Цинодонты и происхождение млекопитающих (продолжение)
Продвинутые цинодонты
Эволюция черепа
Большинство основных признаков черепа млекопитающих развилось у хищных цинодонтов. Chiniquodontidae еще сохраняют заглазничные и предлобные кости, утраченные Tritheledontidae и Tritylodontidae. С исчезновением заглазничной перегородки височное окно сливается у этих групп с глазницей, как у ранних млекопитающих. У Chiniquodontidae вторичное небо простирается назад по существу так же далеко, как у примитивных млекопитающих.
У современных млекопитающих основная часть черепа — крупная, луковицевидная мозговая коробка, образованная в основном дермальными костями, а именно лобной, височной и чешуйчатой. В противоположность этому у примитивных амниот она состоит в первую очередь из хряща и замещающих его костей. Изменения, приведшие к мозговой коробке млекопитающих, начались у ранних терапсид вовсе не из-за необходимости вместить в нее крупный мозг, а для расширения зон отхождения челюстных мышц и укрепления черепа в связи с действием на него сил, возникающих при пережевывании питии. Среди тероцефалов уже прослеживается тенденция к разрастанию покровных костей крыши черепа вниз и вперед от верхнего и заднего краев аддукторной камеры, но этот процесс продвинулся гораздо дальше у цинодонтов. У Thrinaxodon чешуйчатая кость образует заднюю границу аддукторной камеры и простирается вниз до уровня слуховой капсулы. Теменная кость разрастается латерально и вентрально поверх первичной хрящевой стенки мозговой капсулы. Сзади эндоскелетные кости отико-окципитальной части мозговой коробки примитивных амниот сохраняются. У всех цинодонтов заднеушная кость заметна снаружи у основания затылочной поверхности в виде стержневидного парокципитального отростка, а заднетеменная, таблитчатая и верхнезатылочная кости разделены постоянными швами, так же как и боковые и основная затылочные. Эти кости срастаются в единую затылочную пластину у продвинутых млекопитающих.
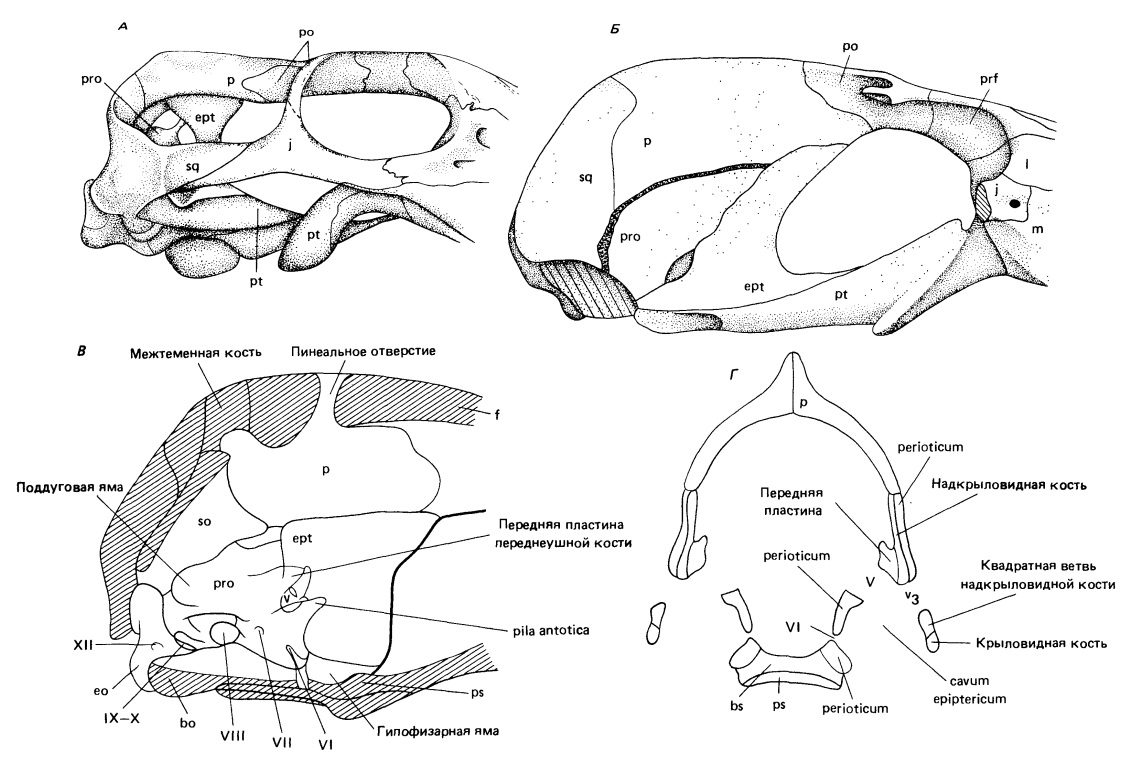
Эволюция мозговой коробки млекопитающих. А. Задняя часть черепа тероцефала Regisaurus. Как и у примитивных амниот, мозговая коробка все еще образована в основном эндохондральными костями и хрящом. Теменная и чешуйчатая кости — преимущественно поверхностные элементы. Надкрыловидная кость образует широкий вертикальный столбик латеральнее мозга. Б. Латеральный вид мозговой коробки среднетриасового цинодонта Probainognathus. Теменная и чешуйчатая кости распространились вниз, образовав дополнительную площадь прикрепления челюстной мускулатуры. Они достигают слуховой капсулы, формируя сзади сплошную стенку мозговой коробки. Надкрыловидная кость соединяется швом со слуховой капсулой и теменной костью, образуя часть стенки латеральнее первичной хрящевой мозговой коробки. Область последней медиальнее глазницы остается неокостеневшей. В. Медиальный вид латеральной стенки мозговой коробки раннетриасового цинодонта Thrinaxodon. Г. Поперечный разрез задней части мозговой коробки Thrinaxodon, показывающий расположение надкрыловидной полости между околоушной и надкрыловидной костями.
У пеликозавров, как и у примитивных амниот, переднеушная кость (prooticum) — простой элемент, образующий переднюю часть слуховой капсулы и окружающий передний край овального окна (fenestra ovalis). У цинодонтов эта кость, которая теперь называется околоушной (perioticum), разрастается вперед и образует сверху шовные соединения с вентральными пластинами теменной и чешуйчатой костей.
У ранних синапсид надкрыловидная кость (epipterygoideum) была узким стержнем, отходящим вверх от крыловидной кости на уровне базикраниального сочленения. У цинодонтов она разрастается в широкую пластину, соединенную швами дорсально с теменной, а постеродорсально — с околоушной костью, образуя костную стенку латеральнее примитивной мозговой коробки и спереди от слуховой капсулы. Включенную в мозговую коробку надкрыловидную кость можно теперь называть, как и у млекопитающих,— крылоклиновидной (alisphenoideum). От всех прочих элементов мозговой коробки она отличается происхождением из составной части висцеральной дуги.
Постеровентральная часть надкрыловидной (крылоклиновидной) кости у цинодонтов расположена латеральнее околоушной. Между ними находится пространство, называемое надкрыловидной полостью (cavum epiptericum), заключающее полулунный и коленчатый ганглии V-го черепномозгового нерва современных рептилий. Эта область открыта вентрально, т. е. еще не окружена мозговой коробкой.
У примитивных цинодонтов отверстие для нижнечелюстной ветви тройничного нерва находится между околоушной и крылоклиновидной костями. Глазная и, возможно, верхнечелюстная ветви идут вперед медиальнее крылоклиновидной кости и выходят по ее переднему краю. VII-й (лицевой) нерв проходит через околоушную кость позади отверстия для нижнечелюстной ветви V-го нерва.
Спереди околоушная кость у Thrinaxodon разделена на поверхностную пластину, простирающуюся латерально до крылоклиновидной кости, и более медиальную структуру, переднюю пластину. Относительный размер этих двух частей околоушной кости важен для диагностики раннцх представителей основных линий млекопитающих в позднем мезозое. Спереди от крылоклиновидной кости и медиальнее глазницы боковая стенка мозговой капсулы остается по происхождению хрящевой.
Слепки полостей мозговых коробок цинодонтов свидетельствуют, что их мозг не намного крупнее, чем у других рептилий со сравнимым весом тела, но отличается наличием боковых придатков мозжечка (flocculi), прилегающих к внутреннему уху и контролирующих равновесие у современных млекопитающих. Это могло быть связано с более устойчивой вертикальной ориентацией конечностей цинодонтов.
Происхождение среднего уха млекопитающих
У Chiniquodontidae и Tritylodontidae строение нижней челюсти приближается к схеме млекопитающих в связи с крупным размером зубной кости. У всех цинодонтов венечный отросток разрастается дорсально, предоставляя дополнительную площадь для прикрепления аддукторов и обеспечения лучшего рычажного действия при смыкании челюстей. У Cynognathidae разрастается угловой отросток для более эффективного прикрепления поверхностной жевательной мышцы. У Chiniquodontidae развивается также мыщелковый отросток (processus condylaris), образующий у млекопитающих челюстной сустав. Отмечено, что отбор на увеличение длины этого отростка должен был сначала способствовать сохранению постдентальных костей, поскольку селективное преимущество развития сустава между зубной и чешуйчатой костями не могло проявиться, пока эти кости не пришли в соприкосновение.
Факторы, связанные с жеванием, не подходят для объяснения сопутствующих ему изменений постдентальных костей. Если бы усиление нижней челюсти для более эффективного сопротивления возникающим при пережевывании силам было главным фактором этого отбора, следовало бы предположить, что постдентальные кости, включая сочленовную, должны были остаться прочно прикрепленными к зубной у всех терапсид. Однако уже самые ранние цинодонты (а также тероцефалы) утрачивают шовное соединение между зубной и другими нижнечелюстными костями, которые, по-видимому, были способны двигаться независимо, по крайней мере в ограниченной степени. Причины постепенного уменьшения размеров и потери связи этих костей с зубной долгое время были загадкой. Наиболее правдоподобное объяснение разработано Аллином (1975 г.) в связи с происхождением среднего уха млекопитающих.
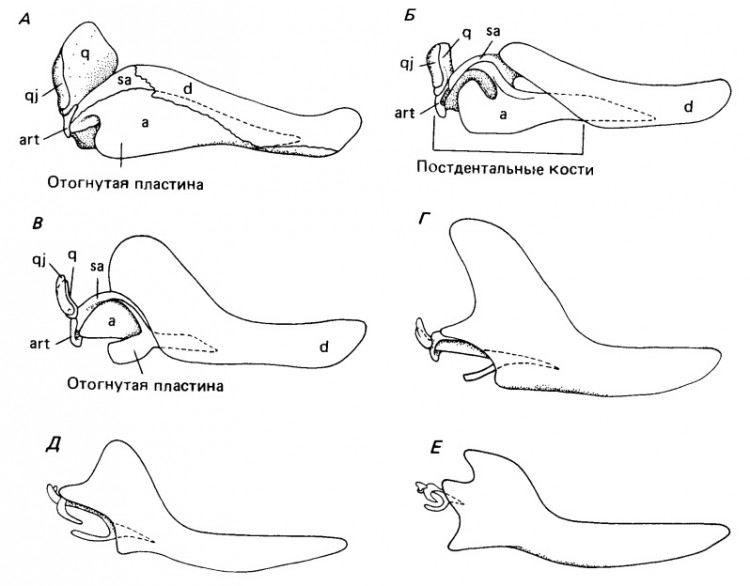
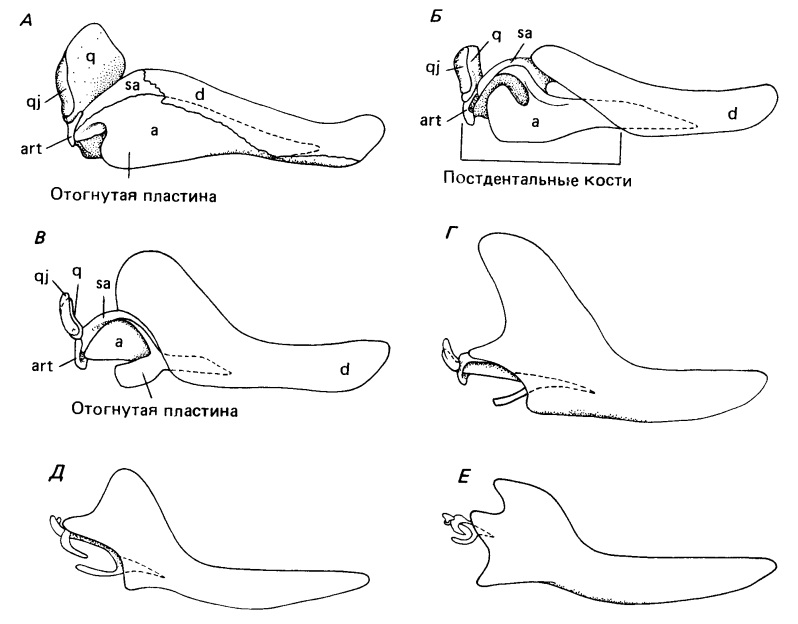
Постепенные изменения в строении нижней челюсти и элементов среднего уха при переходе от пеликозавров к млекопитающим, вид сбоку: А — сфенакодонтный пеликозавр Dimetrodon (все элементы нижней челюсти шовно соединены между собой, угловая кость несет отогнутую пластину); Б — продвинутые тероцефалы (постдентальные кости уже не соединены шовно с зубной, но остаются очень крупными); В — примитивный цинодонт Thrinaxodon; Г — продвинутый цинодонт Probainognathus; Д — раннееюрское млекопитающее Morganucodon; Е — гипотетическая реконструкция юрского "пантотерия" (слуховые косточки у непосредственных предков плацентарных и сумчатых неизвестны, но они, по-видимому, были примерно такого размера).
Все современные млекопитающие отличаются от других амниот наличием 3 слуховых косточек: молоточка и наковальни в дополнение к стремени, гомологичному единственной слуховой косточке (stapes) рептилий и птиц. Как показал Манли (1972 г.), острота слуха у птиц с единственной косточкой не хуже, чем у млекопитающих, так что появление дополнительных косточек, не давая заметного селективного преимущества, указывает на совершенно различную эволюционную историю этих групп. Форма задней стороны черепа, а также направленность и крупный размер стремени у ранних пеликозавров говорят о том, что оно не могло принимать участия в системе выравнивания акустических импедансов, как у ящериц, крокодилов или черепах. Возможно, пеликозавры воспринимали громкие низкочастотные звуки всей поверхностью щек и нижней челюсти, как современные безухие ящерицы и змеи, максимальная чувствительность которых приходится на диапазон 200-500 Гц. Как и у предков других групп амниот, включивших стремя в среднее ухо, у пеликозавров укрепилось соединение мозговой коробки с крышей черепа, что освободило стремя от поддерживающей роли, которую оно играло у ранних амниот. По какой-то причине у синапсид стремя осталось массивным, и никаких свидетельств наличия у них барабанной перепонки рептилийного типа, прикрепленной к задней части щеки, нет. Неудача в развитии барабанной перепонки в этом месте у предков млекопитающих, возможно, связана с формой задней стороны черепа и нижней челюсти у пеликозавров, особенно у сфенакодонтов, не такой, как у предков ящериц, черепах и крокодилов. У сфенакодонтов и ранних терапсид челюстной сустав и дистальный конец стремени расположены ниже уровня зубного ряда, медиальнее заднего конца нижней челюсти. Если звук должен передаваться к внутреннему уху только по стремени, наиболее подходящей зоной для его приема была бы поверхность нижней челюсти, а не задняя часть щеки, как у обычных рептилий.
Первое свидетельство наличия специализированной области для восприятия воздушных колебаний у пеликозавров — отогнутая пластина угловой кости у ранних сфенакодонтовых пеликозавров. Трудно допустить, что она эффективно передавала звук на этой стадии эволюции синапсид, хотя позднейшая роль угловой кости в поддержании барабанной перепонки, судя по ее строению у поздних цинодонтов и ранних млекопитающих, почти бесспорна. Все же Аллин (1975 г.) предполагает действии отогнутой пластины у пеликозавров наподобие барабанной перепонки. Для эффективности передачи звука пространство под ней должно было заполняться воздухом, что могло бы обеспечиваться дивертикулом евстахиевой трубы, заходящим в эту область по поверхности крыловидной мышцы.
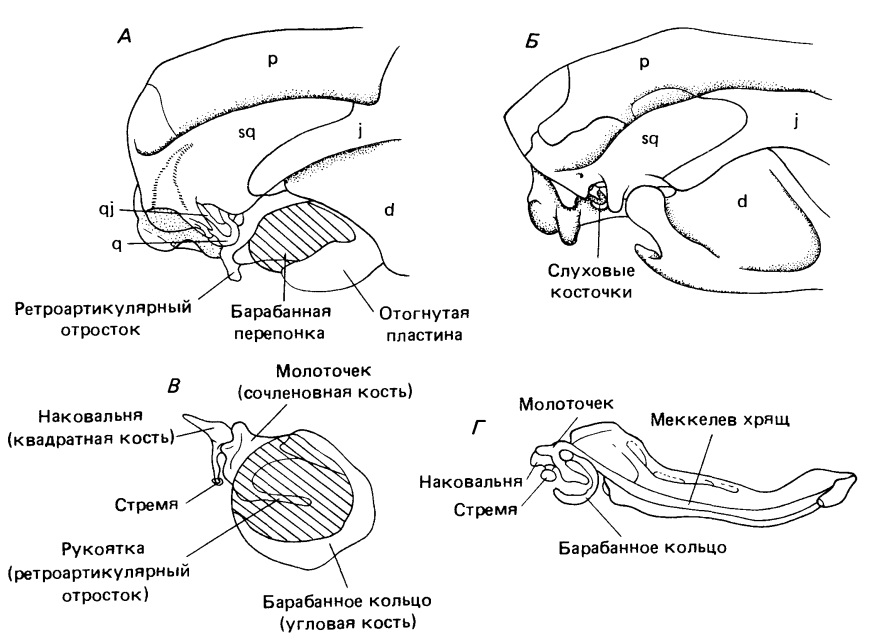
А — область нижней челюсти и уха цинодонта Thrinaxodon и (Б) современного опоссума. В — увеличенное изображение слуховых косточек опоссума. Г — медиальный вид нижней челюсти зародыша млекопитающего, демонстрирующий параллелизм между онтогенетическим развитием среднего уха и челюстного сустава с филогенетическим происхождением этих структур.
Значение заднего конца нижней челюсти для слуха гораздо легче понять у ранних цинодонтов с явно подвижными постдентальными костями. У Procynosuchus и Thrinaxodon широкая отогнутая пластина угловой кости четко отделена от тела этой кости. Существует предположение, что все постдентальные элементы колебались как единое целое. Движение сочленовной кости, по-видимому, передавалось квадратной, которая лишь рыхло прикреплялась к чешуйчатой. Такой характер прикрепления объясним, только если допустить ее подвижность, согласованную с колебаниями костей нижней челюсти. В свою очередь квадратная кость вполне могла активировать стремя, все еще остающееся крупным элементом, но облегченное наличием очень крупного отверстия для стапедиальной артерии.
У более поздних цинодонтов — как растительноядных Traversodontidae и Tritylodontidae, так и хищных Chiniquodontidae и Tritheledontidae — постдентальные кости редуцированы еще сильнее. Угловая уменьшается до узкого стержня, лежащего параллельно предсочленовной и надугловой костям. Отогнутая пластина состоит из узкого отростка, идущего постеровентрально в сторону ретроартикулярного отростка сочленовной кости. Вместе эти отростки образуют незамкнутое кольцо. По-видимому, широкая костная поверхность отогнутой пластины у ранних цинодонтов была замещена мембраной (барабанной перепонкой), простиравшейся до сочленовной кости. Эти элементы образуют структуру, сравнимую, за исключением крупных размеров, с барабанным кольцом и молоточком современных сумчатых. Молоточек сочленяется с наковальней точно так же, как и сочленовная кость с квадратной у продвинутых терапсид, а квадратная кость (наковальня) сочленяется со стременем.
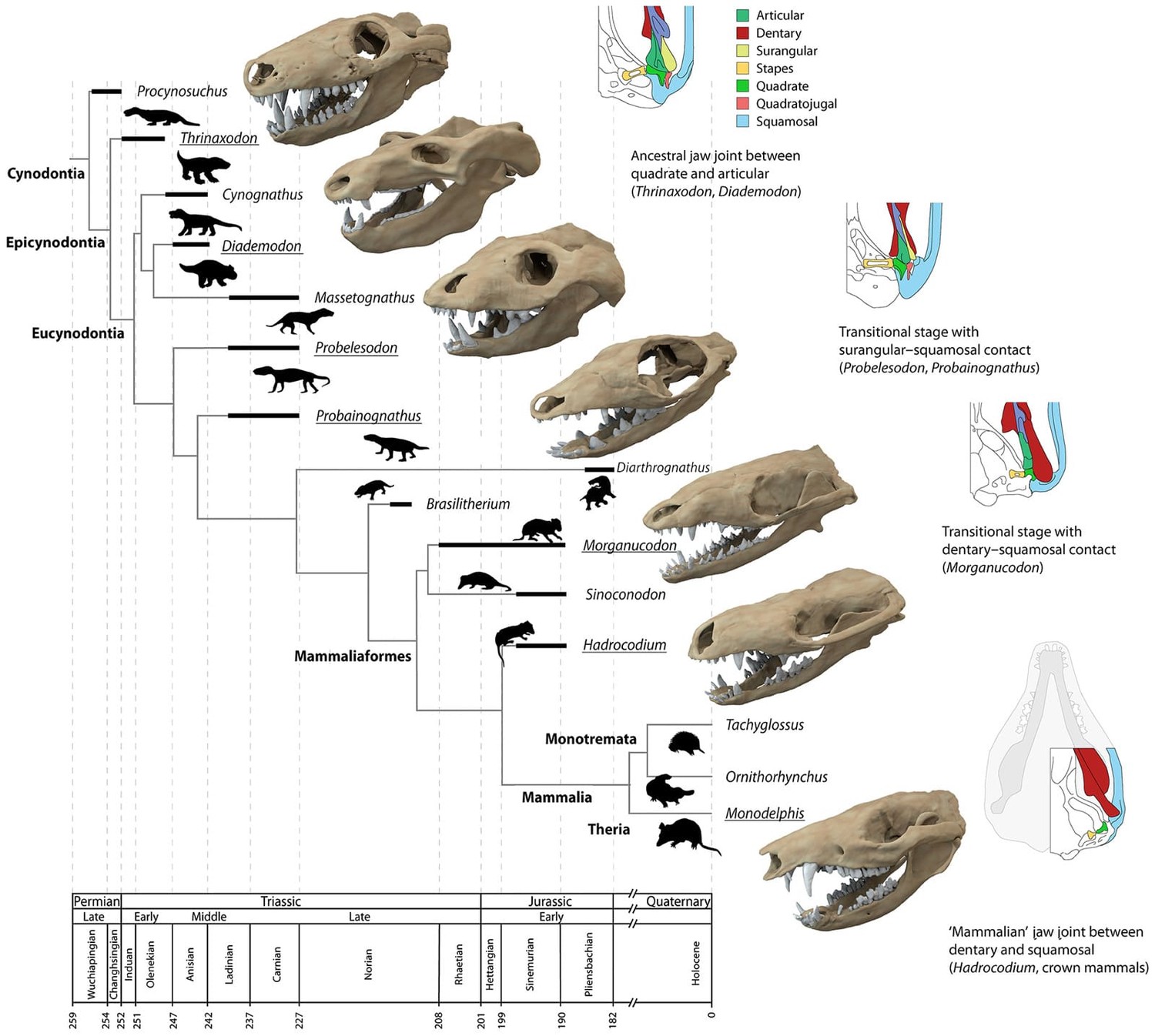
Эволюция челюстного сустава в эволюционной линии млекопитающих: у раннетриасового тринаксодона (Thrinaxodon liorhinus) имелся примитивный челюстной сустав между квадратной и сочленовной костями, у позднетриасового морганукодона (Morganucodon sp.) сустав был двойным, а у раннеюрского гадрокодия (Hadrocodium wui) нижняя челюсть крепилась к черепу так же, как у современных млекопитающих.
Слуховые косточки у взрослых млекопитающих функционально весьма своеобразны, поскольку не служат элементами челюстного сустава, как у всех терапсид. Их функции, указывающие на описанные эволюционные преобразования, изменяются у каждого поколения сумчатых. В ходе их развития в сумке молоточек и наковальня выполняют рептилийную роль сочленовной и квадратной костей. Лишь когда молодые особи покидают сумку, они отделяются от нижней челюсти и включаются в состав среднего уха.
У Probainognathus надугловая и зубная кости разрастаются назад до чешуйчатой, образуя вторичный челюстной сустав. По мере дальнейшего развития зубной кости можно различить два функциональных челюстных сустава: медиальный рептилийный из сочленовной и квадратной костей и латеральный, типичный для млекопитающих, — из зубной и чешуйчатой. Оба сохраняются у ранних млекопитающих. До сих пор не установлено, когда молоточек и наковальня вошли в состав среднего уха, но желоб на внутренней поверхности зубной кости, отмечающий место прикрепления постдентальных костей у раннеюрских млекопитающих, у позднеюрских родов отсутствует.
Челюстная механика
Давление отбора на оптимизацию восприятия воздушных колебаний должно было оставаться сильным на протяжении всей эволюции цинодонтов, противодействуя даже давлению, направленному на дальнейшее упрочнение элементов нижней челюсти.
Трудно понять, как челюсти продвинутых цинодонтов противостояли силам, приложенным к ним увеличившимся объемом челюстной мускулатуры. Брамбл (1978 г.) разработал изящную модель распределения сил, возникающих при пережевывании пищи. Долгое время признавалось, что даже у современных млекопитающих площадь челюстного сустава слишком мала и, по-видимому, не способна выдержать полное усилие челюстных мускулов. Эта проблема должна была стоять гораздо острее у продвинутых цинодонтов. Хотя силы отдельных компонентов челюстной мускулатуры действуют в разных направлениях, все они, видимо, давят на челюстной сустав, поскольку это единственная ось, вокруг которой вращается нижняя челюсть.
Важнейшая черта модели Брамбла — признание того, что место, в котором удерживается во рту пища, выступает в роли второй оси вращения. Это проще всего показать на примере умеренно продвинутого цинодонта типа Trirachodon, сконцентрировав внимание на единственной мышце — височной. Ее сила действует на венечный отросток в постеродорсальном направлении. Можно продолжить эту линию антеровентрально до пересечения с зубным рядом. Эта линия называется спроецированной линией действия. Если кусаемая пища расположена спереди от точки пересечения, сила мышцы поднимает заднюю часть нижней челюсти, создавая положительное давление на челюстной сустав. Если же пища и соответственно ось вращения находятся позади точки пересечения, задняя часть нижней челюсти опускается и на челюстной сустав действует отрицательное давление. Чем дальше сдвинута назад точка укуса, тем оно больше. Глубокая жевательная мышца действует в том же направлении, что и височная.
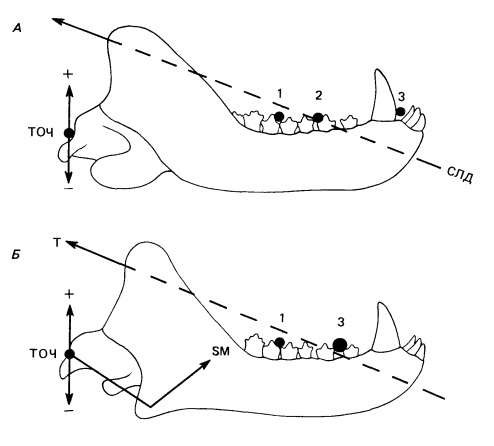
Разложение сил, действующих на челюстной сустав терапсида (на основе двуосной модели Брамбла). Сокращения: СЛД — спроецированная линия действия височной мышцы; ТОЧ — точка опоры челюстного сустава; Т — височная мышца; SM — поверхностная жевательная мышца; 1, 2, 3 — прикусные точки опоры. А. Когда прикусная точка опоры находится в положении 3 (в передней части рта), сокращение височной мышцы прилагает положительное усилие к челюстному суставу. Когда прикусная точка опоры находится в положении 1 (у заднего конца челюсти), на челюстной сустав действует отрицательная сила. Когда прикусная точка опоры находится непосредственно на спроецированной линии действия мышцы, приложенная к суставу сила нейтральна. Б. Уровновешивание сил, генерируемых височной и поверхностной жевательной мышцами. Поверхностная жевательная мышца, сила которой изображена стрелкой SM, действует почти под прямым углом к височной (Т). Когда прикусная точка опоры находится у переднего края челюсти (3), сила, приложенная к челюстному суставу, отрицательна, а когда в положении 1 — положительна. Примерно в середине челюсти будет находиться точка опоры, при которой силы височной и поверхностной жевательной мышц нейтрализуют друг друга.
И напротив, поверхностная жевательная мышца ориентирована так, чтобы создавать уравнивающий эффект. Если рассматривать ее отдельно, максимальная положительная сила прикладывается как раз тогда, когда пища находится ближе к заднему концу челюсти, а максимальная отрицательная сила — когда ближе к переднему.
Когда пища находится у заднего конца челюсти, большая отрицательная сила, развиваемая височной и глубокой жевательной мышцами, уравновешивается положительной силой поверхностной жевательной мышцы. В принципе можно было бы уравнять их по всей длине челюсти, соответственно изменяя усилие первой или второй мышечной массы. Фактически же преобладает отрицательная сила височной мышцы, когда пища в задней части челюсти, и положительная, когда она спереди. Однако никогда сила, приложенная к челюстному суставу, не будет так велика, как могло бы быть при работе только височной или только поверхностной жевательной мышцы.
Соотношение между силами зависит от высоты венечного отростка и ориентации височной мышцы. В ходе эволюции цинодонтов точка пересечения спроецированной линии действия с нижней челюстью все более сдвигается вперед. У Probainognathus она проходит выше переднего конца челюсти, так что на челюстной сустав действует отрицательная сила независимо от места укуса.
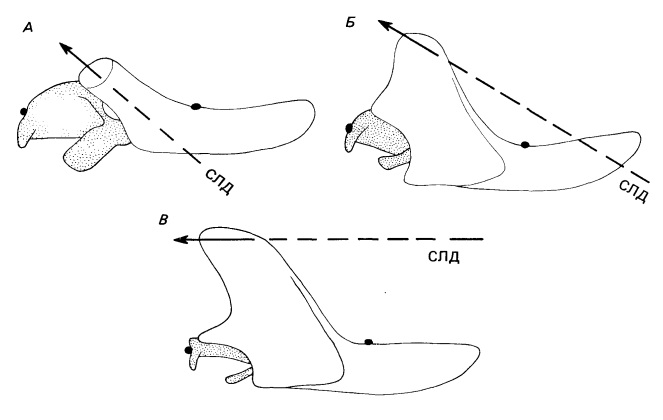
Челюсти трех цинодонтов. Показано изменение сил, действующих на челюстной сустав, при постепенном росте вверх венечного отростка. Рассмотрена только височная мышца. А. У Procynosuchus спроецированная линия мышечного действия (СЛД) пересекает нижнюю челюсть позади зубного ряда, и сила, приложенная к челюстному суставу, положительна независимо от места прикусывания пиши. Б. У Trirachodon СЛД пересекает середину зубного ряда, так что более передний укус будет создавать положительное усилие, а более задний — отрицательное. В. У Probainognathus СЛД проходит выше конца челюсти, и сила, приложенная к челюстному суставу, отрицательна независимо от того, где прикушена пища. С образованием поверхностной жевательной мышцы укус цинодонтов стал значительно сильнее, в то время как челюстной сустав уменьшился. Этот процесс объясняет, как могли редуцироваться постдентальные кости поздних терапсид в связи с эволюцией среднего уха млекопитающих. (Bramble, 1978)
Высказано предположение, что эта сила компенсируется депрессором нижней челюсти (m. depressor mandibulae), который начинается на задней стороне черепа и прикрепляется к сочленовной кости, предотвращая смещение нижней челюсти. Начало этого мускула могло помещаться внутри желоба, образованного латеральным выпячиванием чешуйчатой кости. Согласно другим воззрениям, внутри этого желоба находился наружный слуховой проход, а ретроартикулярный отросток сочленовной кости был слишком хрупким, чтобы служить местом прикрепления мощного депрессора.
Посткраниальный скелет
Конечности средне- и позднетриасового Chiniquodon значительно стройнее, чем у примитивных цинодонтов, однако строение и их самих, и их поясов не особенно продвинуто по сравнению с Thrinaxodon. Позднетриасовые и раннеюрские Tritylodontidae своим посткраниальным скелетом гораздо ближе к млекопитающим, чем среднетриасовые Chiniquodontidae.
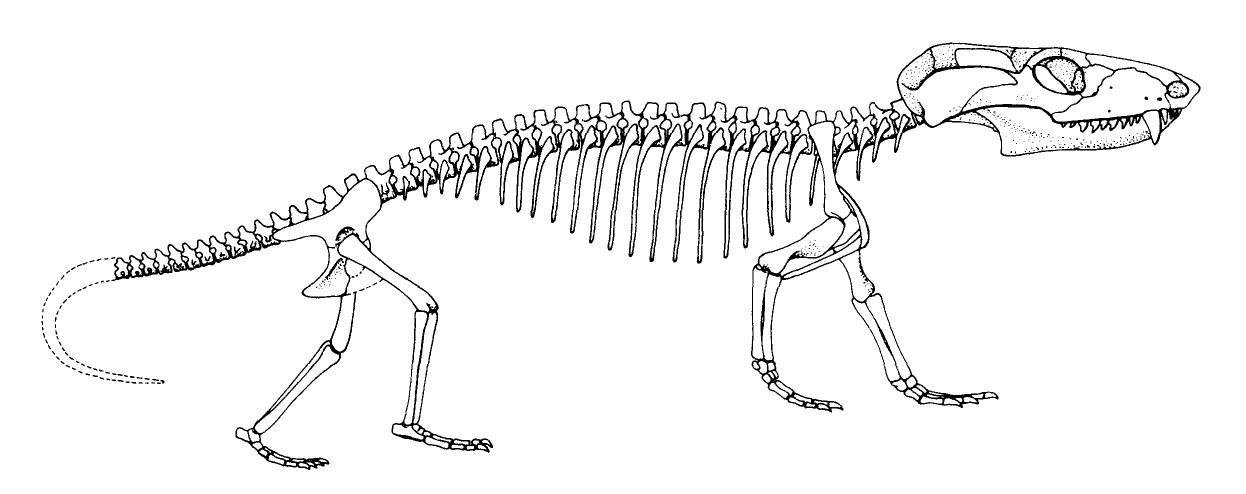
Прорисовка частично реконструированного скелета молодой особи чиникводона (Chiniquodon theotonicus), или "Probelesodon".
В обеих группах реберные пластины, столь заметные у примитивных цинодонтов, утрачены, и ребра такие же, как у большинства рептилий и ранних млекопитающих. Роль аксиальной мускулатуры в поддержании туловища, по-видимому, гораздо важнее. Утрачены даже дополнительные зигапофизные сочленения, обычные у более примитивных форм. Поразительно, что различия между грудными и поясничными ребрами, заметные у Thrinaxodon, исчезли у большинства продвинутых цинодонтов, хотя у Tritylodontidae наблюдается четкая разница между грудными и поясничными позвонками.
Тело эпистрофея у Tritylodontidae по форме становится таким же, как у млекопитающих, с развитием крупного переднего зубовидного отростка, который только намечается у ранних цинодонтов. Однако дуга атланта и гипоцентр еще не слились друг с другом. Концы тел плоские (амфиплатные), как у млекопитающих, с утратой между ними хордальных остатков, соответствующих амфицельным позвонкам более примитивных синапсид. У Tritylodontidae всего 2 крестцовых позвонка, как и у ранних млекопитающих, а следующие за ними хвостовые позвонки удлинены и образуют длинный, но тонкий хвост.
Оставаясь еще гораздо более примитивным, чем у плацентарных и сумчатых, строение их плечевого пояса вплотную приближается к характерному для примитивных раннеюрских млекопитающих. Коракоидные элементы редуцированы сильнее, чем у ранних цинодонтов, а гленоидная впадина очень широко открыта наружу. Акромион и основание переднего края пластины лопатки отклонены латерально от переднего края коракоидной области, по-видимому обеспечивая проход для части надкоракоидной мышцы (m. supracoracoideus), которая, как считается, отходила от медиальной поверхности лопаточной пластины. Этот мускул гомологичен надостной мышце (m. supraspinatus) продвинутых млекопитающих и помогает стабилизировать плечевой сустав, когда плечевая кость удерживается в более вертикальном положении.
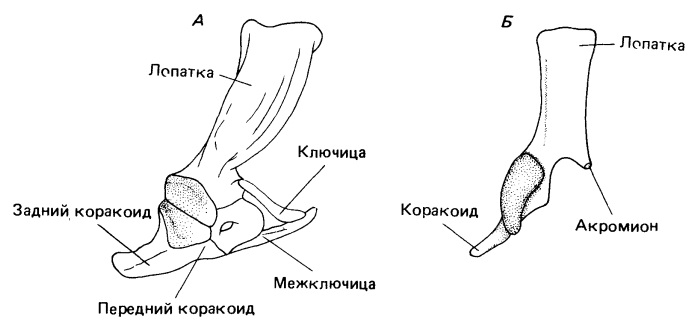
Изменения в плечевом поясе продвинутых терапсид: А — умеренно продвинутый цинодонт; Б — тритилодонт Oligokyphus.
У плечевой кости длинный диафиз и крупная полусферическая головка. Судя по всему, ориентация передней конечности у Tritylodontidae стала такой же, как у примитивных млекопитающих.
Тазовый пояс и задняя конечность раннеюрских Tritylodontidae внешне выглядят так же, как у млекопитающих. Подвздошная кость длинная и направлена антеродорсально, ее продолжение назад, обычное для ранних синапсид, отсутствует. Гребень, проходящий по всей длине латеральной поверхности этой кости, разделяет места отхождения ягодичной (подвздошно-бедренной) мышцы (m. gluteus) сверху и подвздошной (m. iliacus) и поясничной (m. psoas) (внутренней лобково-седалищно-бедренной) мышц снизу. Лобковая и седалищная кости развернуты назад так, что большей частью находятся позади вертлужной впадины. Эти три скелетных элемента остаются у всех цинодонтов самостоятельными зонами окостенения, но полностью сливаются у млекопитающих. Запирательное отверстие, начавшее развиваться у ранних цинодонтов, крупное.
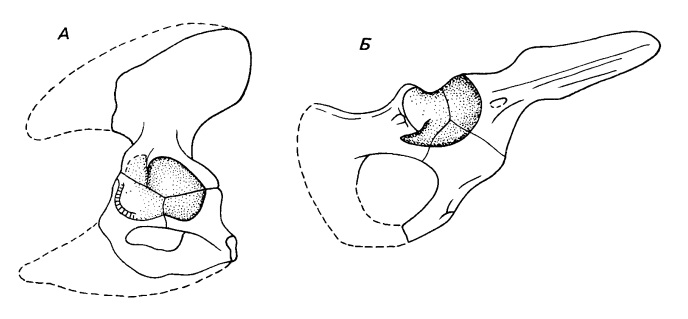
Изменения в тазовом поясе при переходе от примитивных к продвинутым терапсидам: А — примитивный цинодонт Thrinaxodon; Б — тритилодонтид Oligokyphus.
Бедренная кость Tritylodontidae практически прямая с крупной, медиально загнутой полусферической головкой. Положение и движение задней конечности, вероятно, близки к свойственным примитивным современным млекопитающим.
Таким образом, начальные стадии развития большинства признаков, характеризующих скелет млекопитающих, хорошо заметны у цинодонтов из раннего и среднего триаса. К сожалению, в позднем триасе сведения о непосредственных предках млекопитающих становятся менее полными. Tritheledontidae дают лишь смутное представление о мелких позднетриасовых формах. Tritylodontidae же прекрасно демонстрируют заключительные стадии эволюции терапсид, но высокоспециализированная зубная система отдаляет их от предков млекопитающих.
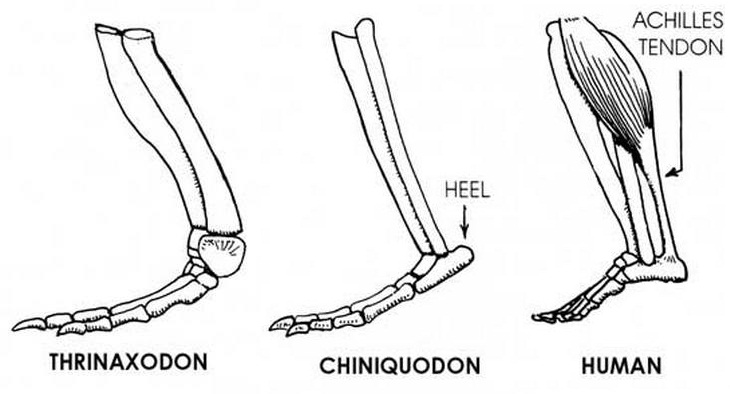
Формирование у цинодонтов выступа пяточной кости, к которому крепится ахиллесово сухожилие. Автор — Д. Петерс.