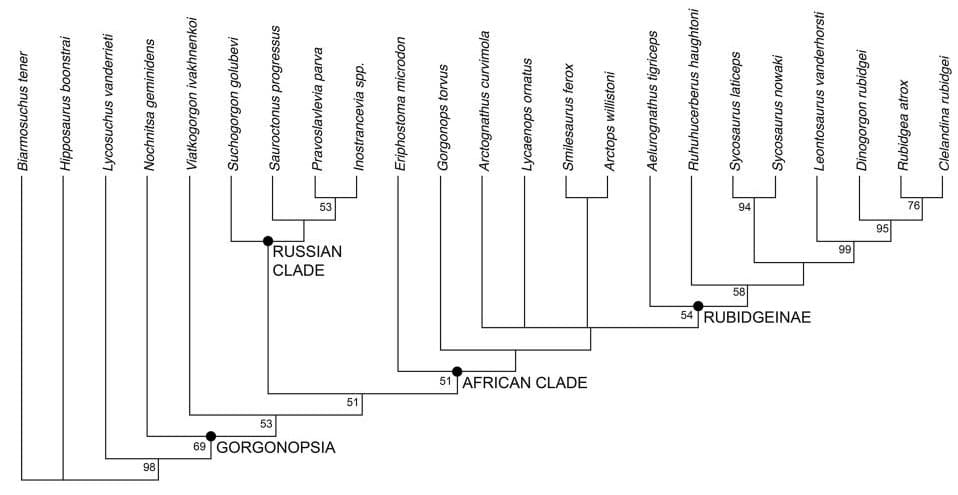
Кладограмма, иллюстрирующая родственные связи горгоноповых (по Каммереру, 2018 г.).
Согласно современным воззрениям, семейство имеет примерно следующий состав:
"Российская клада";
род †Ночницы (Nochnitsa Kammerer et Masyutin, 2018);
род †Вяткогоргоны (Viatkogorgon Tatarinov, 1999);
род †Сухогоргоны (Suchogorgon Tatarinov, 2000);
род †Завроктоны (Sauroctonus Bystrov, 1955);
подсемейство †Иностранцевиины (Inostranceviinae Pravoslavlev, 1927);
род †Православлевии (Pravoslavlevia Vjuschkov, 1953);
род †Иностранцевии (Inostrancevia Amalitzky, 1922);
"Африканская клада";
род †Форкисы (Phorcys Kammerer et Rubidge, 2022);
род †Эрифостомы (Eriphostoma Broom, 1911);
род †Горгонопсы (Gorgonops Owen, 1876);
род †Алопозавры (Aloposaurus Broom, 1910);
род †Ционозавры (Cyonosaurus Olson, 1937);
род †Элурозавры (Aelurosaurus Owen, 1881);
род †Ньялилы (Njalila Gebauer, 2007);
род †Лиценопсы (Lycaenops Broom, 1925);
род †Смилезавры (Smilesaurus Broom, 1948);
род †Арктопсы (Arctops Watson, 1914);
род †Арктогнаты (Arctognathus Broom, 1911);
подсемейство †Рубиджеины (Rubidgeinae Broom, 1938);
род †Элурогнаты (Aelurognathus Haughton, 1924);
род †Рухухуцерберы (Ruhuhucerberus Maisch, 2002);
род †Сикозавры (Sycosaurus Haughton, 1924);
триба †Рубиджеини (Rubidgeini (Broom, 1938));
род †Леонтозавры (Leontosaurus Broom et George, 1950);
род †Диногоргоны (Dinogorgon Broom, 1936);
род †Рубиджеи (Rubidgea Broom, 1948);
род †Клеландины (Clelandina Broom, 1948).
Сомнительные рода:
род †Арктозухи (Arctosuchus Broom, 1911);
род †Цердодоны (Cerdodon Broom, 1915);
род †Цердогнаты (Cerdognathus Broom, 1915);
род †Цинариоды (Cynariodes Broom, 1930);
род †Цинискодоны (Cyniscodon Broom, 1915);
род †Цинодраконы (Cynodracon Owen, 1876);
род †Дельфациогнаты (Delphaciognathus Broom, 1932);
род †Геновумы (Genovum von Huene, 1950);
род †"Цердорины" ("Cerdorhinus" Droom, 1936);
род †Ликозавры (Lycosaurus Lycosaurus Owen, 1876); род †Псевдогиппозавры (Pseudohipposaurus Boonstra, 1952);
род †Сцимногнаты (Scymnognathus Broom, 1915);
род †Леогоргоны (Leogorgon Ivachnenko, 2003).