Цинодонты и происхождение млекопитающих
Несмотря на то что ряд признаков, присущих млекопитающим, проявляется и в других группах терапсид, только цинодонты по своей общей морфологии существенно приблизились к схеме их строения. Череп цинодонтов продвинут по сравнению с другими терапсидами по комплексу признаков, предвосхищающих его состояние у млекопитающих. Чтобы оценить их значение, необходимо рассмотреть некоторые из основных отличий последних от рептилий.
Особенно важными чертами современных млекопитающих обычно считаются крупный мозг и живорождение. Плацентарный тип живорождения появился предположительно уже в среднеюрское время, однако размер мозга заметно увеличился лишь в течение кайнозоя (размеры больших полушарий у триасовых цинодонтов и тероцефалов значительно увеличились по сравнению с более примитивными их представителями из перми, хотя оставались небольшими по сравнению с юрскими и меловыми млекопитающими).
Значительно более фундаментальный признак млекопитающих — высокая интенсивность их метаболизма. Современные формы потребляют примерно в 10 раз больше пищи и кислорода, чем рептилии такого же размера. Более интенсивный обмен веществ позволяет млекопитающим сохранять активность в течение более длительных периодов времени и поддерживать высокую, постоянную температуру тела, независимую от внешней среды. Такая радикальная физиологическая перестройка влияет практически на все системы организма и прямо или косвенно обусловливает почти все различия, наблюдающиеся между рептилиями (и современными, и палеозойскими) и млекопитающими.
Изменения могли начаться со сдвига в количественном соотношении анаэробного и аэробного метаболизма произвольной мускулатуры. Для энергоснабжения мышечных сокращений рептилии используют в первую очередь анаэробное разложение глюкозы. Она присутствует в самих мышцах, реакция может происходить сразу и слабо зависит от температуры. За счет анаэробного метаболизма рептилии могут пробегать короткие расстояния так же быстро, как и млекопитающие. Важнейший его недостаток — накопление молочной кислоты, приводящее к мышечной усталости уже через 1-2 мин энергичной работы. После нескольких коротких пробежек ящерица почти неспособна к дальнейшим усилиям. Распад молочной кислоты требует нескольких часов; лишь после этого мышцы вновь готовы активно сокращаться.
Млекопитающие также используют анаэробный метаболизм для коротких быстрых «рывков», однако длительная локомоция обеспечивается у них в большей мере аэробными процессами. Они дают существенно больше энергии с образованием в качестве «отходов» только воды и двуокиси углерода. В результате мышцы могут энергично сокращаться часами без заметной усталости.
Более вертикальное положение конечностей и структурные адаптации для их более эффективного движения вперед-назад у терапсид, как и у динозавров, возможно, связаны с повышенной способностью поддерживать продолжительную мышечную активность без утомления. Судя по строению конечностей и их поясов, не исключено, что даже у эотитанозухид интенсивность метаболизма была значительно выше, чем у пеликозавров и других примитивных амниот. Трудно объяснить вертикальное положение конечностей многих горгонопов и поздних терозавров, если не предполагать у них способности к длительной локомоторной активности.
Высокая интенсивность метаболизма является значительным преимуществом. Однако ее выгоды обходятся дорогой ценой. Современные рептилии потребляют гораздо меньше пищи и могут обходиться без нее долгое время. Они легче приспосабливаются к заметным сезонным и суточным колебаниям температуры, поскольку температура их тела меняется без отрицательных последствий для организма.
Высокая интенсивность обмена веществ требует значительного и надежного пищевого снабжения. Все ранние амфибии и рептилии были хищниками. В самой поздней пеликозавровой фауне с доминированием казеид впервые большинство составляют травоядные формы. Травоядные дицинодонты гораздо многочисленнее хищных в поздней перми и раннем триасе. Наличие большого числа первичных консументов, возможно, обеспечило достаточно надежный источник пищи, чтобы послужить основой для достижения хищными терапсидами относительно высокой интенсивности метаболизма.
Это требует также постоянного снабжения кислородом и быстрого удаления углекислого газа, а значит — большей эффективности кровообращения и дыхания, чем у современных рептилий. Можно предположить, что у предков млекопитающих возросла способность к газообмену в легких и произошло полное разделение желудочков сердца, т. е. прекратилось смешивание оксигенированной и неоксигенированной крови. Однако ни одно из этих изменений не отразилось непосредственно на скелете, и можно только догадываться о степени их выраженности у терапсид.
Другие аспекты повышения интенсивности метаболизма проявляются в скелетной анатомии более явно. Постоянное снабжение легких млекопитающих кислородом обеспечивается отделением воздухоносных путей от ротовой полости за счет развития протяженного вторичного неба. Потребность в дополнительной пище удовлетворяется более эффективной кормовой стратегией и ускорением переваривания. Современные рептилии заглатывают пищу целиком или большими кусками, и пищеварение идет медленно. Птицы используют мускульный желудок для измельчения корма, что способствует его более быстрому перевариванию. У продвинутых терапсид и млекопитающих пища размельчается во рту с помощью специализированной зубной системы в ходе продолжительного пережевывания.
Для цинодонтов особенно характерны сложные зубы, особая челюстная механика и модификация челюстной мускулатуры. Ни одного из этих изменений не наблюдается у горгонопов. У некоторых тероцефалов обширное вторичное небо и сложные заклыковые зубы, но ни у одного из них не заметно изменений в распределении челюстной мускулатуры. Зато они отмечаются уже у древнейших известных цинодонтов. Дополнительные преобразования последовательно происходят в этой группе в течение всего триаса.
Procynosuchus и происхождение челюстной мускулатуры млекопитающих
Позднепермско-раннетриасовый Procynosuchus уже заметно продвинут в развитии вторичного неба, сложных заклыковых зубов, а также в строении челюстной и височной областей, свидетельствующем о начале реорганизации челюстной мускулатуры в сторону схемы, свойственной млекопитающим.
Вторичное небо у Procynosuchus сформировано вентральным и медиальным разрастанием предчелюстных, верхнечелюстных и небных костей. Увеличенные участки не контактируют по средней линии и не продолжаются до конца зубного ряда. Однако отделение воздухоносных путей от ротовой полости, вероятно, завершало вторичное небо из мягких тканей. Заклыковые зубы многобугорчатые и внешне напоминают моляры позднетриасовых млекопитающих. Их коронки удлиненные, с рядом линейно расположенных бугров. Верхние и нижние зубы не точно смыкаются друг с другом и постоянно заменяются, как у большинства рептилий.
Большое височное окно указывает на наличие очень крупных мускулов, смыкающих челюсти. В отличие от других групп терапсид нижняя височная, или скуловая, дуга выступает наружу за уровень зубного ряда. Это позволяет челюстным мускулам прикрепляться к латеральной поверхности зубной кости. Такая же схема свойственна и дицинодонтам, но как результат поднятия скуловой дуги над уровнем зубного ряда, а не ее латерального выпячивания. Зубная кость цинодонтов несет высокий венечный отросток с латеральной выемкой для прикрепления мускула. У млекопитающих он называется глубокой жевательной мышцей (m. masseter profundus), а выемка — ямой жевательной мышцы (fossa masseterica).
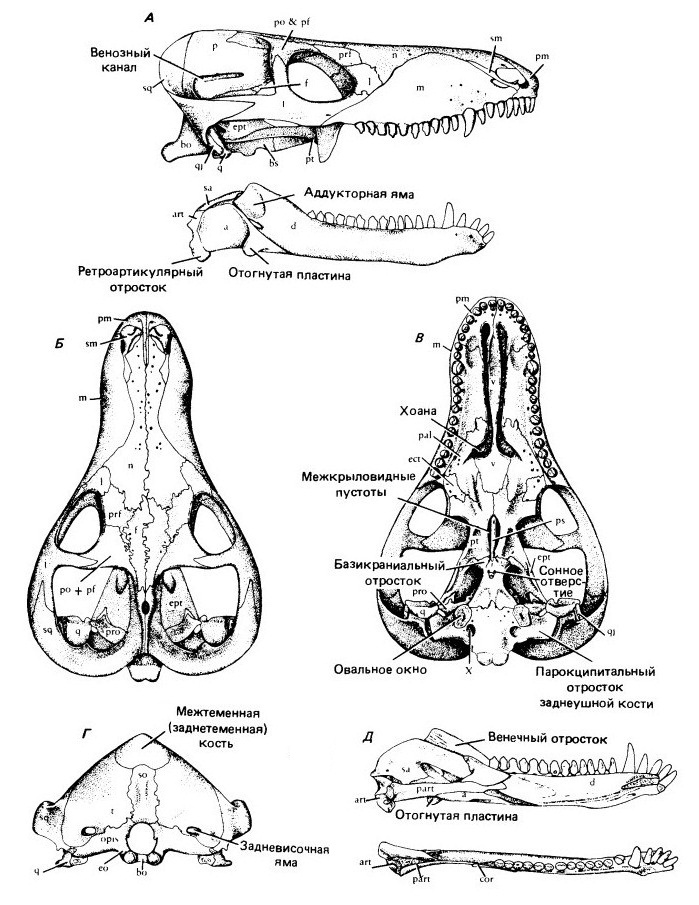
Прорисовки реконструированного черепа процинозуха (Procynosuchus delahareae): А — череп и нижняя челюсть латерально, Б — сверху, В — с неба, Г — сзади, Д — нижняя челюсть медиально и со стороны прикуса.
У примитивных рептилий и амфибий имеется три главных аддуктора нижней челюсти: задний, наружный и внутренний, включающий ложновисочную (m. pseudotemporalis) и крыловидную (m. pterygoideus) мышцы. Судя по очертаниям черепа и нижней челюсти, у пеликозавров и ранних терапсид сохраняется эта примитивная схема. У более поздних синапсид задний аддуктор нижней челюсти и ложновисочная часть внутреннего аддуктора сильно редуцируются. Большая часть челюстной мускулатуры млекопитающих и все мышцы височной области произошли от наружного аддуктора рептилий. У цинодонтов зона прикрепления его наружного слоя смещается на латеральную поверхность зубной кости, и этот слой становится жевательной мышцей. Ее начало переходит на внутреннюю поверхность скуловой дуги. Мышца, сохраняющая свое место прикрепления на медиальной и дорсальной поверхностях венечного отростка и начинающаяся на крыше черепа и латеральной стенке мозговой коробки, называется височной (m. temporalis).
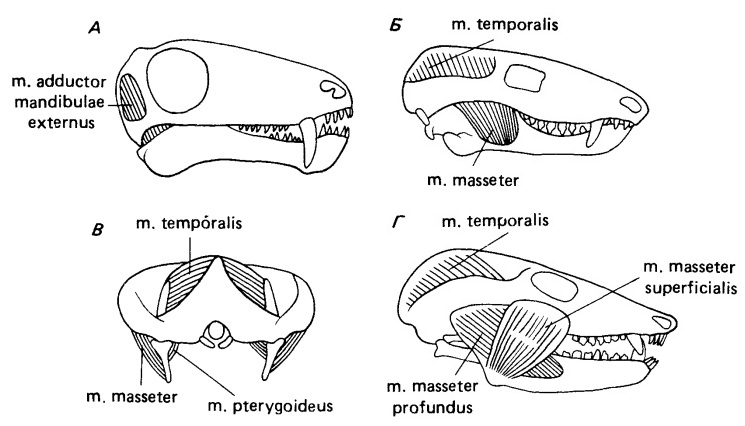
Эволюция челюстной аддукторной мускулатуры млекопитающих. Из 3 основных элементов аддукторной мускулатуры примитивных тетрапод внутренний и задний аддукторы нижней челюсти подверглись у предков млекопитающих очень сильной редукции. Все крупные мускулы, берущие начало в аддукторной камере млекопитающих, произошли от наружного аддуктора нижней челюсти. У примитивных терапсид типа Biarmosuchus (А) наружный аддуктор, как и у примитивных амниот, состоит из единственной функциональной единицы, которая полностью помещается внутри аддукторной камеры и прикрепляется к дорсальной и медиальной поверхности задней стороны нижней челюсти. У примитивных цинодонтов типа Thrinaxodon (Б) скуловые дуги выгнуты наружу и поверхностная часть наружного аддуктора прикрепляется к латеральной поверхности нижней челюсти. Место его отхождения перемещается на внутреннюю поверхность скуловой дуги. С разделением аддуктора нижней челюсти на 2 функциональные единицы внутренняя его часть, берущая начало на крыше черепа и латеральной поверхности мозговой коробки, теперь называется височной мышцей, а латеральная часть — жевательной. Сзади (В) заметно, что сила жевательной мышцы тянет нижнюю челюсть латерально и вверх, уравновешивая таким образом силу височной мышцы, направленную медиально. У более продвинутых цинодонтов типа Probainognathus (Г) жевательная мышца разделяется на 2 части. Волокна глубокой жевательной мышцы, как и единый мускул у Thrinaxodon, ориентированы почти параллельно волокнам височной мышцы, а у новой поверхностной жевательной мышцы они направлены косо вперед.
Увеличившийся размер височного окна у ранних цинодонтов оставляет большее пространство для челюстной мускулатуры, в результате чего усиливается укус. Развитие жевательной мышцы также перераспределяет силы, приложенные к нижней челюсти, позволяя (в отличие от примитивных групп рептилий) управлять ее медиолатеральным движением. Этот контроль становится очень важным при установлении точного смыкания коренных зубов у млекопитающих.
Thrinaxodon и ранние стадии развития скелета млекопитающих
Из всех цинодонтов наиболее хорошо известен раннетриасовый Thrinaxodon. Детальное изучение его черепа и посткраниального скелета дало хорошее представление о терапсидах, близких к предкам млекопитающих.
Череп
Почти половину длины черепа у Thrinaxodon занимает увеличенная аддукторная камера. Очень узкая черепная коробка несет острый сагиттальный гребень, еще сохраняющий пинеальное отверстие. Чешуйчатая кость широко охватывает сзади аддукторную камеру и образует латеральную поверхность затылка. Как и у Procynosuchus, на месте заглазничной и заднелобной костей более примитивных терапсид находится единственное окостенение, встречающееся с предлобной костью над глазницей.
Thrinaxodon продвинут по сравнению с Procynosuchus в связи с формированием сплошного вторичного неба с шовным соединением верхнечелюстных и небных костей по средней линии под носовым проходом. У взрослых особей крыловидные кости встречаются на средней линии и закрывают межкрыловидную полость.
Нижняя челюсть состоит в основном из зубной кости с венечным отростком, заходящим дорсально выше скуловой дуги. Яма жевательной мышцы достигает его основания. Постдентальные кости редуцируются не сильно, но, как и у Procynosuchus, утрачивают шовное соединение с зубной костью и могут двигаться отдельно от нее. Отогнутая пластина угловой кости по-прежнему остается крупным образованием, латерально выступающим над остальной костной поверхностью. Как у примитивных рептилий, сочленовная кость полностью образует сочленение с черепом. Квадратная и тесно связанная с ней квадратноскуловая кости сильно редуцированы и рыхло закреплены в вырезках у основания чешуйчатой кости.
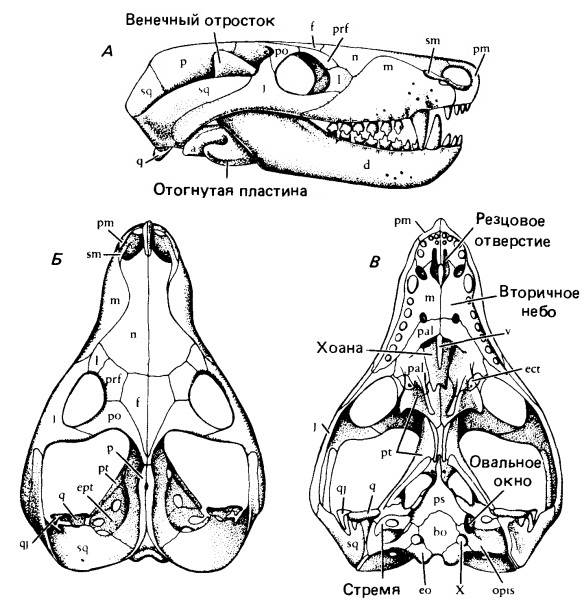
Прорисовки реконструированного черепа тринаксодона (Thrinaxodon liorhinus): А — сбоку, Б — сверху, В — с неба.
Зубная система почти такая, как у млекопитающих. С каждой стороны находятся 4 верхних и 3 нижних резца. Это число сохраняется и у большинства более продвинутых цинодонтов. Клыки присутствуют на верхней и нижней челюстях, хотя они не такие крупные, как у многих горгонопов. Заклыковых зубов — от 7 до 9. Коронки латерально сжаты, несут ряд линейно расположенных бугров. Однако в отличие от млекопитающих зубы все еще регулярно сменяются и не демонстрируют специфического прикуса.
Первые тенденции к повышению интенсивности метаболизма среди терапсид привели к отбору на более совершенный посткраниальный скелет, облегчающий быструю и маневренную локомоцию. В течение всей эволюции цинодонтов и мезозойских млекопитающих идут постепенные изменения морфологии позвонков, конечностей и их поясов. Начальные стадии этих изменений отчетливо заметны у Thrinaxodon.
Происхождение комплекса атлант - эпистрофей
У терапсид подвижность головы относительно туловища заметно изменилась. Схема примитивных амниот доходит с небольшими модификациями через пеликозавров до примитивных терапсид. Затылочный мыщелок представляет собой примерно полусферическую структуру, расположенную под большим затылочным отверстием (foramen magnum). Дуги атланта и гипоцентр (= интерцентр) образуют незамкнутое кольцо, окружающее мыщелок, как впадина в шарнирном суставе. Подвижность этого узла ограничена проатлантом, парные элементы которого служат связующим звеном между сочленовными поверхностями боковых затылочных костей и парными дугами атланта. Эта связь исключает как вращение, так и дорсовентральное сгибание на границе между черепом и позвоночным столбом.
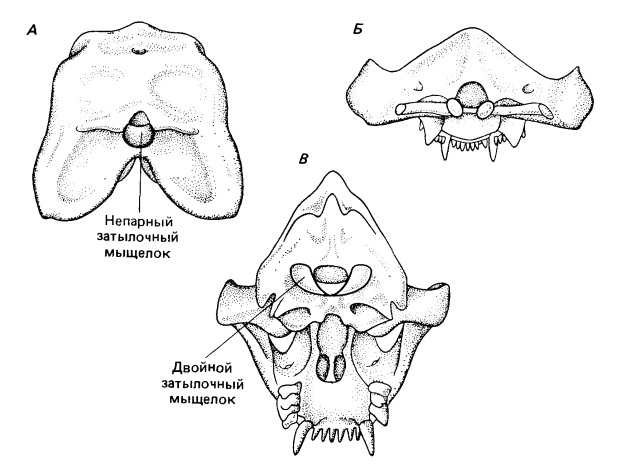
Сочленение между черепом и шейными позвонками значительно изменяется при переходе от пеликозавров к млекопитающим. А. У примитивных пеликозавров затылочный мыщелок представляет собой единую полусферическую структуру, расположенную непосредственно под большим затылочным отверстием. Б. У терапсид мыщелок разделяется, образуя парную структуру. В. Место сочленения перемещается у млекопитающих вверх, так что ось сустава проходит на половине высоты большого затылочного отверстия, а это уменьшает натяжение спинного мозга, когда череп отклоняется вверх или вниз. Раздвоение затылочного мыщелка препятствует боковым отклонениям или вращению в этом месте.
У примитивных терапсид подвижность ограничивалась небольшой способностью к вращению и сгибанию в каждой точке сочленения между черепом и передними элементами шейного отдела. У млекопитающих два этих вида движений происходят в основном между различными элементами комплекса атлант - эпистрофей. Дорсовентральное сгибание по дуге до 90° осуществляется в первую очередь между затылочным мыщелком и атлантом. У цинодонтов и ранних млекопитающих оно обеспечивается разделением первоначально единого затылочного мыщелка на две сочленовные поверхности, постепенно смещающиеся из положения под затылочным отверстием в обе стороны от него. У примитивных амниот сгибание в этом суставе привело бы к значительному растягиванию верхней стороны спинного мозга и сдавливанию его нижней половины. Поскольку мыщелки сдвинулись вверх до середины высоты большого затылочного отверстия, эти действующие на спинной мозг силы сводятся к минимуму. Однако парная природа мыщелковой поверхности препятствует вращению и боковым изгибам в этом суставе. У Thrinaxodon мыщелок отчетливо парный, но проатлант сохраняется. У всех цинодонтов сочленовные поверхности все еще остаются в относительно вентральном положении.
Исключение вращения между головой и атлантом компенсируется специализацией сочленения атлант - эпистрофей. У примитивных терапсид вращение между этими элементами ограничено из-за хорошо развитых зигапофизных суставов между их дугами. У ранних цинодонтов зигапофизы редуцированы. Кроме того, тело атланта соединяется швом, а позже и сливается с телом II-го позвонка с образованием выступа (зуба эпистрофея), вокруг которого вращаются дуги атланта, не утрачивающие связи с его гипоцентром. Последние элементы не соокостеневали ни у продвинутых цинодонтов, ни у древнейших млекопитающих, но, вероятно, прочно соединялись хрящом и связками. У современных млекопитающих они формируют функциональное кольцо.
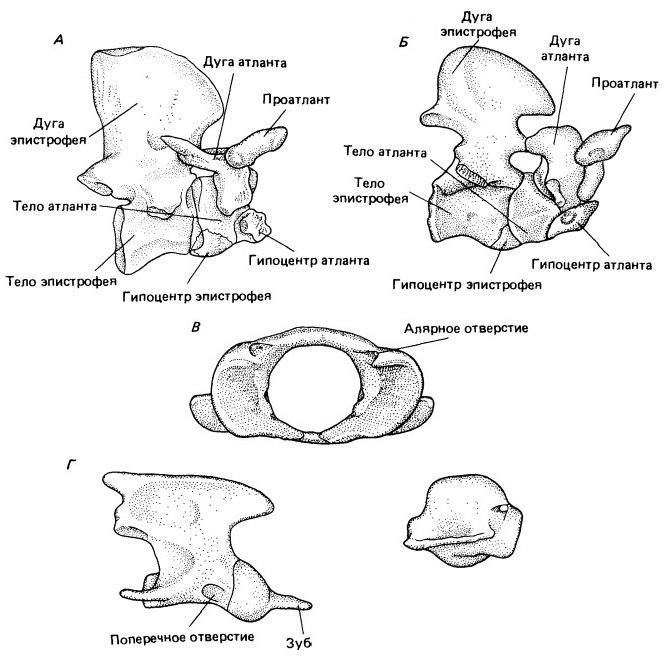
Изменения в комплексе атлант - эпистрофей при переходе от пеликозавров к млекопитающим. А. Пеликозавр Ophiacodon. Ограниченное сгибание и вращение возможно в ряду суставов между затылочным мыщелком и шейными позвонками. Связь между проатлантом и затылком ограничивает дорсовентральное сгибание и вращение между черепом и атлантом. Дуга атланта препятствует вращению между атлантом и эпистрофеем. Б. Цинодонт Thrinaxodon. Зигапофизное сочленение между атлантом и эпистрофеем упрощено и допускает вращение между этими элементами. Плевроцентры атланта и эпистрофея объединяются, образуя ось, вокруг которой могут вращаться дуга и гипоцентр атланта. В, Г. Современные млекопитающие: В — вид спереди атланта кошки; Г — вид сбоку комплекса атлант - эпистрофей (разделен, чтобы были видны сочленовные поверхности). Дуга и гипоцентр атланта сливаются в кольцевидную структуру. Передняя поверхность ограничивает движение черепа сгибанием в вертикальной плоскости. Вращение и боковое сгибание локализованы между атлантом и эпистрофеем.
У примитивных синапсид и других ранних амниот шейные позвонки направлены горизонтально в сторону черепа. У цинодонтов наблюдается постепенная тенденция к их отклонению дорсально к большому затылочному отверстию, как у млекопитающих. Как и у большинства современных млекопитающих, у Thrinaxodon уже семь шейных позвонков, отличающихся сохранением гипоцентров, утрачиваемых в туловищном отделе, относительно небольшим углом зигапофизов и простотой ребер.
Туловище и хвост
Форма туловищных позвонков и ребер у терапсид и ранних млекопитающих постепенно изменяется, ограничивая боковые изгибы тела и облегчая его сгибание в сагиттальной плоскости. У Thrinaxodon на протяжении всего туловищного отдела зигапофизы круто приподняты для уменьшения боковых изгибов. Впервые заметно четкое различие между грудным и поясничным отделами в длине ребер. Насчитывается 13 грудных, 7 поясничных и 5 крестцовых позвонков. Наиболее любопытный признак туловищного отдела — разрастание проксимальных частей ребер в широкие реберные пластины. Ребра поясничного отдела состоят из одних таких пластин без дистальных стержней. Реберные пластины логически связаны с усилением жесткости позвоночного ствола, позволяющей постоянно держать туловищный отдел над землей. Их наличие способствует также уменьшению степени его боковых изгибов. Реберные пластины постепенно редуцируются в нескольких линиях более продвинутых цинодонтов, что, возможно, связано с развитием эпаксиальной мускулатуры, поддерживающей туловище без помощи расширенных ребер. Здесь уместно вспомнить, что у первых наземных позвоночных, ихтиостегид, укрепляющие туловище ребра также были крайне широкими, но редуцировались в последующих линиях амфибий.
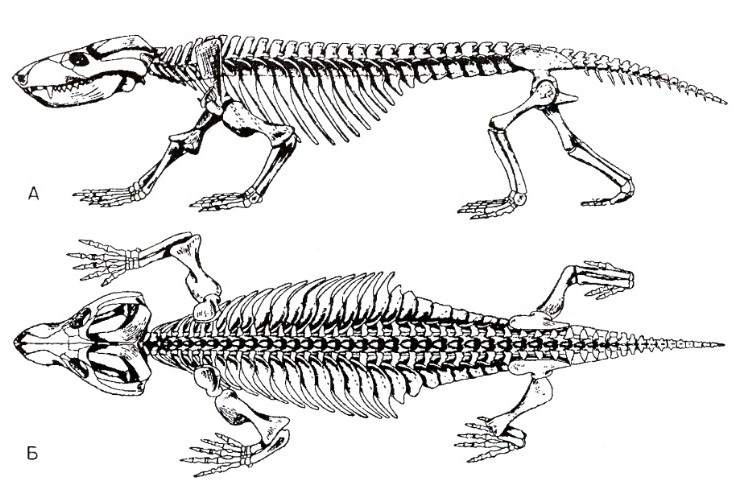
Прорисовки реконструированного скелета тринаксодона (Thrinaxodon liorhinus): А — сбоку, Б — сверху.
В отличие от более примитивных терапсид хвост у Thrinaxodon довольно короткий и включает всего 10-15 позвонков. Этот признак кажется маловажным, однако фактически укорочение хвоста связано с фундаментальными изменениями в управлении положением тела и локомоцией. И терапсиды, и текодонты произошли от примитивных амниот с «растопыренными» конечностями, но разными путями. Архозавры сохраняют длинный хвост, характерный для примитивных амниот. Когда конечности продвинутых текодонтов приобрели более габитуально вертикальное положение, поддержание выгнутой формы туловища обеспечивалось уравновешивающим действием длинного тяжелого хвоста. Его вес позволил также некоторым текодонтам и динозаврам развить факультативную и даже габитуальную бипедальность. У цинодонтов, наоборот, хвост постепенно укоротился. Развитие реберных пластин и более эффективной туловищной мускулатуры, вероятно, позволило сделать это на ранней стадии эволюции группы, но это помешало развитию у терапсид бипедальности. У некоторых из поздних форм вновь появился длинный хвост, но в виде тонкого придатка, как у современных грызунов, без тяжелой мускулатуры их предков-рептилий.
Изменения в тазовом поясе, связанные с увеличением амплитуды передне-задних движений конечностей, также заметно различаются в двух этих группах. У текодонтов и примитивных динозавров удлиняются лобковая и седалищная кости, увеличивая расстояние между протракторами и ретракторами бедра. У цинодонтов лобковая и седалищная кости уменьшаются, зато пластина подвздошной разрастается, особенно вперед, что увеличивает площадь отхождения и протракторов, и ретракторов, механика действия которых значительно отличается от свойственной динозаврам. Эти изменения у цинодонтов уже были хорошо заметны к началу триаса, когда ранний архозавр Chasmatosaurus сохранял таз, сходный с типичным для наиболее примитивных диапсид.
Скелет конечностей и их поясов
У всех цинодонтов изменения в строении поясов и конечностей связаны с более вертикальным положением последних, которое позволяло поддерживать туловище без необходимости в обычной для более примитивных амниот мощной мускулатуре, отходящей от вентральной части поясов. Происходят постепенная редукция коракоидной и лобково-седалищной пластин, а также изменения гленоидной и вертлужной впадин, повышающие их устойчивость к вертикально направленным силам, создаваемым конечностями. У цинодонтов строение тазового пояса приближается к свойственной примитивным ныне живущим сумчатым и плацентарным. Плечевой пояс эволюционирует медленнее, и передние конечности сохраняют «растопыренное» положение в течение всего триаса.
У примитивных амниот широкая вентральная пластина ключицы и крепкая Т-образная межключица обеспечивают сопротивление мощной медиально направленной силе, создаваемой направленными в сторону конечностями. Эти дермальные элементы остаются крупными у всех цинодонтов и даже у примитивных млекопитающих, показывая, что передние конечности продолжают развивать существенное медиальное усилие вследствие их «растопыренного» положения. У современных однопроходных эти кости сохранили по существу цинодонтное строение.
У всех цинодонтов происходит последовательная редукция размеров коракоида и прокоракоида. У Thrinaxodon коракоид и лопатка принимают примерно одинаковое участие в формировании гленоидной впадины, а небольшой прокоракоид образует узкий участок ее переднего края. Лопатка длинная и тонкая; она вогнута медиально в соответствии с конфигурацией туловища. По сравнению с пеликозаврами ее передний край редуцирован, но вперед отходит короткий отросток, акромион, сочленяющийся со стержнем ключицы. Лопаточная часть гленоидной впадины ориентирована вентрально и латерально, вероятно принимая на себя большую часть веса тела, действующего на переднюю конечность.
Плечевая кость остается тяжелой и сложной, как у пеликозавров. Ее головка отклонена дорсально для более эффективной передачи силы от конечности к вертикальной лопатке. Движение плеча, видимо, по-прежнему совершалось в первую очередь в горизонтальной плоскости, а работа передней конечности, вероятно, напоминала таковую у пеликозавров. Раскрытие гленоидной впадины и развитие более обособленной головки плечевой кости позволяли ей при ретракции проходить ближе к телу, что было необходимым исходным этапом развития передне-заднего парасагиттального движения конечности млекопитающих.
Тазовый пояс и задняя конечность ранних цинодонтов демонстрируют по сравнению со схемой пеликозавров целый ряд важных изменений, предвосхищающих особенности строения млекопитающих. Данные по современным рептилиям и млекопитающим свидетельствуют, что в процессе такого перехода произошли также сильные преобразования функциональной системы мускулов. У пеликозавров вертлужная впадина направлена прямо вбок. Диафиз бедренной кости прямой, а головка терминальная, что говорит о почти горизонтальном отхождении под прямым углом к туловищу. Поверхность сочленения с берцовыми костями свидетельствует о расположении дистального конца бедра даже несколько выше вертлужной впадины. Берцовые кости были ориентированы не вертикально, а под углом наружу. Это заметно по их сочленению с таранной и пяточной костями.
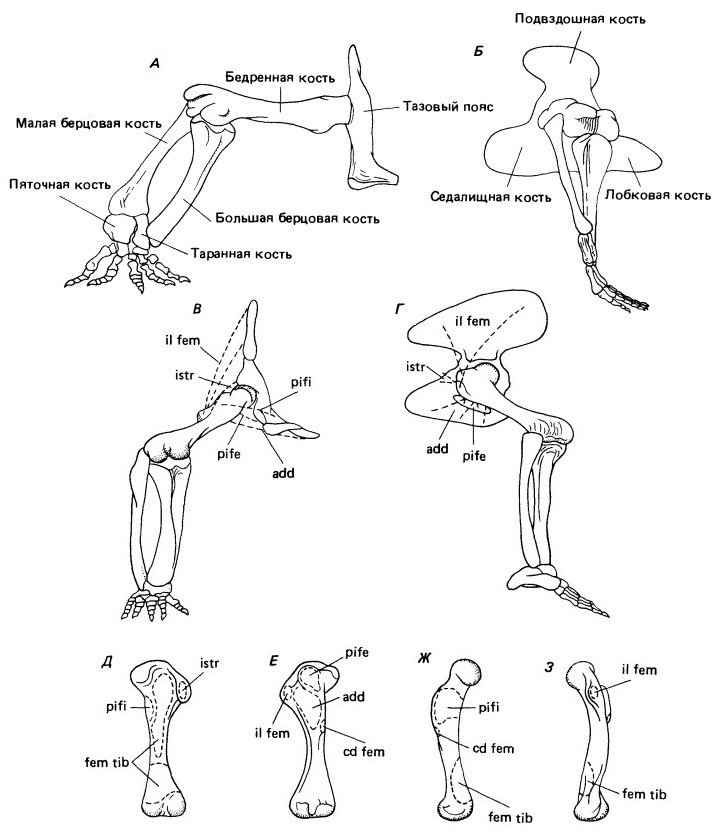
Тазовый пояс и задняя конечность. Изменения их структуры и функции при переходе от пеликозавров к цинодонтам. Вид спереди (А) и латерально (Б) таза и задней конечности пеликозавра Dimetrodon. Вид спереди (В) и латерально (Г) таза и задней конечности цинодонта Thrinaxodon. Вид (Д) сверху, (Е) снизу, (Ж) спереди и (З) сзади бедренной кости Thrinaxodon с местами прикрепления мускулов. Ориентация основных мускулов изменяется, что повышает эффективность передне-заднего движения конечности, расположенной ближе к телу. Постановка стопы при этом переходе также изменяется с пальце- на стопоходящую. У Thrinaxodon пяточная кость служит рычагом для сгибания стопы вниз.
Лобково-седалищная пластина была обширной и простиралась симметрично вперед и назад от вертлужной впадины. От ее поверхности отходили два крупных мускула, прижимающие бедро к туловищу или поднимающие тело с упором на заднюю конечность. Наружная лобково-седалищно-бедренная мышца (m. puboischiofemoralis exterior) прикреплялась в основном к внутреннему вертлугу, заметному гребню на антеровентральном крае бедренной кости дистальнее ее головки. Аддуктор прикреплялся дистальнее вдоль аддукторного гребня.
Главным ретрактором бедра, служившим для передвижения тела вперед с упором на заднюю конечность, была бедренно-хвостовая мышца (m. caudifemoralis). Она отходила от основания хвоста, тянулась горизонтально вперед и прикреплялась к высокому IV-му вертлугу (примерно на границе верхней трети длины бедренной кости на ее вентральной поверхности). Вторым ретрактором была седалищно-вертлужная мышца (m. ischiotrochantericus), отходящая от дорсальной поверхности седалищной кости и прикрепляющаяся к постеродорсальной поверхности бедренной кости сразу же под ее головкой.
Внутренняя лобково-седалищно-бедренная мышца (m. puboischiofemoralis interior), начинающаяся на дорсальной (внутренней) поверхности лобковоседалищной пластины спереди от подвздошной кости и крепящаяся на дорсальной стороне бедренной кости, служила главным протрактором, возвращающим бедро вперед после толчковой фазы. Бедро могло подниматься подвздошно-бедренной мышцей (m. iliofemoralis), бравшей начало на дорсальной поверхности бедренной кости у самой ее головки. Ориентация главных протракторов и ретракторов свидетельствует о том, что бедро двигалось в основном в горизонтальной плоскости.
У нижне- и среднетриасовых цинодонтов вертлужная впадина была более открыта спереди и снизу. Головка бедренной кости отклонена от диафиза вверх и вперед, поэтому кость может направляться вперед под углом около 55° к сагиттальной плоскости и более эффективно двигаться по дорсовентральной дуге. Обе берцовые кости ориентированы вертикально.
Тазовый пояс видоизменен в соответствии со сменой направленности главных мышц, места прикрепления которых на бедренной кости также изменились. Лобково-седалищная пластина уменьшена и развернута немного назад, что отражает снижение размеров наружной лобково-седалищно-бедренной и аддукторной мышц и их важности для подъема тела на задней конечности. Поскольку лобковая и седалищная кости были развернуты назад, эти мускулы могли служить ее ретракторами. Область их прикрепления сместилась проксимальнее. Зона отхождения бедренно-хвостовой мышцы уменьшишь, и значение ее как ретрактора, несомненно, упало.
Ориентация и функция у цинодонтов седалищно-вертлужой мышцы, которая называется у млекопитающих внутренней запирательной (m. obturator internus), остаются такими же, как у пеликозавров, но зона ее прикрепления преобразуется в заметную новую структуру, большой вертлуг (trochanter major), находящийся сразу же под головкой бедренной кости на ее заднем крае.
Место отхождения подвздошно-бедренной мышцы у цинодонтов смещается вперед в связи с распространением в этом же направлении пластины подвздошной кости. Хотя эта зона лежит немного спереди вертлужной впадины, мышца действует как ретрактор, потому что область ее прикрепления находится сзади и вентральнее повернутой внутрь головки бедренной кости. Эта мышца, которая образует часть ягодичного комплекса у млекопитающих, служит одним из главных ретракторов бедра.
Внутренняя лобково-седалищно-бедренная мышца сохраняет свою роль ретрактора бедра. Место ее прикрепления перемещается с дорсальной на переднюю поверхность бедренной кости, в зону, известную у млекопитающих под названием малого вертлуга (trochanter minor). Эта бугорчатость находится в том же положении, что и внутренний вертлуг у рептилий, и можно считать их структурно гомологичными, хотя они и предназначены для прикрепления разных мускулов. У ранних цинодонтов наружная лобково-седалищно-бедренная мышца переместилась проксимально и назад, заняв межвертлужную яму, тогда как одноименная внутренняя мышца стала крепиться к малому вертлугу. Смещение зоны прикрепления внутреннего мускула вперед и вентральнее было необходимо вследствие изменения ориентации головки бедренной кости. Если бы у цинодонтов и млекопитающих сохранилось его примитивное дорсальное прикрепление, он бы вращал бедро и отклонял конечность латерально, а не двигал ее вперед.
У примитивных цинодонтов зона отхождения внутренней лобково-седалищно-бедренной мышцы осталась такой же, что и у пеликозавров, т. е. вдоль передней поверхности лобково-седалищной пластины. У продвинутых цинодонтов и млекопитающих она переместилась вперед и дорсальнее, на передний край подвздошной пластины, поясничные позвонки и прилегающую часть стенки тела. В этом положении мышца стала называться пояснично-подвздошной (m. iliopsoas).
Последовательные изменения, в результате которых тазовый пояс и задняя конечность продвинутых цинодонтов приобрели почти такую же форму, как у млекопитающих, наметились к началу триаса.
Сочленовные поверхности берцовых костей у Thrinaxodon видоизменены в соответствии с их более вертикальной ориентацией. Крупное функциональное изменение произошло в голеностопном суставе. Как у чешуйчатых и у примитивных архозавров, между голенью и стопой развилось простое сочленение с рычажной системой. Его схема у млекопитающих, аналогичная таковой у крокодилов, наблюдается уже у ранних цинодонтов. Таранная кость прочно присоединяется к большой берцовой, и основной сустав находится между таранной и пяточной костями, причем последняя интегрирована в стопу и развивает пяточный бугор. Как и у крокодилов, икроножные мышцы прикрепляются к пяточному бугру. Животные были стопоходящими. У среднетриасовых цинодонтов фаланговая формула редуцируется до 2 - 3 - 3 - 3 - 3. Однако у Thrinaxodon, судя по всему, все еще сохранялось по одной дополнительной фаланге дисковидной формы в III-м и IV-м пальцах.
Cynognathus
На следующей ступени эволюционного развития находится ранне- и среднетриасовй род цинодонтов Cynognathus. Его морда вытянута и сужена позади крупных клыков. Аддукторная камера расширена наружу и назад; окружающие ее кости шире и толще, чем у Thrinaxodon, что свидетельствует об очень сильном укусе. Камера не тянется так далеко вперед, значит, челюсти раскрывались шире, позволяя схватить более крупную добычу. Заклыковые зубы сжатые латерально, грубо пильчатые. Затылок широкий, возможно, для прикрепления очень массивной туловищной мускулатуры; латерально он сливается с концами очень высоких скуловых дуг.
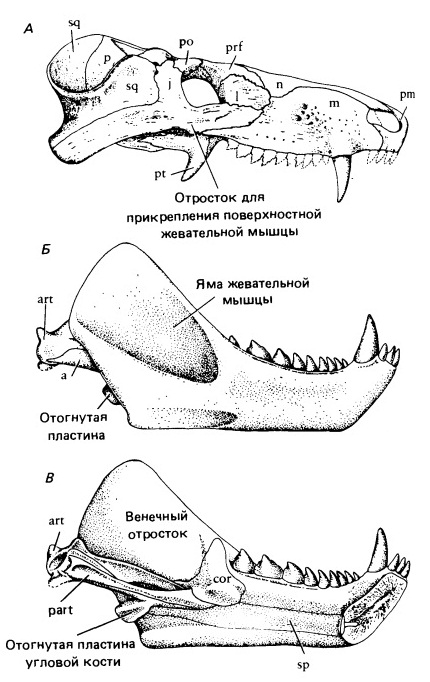
Прорисовки реконструированного черепа (а) и нижней челюсти (Б, В) циногната (Cynognathus crateronotus).
Зубная кость составляет значительно большую часть нижней челюсти, чем у более примитивных цинодонтов. Внешне она напоминает зубную кость крупных хищных млекопитающих с высоким венечным отростком. Яма жевательной мышцы глубокая, заходит далеко вперед. Нижний край зубной кости продолжается назад угловым отростком (processus angulare). У современных млекопитающих этот отросток служит местом прикрепления поверхностной жевательной мышцы (m. masseter superficialis), отходящей от латеральной поверхности передней части скуловой дуги (или крыловидной мышцы (m. pterygoideus) — например, у опоссума). У Cynognathus вниз от нее отходит жевательный отросток, связанный с началом этого мускула. Поверхностная жевательная мышца отделяется от переднего края развившейся ранее глубокой жевательной мышцы примитивных цинодонтов. Волокна поверхностной жевательной мышцы ориентированы постеровентрально почти под прямым углом к волокнам глубокой жевательной мышцы. Появлением поверхностной жевательной мышцы завершаются основные изменения, ведущие к развитию челюстной мускулатуры млекопитающих.
Хотя нижняя челюсть внешне выглядит, как у млекопитающих, более детальное изучение показывает, что зубная кость связана с целым рядом более мелких костей, объединенных в узкий стержень, вставленный в желобок на ее медиальной поверхности. Он состоит из сочленовной, предсочленовной, угловой и надугловой костей. Венечная кость остается в виде плоской пластины, прикрывающей передний конец этого стержня. Тип челюстного состава такой же, как у примитивных терапсид; он образован сочленовной и квадратной костями.