К вопросу о моно- или полифилии млекопитающих
На основании изучения древнейших позднетриасовых и раннеюрских млекопитающих можно сделать вывод, что их предков следует искать среди неизвестных, сравнительно мелких хищных цинодонтов из конца среднего или самого начала позднего триаса. Данную точку зрения, возможно, первым высказал Дж. Хопсон (1967 г.). В целом сочувственно, но сдержанно к такой идее относится К. Гоу (1997 г.).
Особую актуальность вопрос о моно- или полифилетическом происхождении млекопитающих приобрел после работ Дж. Симпсона (1959, 1960 гг.), пришедшего к выводу об условности разграничения этих понятий на палеонтологическом уровне. До того усилия исследователей сводились к поискам ближайшей к млекопитающим группе рептилий, в качестве которой ранее обычно называли цинодонтов из группы тритилодонтов, а позднее — трителедонтов. А.П. Быстров (1955, 1957 гг.) предпочел рассматривать всех зверообразных как членов единого класса рептилий. Особенно категорично этот автор высказался о горгонопах, которых называл типичными рептилиями.
Сегодня очевидно, что Tritylodontidae, Brasilodontidae, Tritheledontidae, Dromatheriidae, Therioherpetidae и Probainognathidae не являются прямыми предками млекопитающих. Обычно их называют сестринскими группами по отношению к млекопитающим, но даже это весьма условно. Приходится допускать или вторичность утраты многими из этих цинодонтов пояска на заклыковых зубах, или вторичное его развитие у примитивнейших млекопитающих. Это обстоятельство давно уже ставит в тупик исследователей. А. Кромптон (1972 г.) пришел к выводу об ответвлении млекопитающих от цинодонтов, более примитивных, чем Thrinaxodon, близкого мнения ранее придерживался и Л.П. Татаринов (1976 г.). Дж. Хопсон (1994 г.) из-за наличия дополнительного челюстного сустава маммального типа у Probainognathus, предположил, что предки млекопитающих были близкими к этому цинодонту, но обладали зубами тринаксодонтного типа.
Настоящие предковые цинодонты сочетали прогрессивную промаммальную морфологию с относительно примитивной зубной системой. Они должны были иметь относительно грацильную нижнюю челюсть, но их угловая кость не имела определенного сходства с барабанной костью млекопитающих. Но высшие зверообразные, как тероцефалы, так и, особенно, цинодонты, во многих отношениях образуют переход от примитивных терапсид к древнейшим позднетриасовым млекопитающим. В некоторых отношениях высшие цинодонты могли даже опережать наиболее примитивных позднетриасовых и раннеюрских млекопитающих. Эти цинодонты, по сравнению с исходными млекопитающими, имеют более массивную нижнюю челюсть, коренные зубы сильнее уклоняются от прототриконодонтного типа. У примитивнейших млекопитатощих еще не было сформировано среднее ухо из 3 косточек — древнейшей формой, обладавшей таким ухом, был, по-видимому, раннеюрский Hadrocodium. Процес постепенной маммализации терапсид схематически изображен ниже.
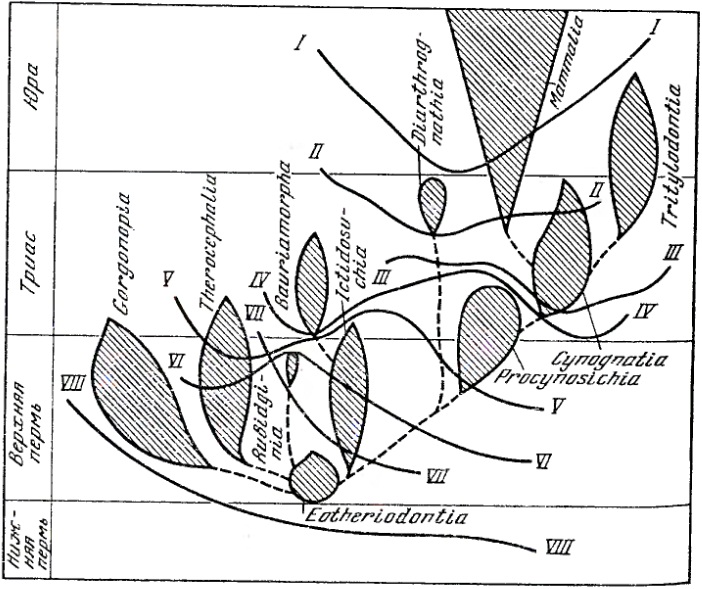
Схема путей маммализации терапсид (по Татаринову, 1987). Кривые линии соответствуют уровням, на которых приобретались: I — звукопроводящий аппарат из 3 косточек; II — вторичное челюстное сочленение между зубной и чешуйчатой костями; III — зачаточная барабанная перепонка в вырезке угловой кости; IV — мягкие, снабженные собственной мускулатурой губы; V — сенсорная зона на верхней губе (протовибриссы); VI — расширенные большие полушария головного мозга; VII — трехзубчатые заклыковые зубы; VIII — верхние обонятельные раковины.
Некоторые отличия от прогрессивных цинодонтов улавливаются и в строении задней части мозговой коробки примитивных позднетриасовых млекопитающих. В этом отношении весьма интересны особенности строения ушной капсулы одного из древнейших млекопитающих, позднетриасового Adelobasileus. Неясным остается также вопрос, насколько продвинутые цинодонты и архаичные млекопитающие продвинулись в отношении перехода от поли- к дифиодонтности. Но высшие цинодонты, высшие аномодонты и даже некоторые диноцефалы приобретали вертикальную постановку конечностей, свойственную млекопитающим
Создается определенное ощущение, что ближайшие общие предки млекопитающих еще не приобрели всего комплекса признаков, характерных для млекопитающих. Но речь не идет о настоящей полифилии, поскольку маммальные признаки почти целиком приобретались в очень узкой группе цинодонтов. В таком случае можно было бы подразумевать «парафилию», если бы этот термин не получил повсеместное употребленение в кладистике для обозначения ложнородственных групп. Наглядное представление о реальном ходе филогении можно видеть в работе С. Келян-Яворовской (1992 г.), в которой цинодонтовый ствол соединяется с млекопитающими 6-7 отдельными ветвями. Усложненная картина происхождения млекопитающих показана и в ряде других работ.
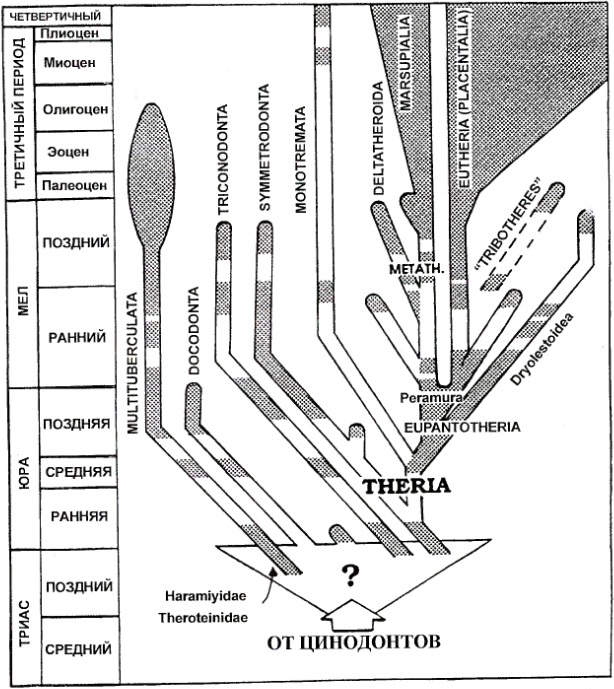
Схема филогенетических связей высших цинодонтов и млекопитающих (по Келян-Яворовской, 1992).
В последние десятилетия почти всеобщее распространение получили кладистические принципы филогении, в которых филогенетическое древо строится по последовательности отдельных признаков, приобретенных в ходе эволюции. Этим признакам отчасти произвольно приписывается ключевое значение, и по их последовательности выделяются все таксоны, число которых резко возрастает. Один из типичных примеров такого подхода на материале происхождения млекопитающих приведен в работах Т. Роу (1999 г.) и Н. Хоттона (1991 г.). В принципе, кладистический метод стремится довести предка исследуемой группы до одного корня, а в идеале — до одного вида. Увлечение кладистической филогенетикой охватило и многих молекулярных генетиков, которые стремятся вывести из одного общего генотипа даже самые далекие в генетическом отношении группы организмов.
Иной подход к систематике больших по объему таксонов демонстрирует М.Ф. Ивахненко (2008 г.), который последовательно придерживается принципа выделения «синдрома признаков», характеризующих исследуемый таксон. Принцип «синдрома признаков» М.Ф. Ивахненко распространяет также и на другие группы тетрапод. Но переход от одного синдрома к другому остается не вполне отработанным. Еще важнее то, что, по крайней мере, в конкретном случае млекопитающих преждевременно говорить о выделении комплексного синдрома признаков, характеризующих всех или почти всех примитивных представителей класса. Л.П. Татаринов (2009 г.) полагает более целесообразным в подобных случаях говорить о широких параллелизмах в развитии комплекса признаков, характеризующих самые разные высшие таксоны. То же замечание остается в силе и для многих примеров из области кладистической филогенетики.
Процесс эволюционного формирования у цинодонтов предшественников маммального уха из 3 слуховых косточек и связь этого процесса с преобразованием челюстного сочленения описаны во многих работах — например, у Хопсона (1966 г.), Бархьюзена (1968 г.), Аллина (1975, 1986 гг.), Татаринова (1987 г.). Один из возможных путей постепенного формирования у цинодонтов и предков млекопитающих среднего уха маммального типа показан на следующем рисунке. В этой трансформации большую роль мог сыграть передненижний отросток угловой кости, гомолог которого у сумчатых поддерживает снизу барабанную перепонку (вся угловая кость у сумчатых превратилось в барабанную кость). Некоторые архаичные млекопитающие, например, позднетриасовый Eozostrodon, еще сохраняли первичный сустав между сочленовной и квадратной костями.
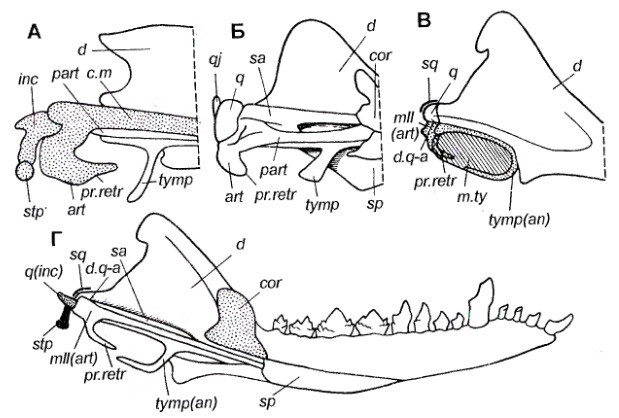
Схема эволюции среднего уха у цинодонтов и млекопитающих (по Татаринову, 2009).
Задняя часть нижней челюсти: А — эмбрион сумчатого Perameles; Б — цинодонт Diademodon; В — позднетриасовое млекопитающее Eozostrodon; Г — полная нижняя челюсть Eozostrodon.
То, что в этой перестройке мог быть не единственный путь развития, отражено на другом рисунке, где показаны различия в положении наружного слухового прохода у монотремат и остальных современных млекопитающих. У первых этот проход идет ниже остатков задних костей нижней челюсти и ниже нижнечелюстного ретрактора, а у вторых — выше ретрактора и позади остатков задних нижнечелюстных костей. В то же время, признаки явных изменений в тимпанальном направлении у задней части угловой кости отсутствуют. Таким образом, завершение формирования среднего уха у монотремат и остальных млекопитающих вполне может являться дифилетическим.
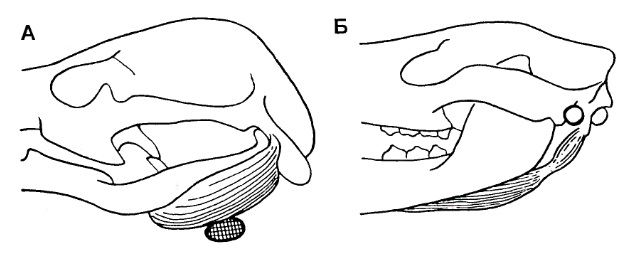
Различия в положении наружного слухового прохода у монотремат (А) и остальных современных млекопитающих на примере сумчатого Didelphis (Б) (по Татаринову, 2009).
Образование маммального челюстного сустава, разумеется, не единственный пример параллелизмов при формировании млекопитающих. Таких примеров масса. Остановимся еще на одном примере — вхождении восходящей пластины эпиптеригоида в комплекс алисфеноида, образующего предушную стенку мозговой коробки. Обычно уменьшенный в размерах эпиптеригоид пропускает выходящие сквозь него нижнечелюстную и верхнечелюстную ветви тройничного нерва, но у монотремат верхнечелюстная ветвь выходит вместе с глазничной ветвью впереди эпиптеригоида. У монотремат, а может быть, первично у всех млекопитающих в формировании алисфеноида участвует покровная кость неясного происхождения.
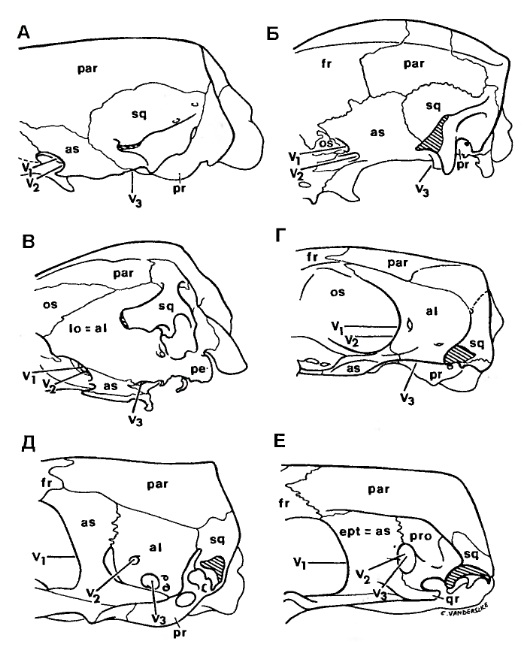
Задняя часть мозговой коробки млекопитающих и высших цинодонтов (по Татаринову, 2009):
А — Asioryctes nemegtensis; Б — Didelphis marsupialis; В — Ornithorhynchus anatinus; Г — Chulsanbaatar vulgaris; Д — Morganucodon oehleri; E — Pachygenelus monus.
У позднетриасовых морганукодонтид эпиптеригоид остается свободным от вхождения в мозговую коробку, что можно было бы связать с педогенезом, но у прогрессивного позднетриасового цинодонта Pachygenelus эпиптеригоид, возможно, срастался с алисфеноидом, но оставался вполне свободным от проотики.
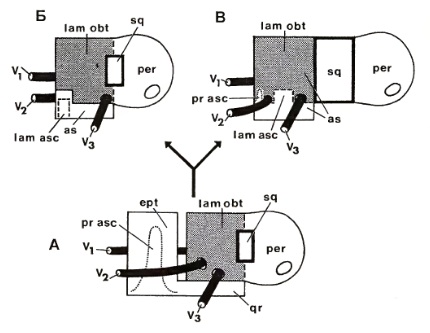
Схема гомологии эмбриональных компонентов мозговой коробки млекопитающих (по Татаринову, 2009):
А — примитивное состояние млекопитающих; Б — монотрематы; В — типичные млекопитающие.
Эволюция предушной стенки эндокрания во многом остается нерешенной проблемой. Создается впечатление, что при формировании этой стенки был как бы «вычеркнут» прогресс, достигнутый у прогрессивных цинодонтов, и ее образование у первичных млекопитающих каждый раз шло частично по особому пути. Этот пример показывает, как много белых пятен остается в проблеме происхождения млекопитающих.
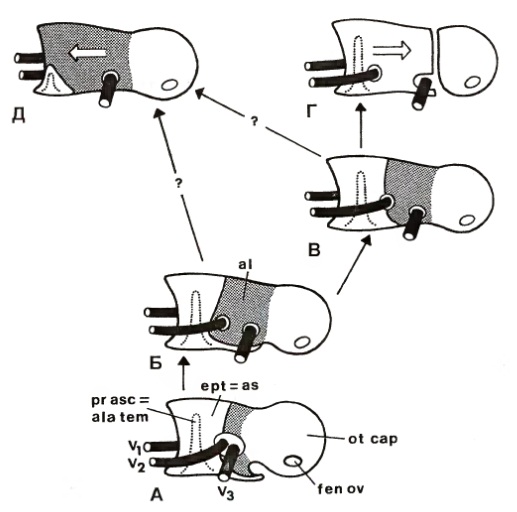
Схема эволюции прсдушной стенки эндокрания у цинодонтов и млекопитающих (по Татаринову, 2009):
А — типичный цинодонт, далекий от млекопитающих; Б — раннсюрское млекопитающее типа Morganucodon; В — раннемеловое нетрибосфеническое млекопитающее типа Vincelestes; Г — позднемеловое сумчатое дидельфидного облика; Д — монотрематы, мультитуберкуляты и, возможно, триконодонты.
В морфологическом плане грани между цинодонтами и млекопитающими, как и между многими другими таксонами самого высокого ранга, с каждым годом стираются. Где кончается одна группа и начинается другая, все чаще приходится решать произвольно. Такова общая закономерность эволюции. Уже все чаще приходится решать подобные вопросы, базируясь на широчайшем распространении параллелизмов в макроэволюции. Но в любом случае, граница между млекопитающими и цинодонтами исчезающе мала, ее просто не существует.