История изучения
Впервые малайский медведь был описан в 1821 г. С. Раффлзом по экземпляру из Бенкулена на западе Суматры в качестве Ursus malayanus. В 1825 г. Т. Хорсфилд при описании Helarctos euryspilus с Калимантана поместил его в отдельный род Helarctos. Это название образовано древнегреческими словами ήλιος (hēlios) — "солнечный" и αρκτος (arctos) — "медведь". Оно обусловлено наличием светлого пятна на груди животного, которое в местных преданиях часто фигурирует в качестве символа восходящего солнца.

Смонтированный скелет малайского медведя (Helarctos malayanus).
Helarctos anmamiticus, выделенный в 1901 г. П.М. Хеде из Аннама во Вьетнаме на основании незначительных отличий в наклоне морды и строении щечных зубов, в дальнейшем был признан младшим синонимом номинативного подвида Helarctos malayanus malayanus. Такая же судьба постигла таксон Ursus malayanus wardi, установленный в 1906 г. Р. Лидеккером по черепу малайского медведя, по мнению автора обладающему сходством с другим медвежьим черепом из Тибета; в дальнейшем выяснилось, что тибетский образец на самом деле принадлежит Ursus thibetanus.
Строение и функции
Размеры малайского медведя наименьшие в подсемействе Ursinae и среди современных медвежьих. Длина головы и туловища составляет 100-140 см, хвоста — 3-7 см, высота в холке — 50-70 см, масса — 20-70 кг. Самцы на 10-20 % больше самок линейно и значительно превосходят их по массе: в то время как первые весят порядка 30-70 кг, вторые — всего 20-40 кг.

Малайский медведь (Helarctos malayanus).
Голова большая и округлая, с короткой и широкой мордой. Ноздри лишены клапанов; фильтрум отсутствует. Ринариум доходит до верхней губы, его боковые края простираются назад и скрывают септу при взгляде сбоку. Губы обладают некоторой способностью вытягиваться, голая только верхняя губа. Длинный и тонкий язык может вытягиваться изо рта на 20-25 см. Строение губ и языка, вероятно, связано с использованием их для извлечения меда и личинок из пчелиных гнезд. Ушная раковина маленькая — 4-6 см в длину, закругленная, с ограниченной подвижностью, не усилена вертикальными хрящевыми гребнями. Высота раковины от супратрагуса до вершины меньше, чем ее наибольшая ширина.


Малайский медведь (Helarctos malayanus).
Телосложение коренастое. Шея длинная. Вокруг шеи кожа собирается в свободные складки, что позволяет при захвате ее челюстями вывернуться и укусить противника. Туловище относительно укороченное. Конечности сравнительно высокие, с большими лапами. Длина стопы — 18-21 см. Передние конечности более изогнутые, чем у бурого медведя (Ursus arctos), их лапы повернуты внутрь. Кисть с 2 карпальными подушечками. Пальцевые подушечки разъединены, имеются волосы в межпальцевых областях. Подошвы широкие, голые. Когти очень большие, серповидно изогнутые, острые, особенно крупные на передних конечностях.

Малайский медведь (Helarctos malayanus).
Волосяной покров мягкий и гладкий, в отличие от других медведей он короткий и относительно редкий. На лбу, за ушами и на плечах образуются завитки волос. Шерсть обычно блестящего угольно-черного цвета, иногда со светлым бурым или коричневатым оттенком на конечностях, однако в целом окраска может варьировать от сероватой до рыжей. У взрослых особей развит плотный черный подпушек, остевые волосы могут быть светлее. Морда впереди глаз серовато-желтая, коричневая или оранжевая, иногда серая или серебристая.

Малайский медведь (Helarctos malayanus) с высунутым языком; хорошо заметна мешковатость кожи в области шеи.
На груди расположено светлое пятно изменчивой формы: чаще всего U-образное, реже Y- или O-образное, либо состоящее из нескольких меньших пятен. Цвет его может быть коричневым, кремовым, белым, охристым или желтым. Иногда пятно полностью отсутствует; такой медведь рыжеватой окраски был обнаружен в юго-восточном Сабахе на Калимантане. Считается, что контрастные пятна на груди служат для устрашения соперников, когда медведь поднимается на задних конечностях. Края лап зачастую желто-коричневые или коричневые. Когти черные.

Примеры изменчивости формы и цвета пятна на груди малайского медведя (Helarctos malayanus).
Кондилобазальная длина черепа составляет 21,8-24,8 (в среднем 24,1) см у самцов и 18,5-22,8 (в среднем 21,2) см у самок, средняя наибольшая длина черепа — 26,5 см у самцов и 22,6 см у самок, средняя ширина верхней челюсти — 7,6 см у самцов и 6,4 см у самок, средняя межглазничная ширина — 7,1 см у самцов и 5,8 см у самок, средняя ширина в области сосцевидных отростков — 17 см у самцов и 14,3 см у самок.
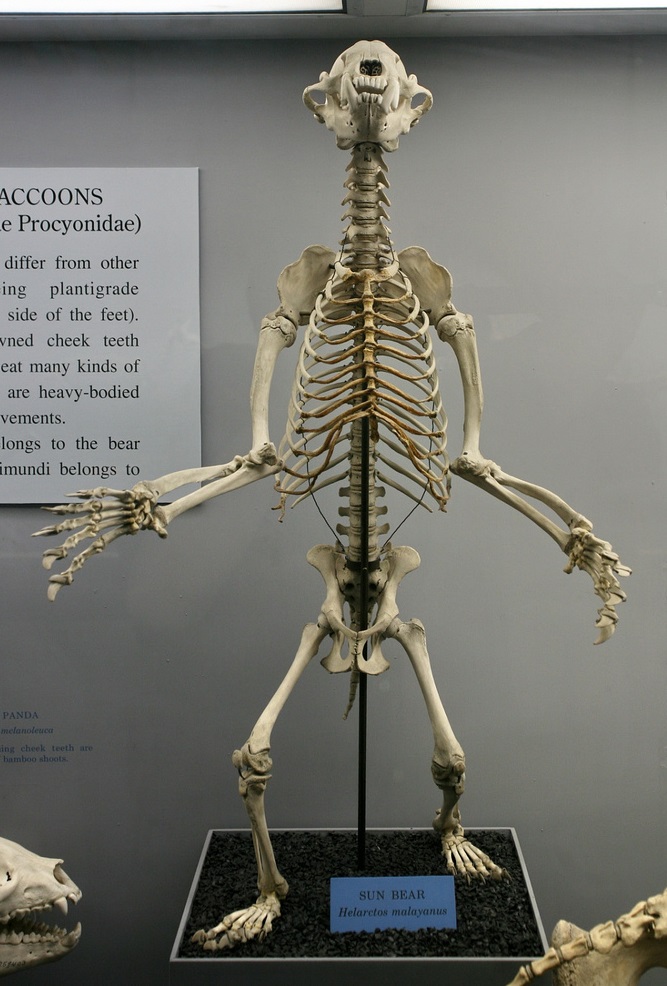
Смонтированный скелет малайского медведя (Helarctos malayanus).
Череп имеет дугообразный верхний профиль. Его лицевой отдел короткий и массивный. Длина носовых костей превышает их ширину в средней части примерно в 2,6 раза. Носовая апертура увеличена. Подглазничное отверстие лежит над М1. Скуловые дуги тонкие и широко расставленные. Заглазничные отростки небольшие. Мозговая коробка сильно расширенная, низкая. Степень цефализации наивысшая среди наземных хищных. Сагиттальный гребень у взрослых собей хорошо выражен. Костное небо узкое по отношению к черепу. Слуховые барабаны вздуты сильнее, чем у других Ursinae, и опускаются ниже поверхности основной затылочной кости. Заднее сонное отверстие отделено от заднего рваного отверстия и нередко смещено вперед от последнего. Сосцевидный отросток небольшой. Околозатылочный отросток крупный, тупой, выступающий вниз. Верхняя альвеолярная линия может проходить кзади и пересекать верхнюю часть суставной впадины и верхний край слухового прохода.
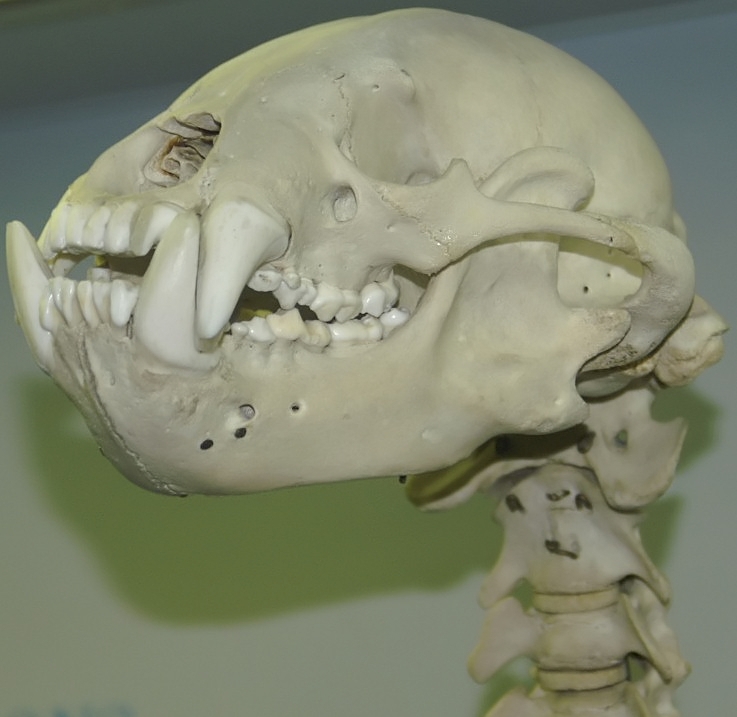
Череп с нижней челюстью малайского медведя (Helarctos malayanus).
Нижняя челюсть короткая, с высоким телом, высота которого возрастает кпереди, и длинным симфизом с почти вертикальным передним краем. Угловой отросток увеличен и приподнят. Сочленовный отросток лежит на уровне зубного ряда. Венечный отросток низкий, нижнечелюстная вырезка не развита. Нижний зубной ряд доходит до основания венечного отростка. При смыкании челюстей образуется небольшое заклыковое пространство. По сравнению с размерами тела, сила укуса очень велика.
Верхний и нижний зубные ряды короткие, что связано с укорочением лицевого отдела черепа и сокращением числа предкоренных зубов. Зубная формула: I3/i3, C1/c1, P2-4/p2-3, M2/m3 = 34-40. Клыки очень крупные для медведей такой величины. Они толстые и сильные, на их задне-наружном крае имеется острый вертикальный гребень. Ширина верхнего клыка у самцов составляет 17-18,5 мм. Щечные зубы небольшие, длина ряда нижних предкоренных короче таковой ряда коренных. Р1/р1 увеличены, при этом p1 по длине и площади коронки сопоставим с р4. Р2-3/р2-3, в противоположность, редуцированы в размерах и часто полностью утрачены, так что Р3/р3 или Р4/р4 примыкают непосредственно к Р1/р1 и упираются в задне-внутреннее основание клыков, так что их альвеолы непрерывны.
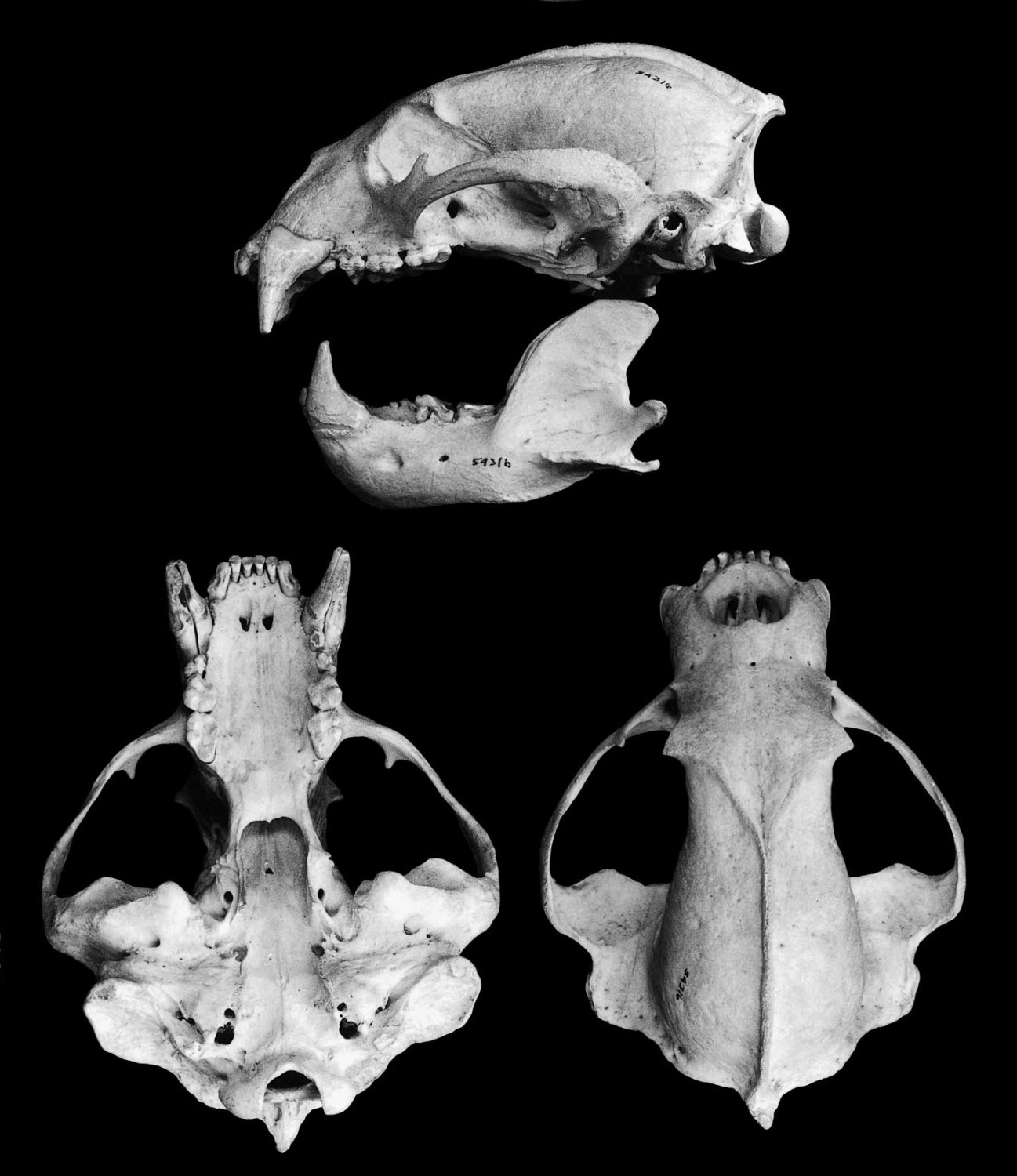
Череп (сбоку, снизу, сверху) и нижняя челюсть (сбоку) малайского медведя (Helarctos malayanus). Наибольшая длина черепа 23,2 см.
Верхний хищнический зуб Р4 уменьшен, с маленьким протоконом и хорошо заметным щечным цингулюмом. Коронка М1 почти квадратной формы, с развитым щечным цингулюмом. Имеется низкий язычный ряд зубцов с мезоконом; срединная долинка широкая, особенно в заднем отделе коронки. Парастиль и метастиль не развиты. М2 немногим длиннее, чем M1, с коротким талоном. Метакон одиночный, значительно меньше, чем паракон. Язычный цингулюм длинный, проходит почти вдоль всего края зуба, расширяя зубную коронку.
р4 простой, округлый, уступает по высоте паракониду m1. Нижний хищнический зуб m1 обычно с двойным метаконидом. Талонидный бассейн не развит. Энтоконид одиночный, ниже, чем гипоконид. Имеется задненаружный цингулид. Длина m1 современных особей — 15,1-17,6 мм, в среднем 16,7 мм. m2 лишь немногим короче m1, без сужения коронки в области смыкания тригонида и талонида. Метаконид выше, чем протоконид; параконид отсутствует. Талонидный бассейн широкий, энтоконид двойной. m3 округлой формы, он заметно крупнее, чем р4. Зубная эмаль с волнистыми полосами Гунтера - Шрегера.
Плечевой пояс по своим адаптациям к лазанию по деревьям сходен с таковым большой панды (Ailuropoda melanoleuca), включая степень напряжения, которое способно выдерживать плечо. Урсодезоксихолевая кислота желчи желчного пузыря составляет в среднем 8 % по сравнению с 17 % у белого медведя (Ursus maritimus), 19 % у бурого медведя (Ursus arctos) и 39 % у черного медведя (Ursus americanus).
Генетика
В кариотипе 74 хромосомы, NFa = 86. Кариотипически малайский медведь отличается от других урсин по наличию парацентрической инверсии в 14-й хромосоме и перицентрической инверсии в 18-й хромосоме, а также отсутствием негативно окрашенного стебля со светлым концевым сателлитом на 25-й хромосоме и наличием редуцированного короткого плеча у 34-й хромосомы. Два последних хромосомных признака общие с бурым медведем.
Родственные связи
Недостаток палеонтологического материала затрудняет выяснение происхождения и родственных связей рода Helarctos. Шлоссер (1899 г.) считал, что он имеет общие признаки с родом Agriotherium, но примитивнее его, и происходит от предка, общего c Ursavus и Agriotherium. Другие авторы (Пилгрим, 1914 г.; Зданский, 1924 г.; Гельбинг, 1932 г.) находили сходство в зубной морфологии между Indarctos и Helarctos, объясняя это проявлением атавистических черт у последнего рода. Отмечалось также, что зубная система Helarctos malayanus лишь немногим прогрессивнее, чем у Ursavus primaevus (Гидли, 1923 г.), в то время как у Ursavus ehrenbergi она уже настолько специализирована, что не позволяет рассматривать его в качестве предка малайского медведя (Тениус, 1947 г.).
По морфобиологическим особенностям Helarctos malayanus — примитивный, но при этом достаточно специализированный медведь, обитающий в тропическом лесу. Примечательно сходство между Helarctos malayanus и Мelursus ursinus в строении ринариума и верхней губы, хотя у малайского медведя нос и губы еще не столь подвижны, как у губача (Покок, 1918, 1941 гг.). Общие производные признаки позволяют предполагать филогенетическую близость таксонов, так что их иногда объединяют в один род (Ван Гельдер, 1977 г.). Родственные связи с губачом также поддерживаются случаями скрещивания этих видов в неволе с получением плодовитых гибридов.
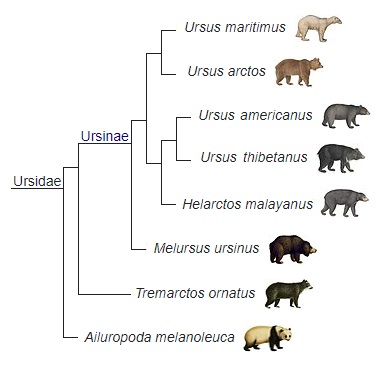
Кладограмма, отображающая родственные связи современных медведей на основе секвенирования их ядерных геномов.
Данные молекулярной биологии противоречивы. Генетическая дистанция Нея между Helarctos malayanus и Ursus arctos (0,026) оказалась меньше, чем таковая между Helarctos malayanus и Ursus thibetanus (0,037) и между Helarctos malayanus и Мelursus ursinus (0,050) (Голдман с соавт., 1989 г.). Одно исследование показывает базальное положение Мelursus ursinus в подсемействе Ursinae, в то время как Helarctos malayanus размещается ближе к кладе Ursus arctos / Ursus maritimus (Уайтс с соавт., 1999 г.). На основе анализа полных последовательностей митохондриальной ДНК (маркер цитохрома b) Helarctos malayanus сближают с кладой Ursus arctos / Ursus maritimus (Талбот и Шилдс, 1996 г.) или же считают его ближайшим родственником Ursus americanus (Чжан и Ридер, 1993 г.).
Согласно очередному анализу полных последовательностей митохондриальной ДНК, эволюционная линия Ursus arctos / maritimus разошлась с линией Ursus americanus / Ursus thibetanus / Helarctos malayanus около 6,72-5,54 млн. лет назад, а линия Helarctos malayanus, по-видимому, отделилась от линии Ursus americanus / Ursus thibetanus примерно 6,26-5,09 млн. лет назад (Юй с соавт., 2007 г.); по другим данным, последнее событие произошло 5,89-3,51 млн. лет назад (Краузе с соавт., 2008 г.). Анализ ядерной ДНК сближает Helarctos malayanus и Melursus ursinus как сестринские таксоны, комбинированный анализ ядерной и митохондриальной ДНК, напротив, удаляет его от Melursus ursinus, объединяя с Ursus americanus и Ursus arctos (Ли с соавт., 2004 г.). Секвенирование ядерных геномов современных медведей показало, что Melursus ursinus и Helarctos malayanus представляют собой примеры ранней радиации Ursinae и не входят в монофилетическую группу видов Ursus (Пейджес с соавт., 2008 г.).
Ископаемые находки
Жизнь во влажном тропическом климате, преимущественно на деревьях, является одной из причин почти полного отсутствия ископаемых остатков Helarctos malayanus. Их редкость в местонахождениях Индокитая и других континентальных частях видового ареала объясняют также конкурентными отношениями с Ursus thibetanus, который в плейстоцене вытеснял Helarctos malayanus. Ископаемые находки известны здесь лишь из среднего плейстоцена Вьетнама (пещеры Лантранг и Ланшон) и позднего плейстоцена провинции Хубэй на востоке Китая (местонахождение Хуанлундун). Более обычны они на островах Калимантан, Суматра и Ява (местонахождение Пунунг, 125-60 тыс. лет назад), где датируются поздним плейстоценом. Таким образом, в прошлом ареал малайского медведя простирался значительно дальше как на север, так и на юг.
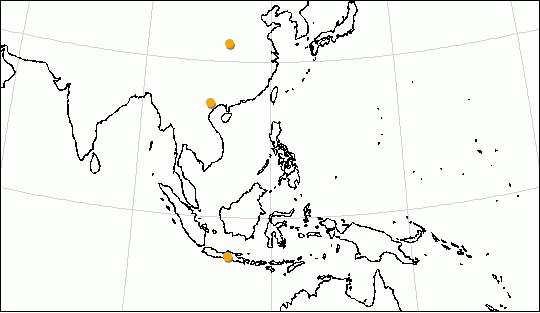
Некоторые места обнаружения ископаемых остатков малайского медведя (Helarctos malayanus).
Зданский (1927 г.) под названием Ursus cf. boeckhi описал фрагмент правой нижнечелюстной кости с m1, найденный в китайской провинции Шанси (местонахождение Лян-Чиа-Хo). Стратиграфическое положение находки неизвестно, предположительно — плиоцен. Тениус (1947 г.) обратил внимание, что тело нижнечелюстной кости заметно повышается к симфизу, как у Helarctos, и выделил китайскую находку в вид Ursus (Helarctos) sinomalayanus, считая его непосредственным предком современного малайского медведя. Единственный сохранившийся зуб m1 несколько крупнее, чем у последнего (длина 20,7 мм). Метаконид одиночный, расположен немного позади протоконида. Эрдбринк (1953 г.) предполагал, что метаконид был двойным, но его передний бугорок слился с ребром, отходящим от вершины протоконида. Гипоконид большой, энтоконид отчетливый, имеется энтоконулид. По середине талона проходит продольный желобок, не замкнутый сзади, как это часто имеет место у Helarctos malayanus. Морло и Кундрат (2001 г.) на основании развития эмалевых зубчиков впереди метаконида m1 отнесли этот образец к Ursus minimus.
Другой вид, Ursus praemalayanus, был описан Кенигсвальдом (1935 г.) по ископаемому правому М2, вероятно, купленным в китайской аптеке в Сингапуре или Гонконге. Автор описания отмечал сходство нового вида с Helarctos malayanus, а Эрдбринк (1953 г.) прямо пишет, что он не нашел между ними заметных различий. Для этой находки Барышников (2007 г.) условно принимает подвидовой статус. В прошлом к роду Helarctos также относили маленьких медведей из плиоцена и раннего плейстоцена Европы (Ursus boekhi, Ursus minimus) (Деперет, 1890 г.; Деперет и Льюэка, 1928 г.) и даже видели в Ursus boekhi предка современного Helarctos malayanus (Рыжевич, 1969 г.). В дальнейшем эта точка зрения была опровергнута (Дем, 1943 г.; Тениус, 1947 г.; Эрдбринк, 1953 г.; Барышников, 1992 г.).
Распространение
Еще недавно ареал малайского медведя охватывал восточную часть Бангладеш, северо-восток Индии (Ассам и прилегающие штаты), юг Китая (провинции Юньнань), Индокитайский полуостров (Мьянма, Таиланд, Лаос, Вьетнам, Камбоджа, Малайзия), Сингапур, Суматру (Индонезия) и Калимантан (Индонезия, Малайзия, Бруней). К настоящему времени в Сингапуре животное уже исчезло (по-видимому, вследствие обширных вырубок лесов в XIX-XX вв.), а в Бангладеш встречается только вдоль границ с Индией и Мьянмой. В Китае после многолетнего перерыва оно было отмечено в одном из приграничных уездов Юньнани. На территории прочих стран ареал в связи со сведением лесов значительно сократился; в особенности это касается Мьянмы, Таиланда и Вьетнама. Относительно хорошо дела обстоят в Лаосе, который благодаря обилию пересеченных ландшафтов, сохранению значительных облесенных площадей и сравнительно редкому человеческому населению является одним из последних оплотов малайского медведя.
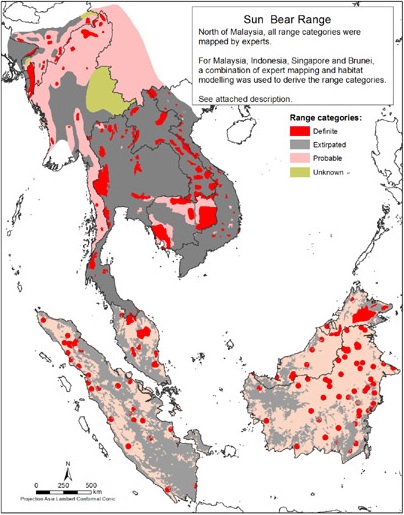
Современный ареал малайского медведя (Helarctos malayanus): красным цветом обозначены районы, где животное сохранилось; темно-серым — где уже исчезло; розовым — где его присутствие на сегодняшний день допускается, но не подтверждено; в закрашенных зеленым районах зверь отсутствует.
Численность популяции малайского медведя уменьшается к северу от Сундаленда, что особенно ярко проявлено на северных и западных окраинах ареала. Возможно, такая ситуация наблюдалась с доисторических времен и не связана напрямую с вмешательством человека. Плотность популяции варьирует от 4,3-5,9 особей на 100 кв. км в национальном парке Кхао Яй в центральной часте Таиланда до 26 особей на 100 кв. км в тропических лесах Харапан на юге Суматры.
Состав
Географическая изменчивость на материковой части ареала не выявлена. На острове Суматра медведи имеют более мелкие размеры, еще мельче звери с Калимантана, так что для последних был установлен подвид Helarctos malayanus euryspilus (Хорсфилд, 1825 г.; Покок, 1941 г.; Мейджаард, 2004 г.). Другие авторы считают вид монотипическим (Фицджеральд и Краусман, 2002 г.).
Исследование краниометрического материала показало, что медведи с Калимантана действительно характеризуются в среднем более мелкими размерами, чем материковые. Вместе с тем один из черепов оказался настолько крупным, что лишь незначительно уступает максимальным размерам, известным для черепов с континентальной части ареала, хотя его щечные зубы относительно невелики.
Полученные результаты позволяют признать для Helarctos malayanus 2 географические группы: в Индокитае и на острове Суматра, с одной стороны, и на острове Калимантан — с другой, которые различаются на подвидовом уровне. Молекулярный анализ медведей с Суматры и Калимантана показал существование у них 5 генетических линий, что объясняется вероятным обменом генов при контакте популяций во время последнего ледникового максимума, когда происходили осушение шельфовой зоны и образование сухопутных мостов.
Таким образом, на сегодняшний день в составе Helarctos malayanus имеется 1 вымерший и 2 современных подвида.
Helarctos malayanus praemalayanus (von Koenigswald, 1935) — ископаемый подвид, формально установленный по крупному правому М2 неизвестного геологического возраста (по-видимому, плейстоценового) из Юго-Восточной Азии (вероятно, Китая). Синоним: Ursus praemalayanus von Koenigswald, 1935.
Helarctos malayanus malayanus (Raffles, 1821) — современный подвид, занимающий весь ареал, за исключением Калимантана. Синонимы: Ursus malayanus Raffles, 1821; Helarctos annamiticus Heude, 1901; Ursus malayanus wardi Lydekker, 1906.
Кондилобазальная длина черепа составляет в среднем (самцы и самки) 23,2 см, по другим данным — 23,5 см для материковых медведей и 22,7 см для животных с Суматры. Щечные зубы увеличены (особенно М2), длина верхнего зубного ряда Р4-М2 (самцы и самки) равна в среднем 46,8 мм. Длина m1 — 15,4-18 мм, в среднем 16,7 мм.
Helarctos malayanus euryspilus (Horsfield, 1825) — современный подвид, населяющий Калимантан. Синоним: Helarctos euryspilus Horsfield, 1825.
Размеры меньше, чем у номинативного подвида. Кондилобазальная длина черепа составляет в среднем (самцы и самки) 20,1 см; по другим данным — 20,6 см. Щечные зубы небольшие, длина верхнего зубного ряда Р4-М2 (самцы и самки) равна в среднем 42,8 мм. Длина m1 — 15,1-17,3 мм, в среднем 16,4 мм.