История изучения
Первый описанный вид "американского гепарда" назвали Uncia inexpectata (Коп, 1899 г.). Находка представляла собой несколько разрознных зубов из Порт-Кеннеди-Кейв, Пенсильвания (ирвингтоний, 1-0,7 млн. лет назад, Лунделиус с соавт., 1987 г.). Затем были найдены фрагментарные остатки аналогичного возраста в Арканзасе (Браун, 1908 г.) и Мэриленде (Гидли и Гэзин, 1938 г.). Симпсон (1941 г.) исследовал все эти образцы и пришел к выводу, что они принадлежали одному и тому же новому виду, названному им Felis (Puma) inexpectata.
Сэведж (1960 г.) описал более обширный черепной и посткраниальный материал гепардоподобной кошки из Цита-Каньон (бланкий, около 2,5 млн. лет назад). Он определил этот материал в качестве нового вида Felis (Puma) studeri, так как полагал, что описанный Сэведжем таксон является nomen dubium, основанным на недостаточном материале. Третий вид, Felis trumani, был установлен Орром (1969 г.) на основании находки полного черепа в Крипт-Кейв, Невада, датированного концом плейстоцена (19 750 ± 650 тыс. лет назад). Хотя Орр не сравнивал этот череп с материалом Felis studeri, он с высокой степенью вероятности демонстрирует много общих черт с более ранним таксоном.
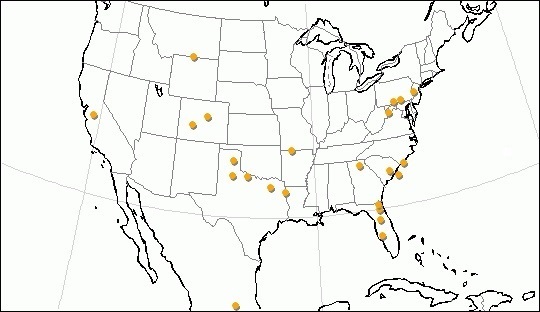
Основные местонахождения ископаемых остатков Miracinonyx.
Куртен (1976 г.) рассмотрел ископаемых пум Северной Америки, включая весь материал, ранее приписываемый Felis inexpectata и Felis studeri, и определил, что существовал только один вид, предшествующий ранчолабреанийским отложениям. Он отнес весь материал к виду Felis (Puma) inexpectata. Последующий материал Felis trumani был обнаружен в Нейчерал-Трэп-Кейв, Вайоминг (ранчолабреаний, 13-11 тыс. лет назад; Мартин с соавт., 1977 г.).
Таксон Miracinonyx был выделен в 1979 г. Адамсом в качестве подрода рода Acinonyx. Его название можно перевести как "чудесный, удивительный гепард". Адамс (1979 г.), а затем Куртен и Андерсон (1980 г.) указывали на существование в Северной Америке двух видов гепардоподобных кошек — Acinonyx trumani и Acinonyx studeri. В своей статье последние авторы заявили, что фрагментарные остатки из позднего ирвингтония, ранее идентифицируемые в качестве Felis inexpectata, являются смешанным материалом, и реклассифицировали Acinonyx studeri в Acinonyx trumani. Кроме того, материал из Гамильтон-Кейв в Виргинии подтверждает существование только двух видов, и более ранние формы, подобные материалу Копа, следует именовать Acinonyx inexpectatus, а не Acinonyx studeri.

Череп и нижняя челюсть мирациноникса (Miracinonyx trumani).
Состав
Miracinonyx inexpectatus (Cope, 1895) Van Valkenburgh, Grady et Kurten, 1990 — вид, известный из раннего - позднего плейстоцена (конец бланкского - конец ранчолабрейского века) США (штаты Калифорния, Техас, Арканзас, Вайоминг, Колорадо, Пенсильвания, Мерилэнд, Джорджия, Западная Виргиния, Виргиния, Южная Каролина, Флорида) и Мексики (штаты Гуанахуато, Южная Нижняя Калифорния). Синонимы: Crocuta inexpectata Cope, 1895; Uncia inexpectata Cope, 1899; Felis inexpectatus Hay, 1902; Felis longicrus Brown, 1908; Smilodontopsis mooreheadi Hay, 1921; Felis inexpectata Simpson, 1941; Felis studeri Savage, 1960; Acinonyx (Miracinonyx) studeri Adams, 1979; Acinonyx studeri Kurten et Anderson, 1980; Miracinonyx studeri Alroy, 2002.

Прорисовка реконструированного скелета мирациноникса (Miracinonyx inexpectatus). Автор — М. Антон.
Miracinonyx trumani (Orr, 1969) Van Valkenburgh, Grady et Kurten, 1990 — вид, известный из позднего плейстоцена (середина - конец ранчолабрейского века) США (штаты Колорадо, Аризона, Вайоминг, Южная Каролина). Синонимы: Felis trumani Orr, 1969; Acinonyx (Miracinonyx) trumani Adams, 1979; Acinonyx trumani Kurten et Anderson, 1980; Miracinonyx trumani Van Valkenburgh, Grady et Kurten, 1990.
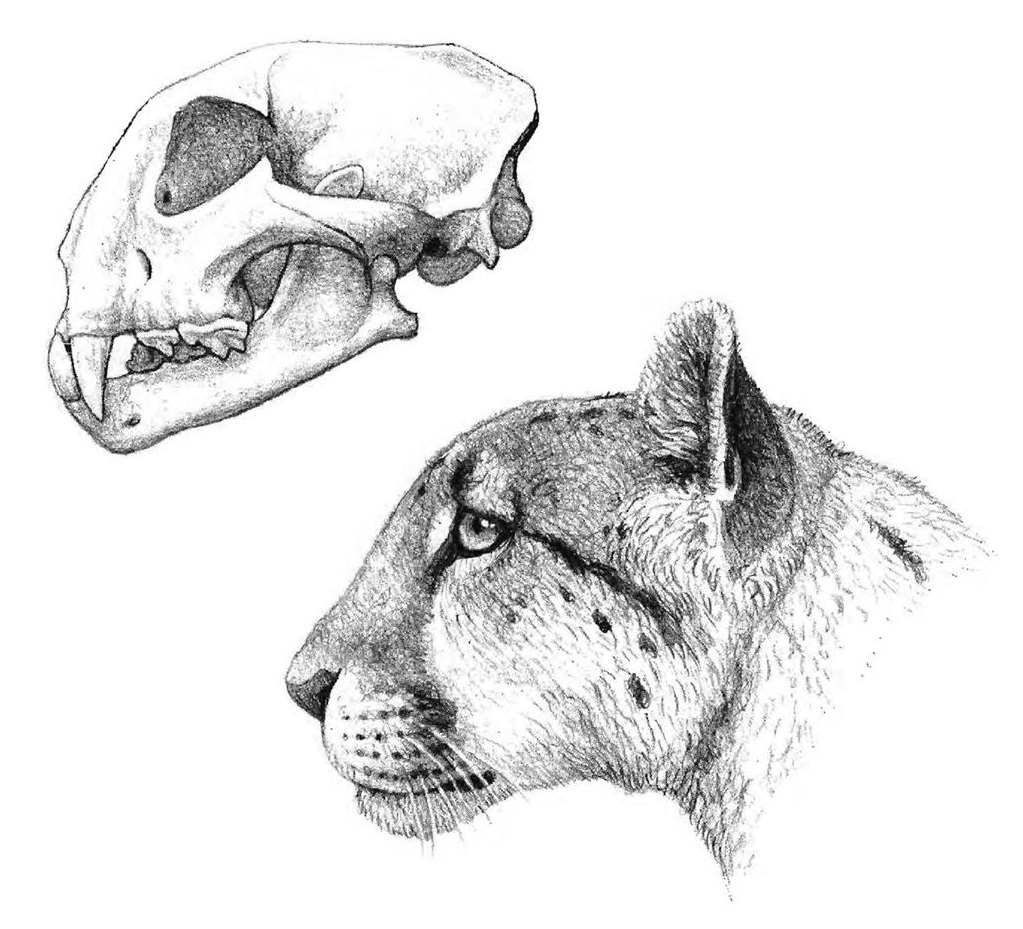
Прорисовка черепа с нижней челюстью и портретная реконструкция мирациноникса (Miracinonyx trumani). Автор — М. Антон.
Остатки, определенные лишь до рода и обозначенные Miracinonyx spp., известны из раннего / среднего плейстоцена США (штаты Калифорния и Флорида).
Строение и функции
Длина головы и туловища мирацинониксов составляла 130-170 см, длина хвоста достигала 95 см, высота в холке — порядка 85 см. Вес варьировал в пределах 55-95 кг. По своим размерам Miracinonyx inexpectatus схож с плио-плейстоценовым европейским Acinonyx pardinensis, тогда как более поздний и мелкий Miracinonyx trumani соразмерен с ныне живущим Acinonyx jubatus. Оба вида мирацинониксов демонстрируют общие морфологические черты и телесные пропорции гепардов.
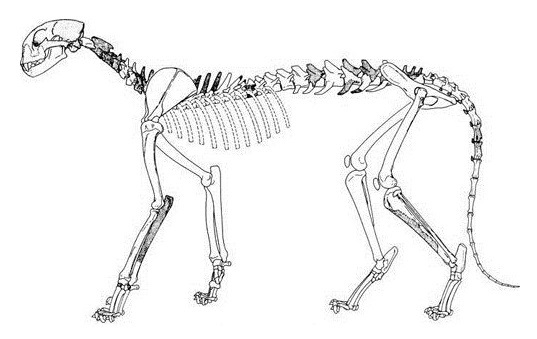
Прорисовка скелета Miracinonyx inexpectatus, реконструированного на основе образца USNM 401092.
Череп укороченный, с широкой лобной областью и куполообразный, как у Acinonyx jubatus. Подобно последнему и в отличие от других кошек, имеется редуцированное подглазничное отверстие, что у Acinonyx jubatus связано с уменьшением вибриссов. Передняя часть скуловой дуги укорочена. Скуловой отросток верхнечелюстной кости, который у Puma concolor заметен под глазницей, уменьшен. Глазницы расположены далеко друг от друга и высоко на лицевой части черепа. Заглазничные отростки короткие. Слуховая кость удлинена и спереди сглажена. Последняя особенность отличает представителей рода Miracinonyx от гепардов Старого Света.
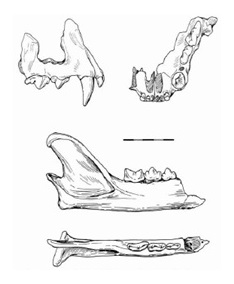
Прорисовки части лицевого отдела черепа Miracinonyx inexpectatus снаружи (вверху слева) и снизу (вверху справа) и правой ветви нижней челюсти снаружи и сверху. Длина масштабной линейки — 5 см.
Отверстия ноздрей и внутренние носовые проходы расширены, как и у Acinonyx jubatus — по-видимому для увеличения объема вдыхаемого воздуха, необходимого для бега. Резцовое отверстие заметно в полостях верхней челюсти и подобно формой и размером таковому у Puma concolor. Фрагменты расширенных предчелюстных и верхнечелюстных костей предполагают, что носовой проход был как минимум таким же или гораздо более обширным, чем у соразмерной Puma concolor, но меньшим, нежели у Acinonyx jubatus.
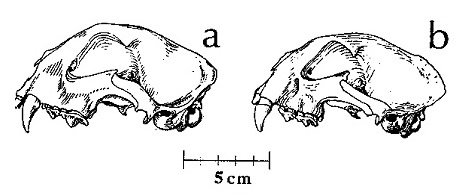
Прорисовки черепов: A — Acinonyx jubatus; B — Miracinonyx trumani.
В нижней челюсти имеется короткая заклыковая диастема. Нижнечелюстной угол уменьшен относительно такового у Puma concolor и Acinonyx jubatus. Суставная поверхность мыщелка закруглена верхне-наружно больше, чем у Acinonyx jubatus, но не до такой степени, как у Puma concolor.
Тело подъязычной кости приблизительно 2,7 см длиной, симметричное с обеих сторон, относительно массивнее, чем другие кости подъязычного аппарата и имеет суставные поверхности на каждом конце. Две другие кости трудно идентифицировать. Каждая подобна в длине подъязычной кости, но тоньше и сужается к одному концу; они могут являться дальними или средними члениками. Если это дальний членик ветви, то Miracinonyx, как и современные Puma concolor и Acinonyx jubatus, не могли издавать рычание. Кроме того, прочие костные элементы приблизительно наполовину меньше и незначительно расширены в каждом конце.
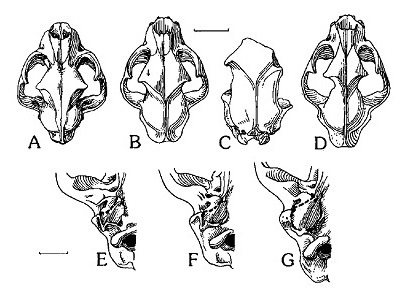
Прорисовки черепов сверху и слуховой области снизу: A, Е — Acinonyx jubatus; B, F — Miracinonyx trumani; C — Miracinonyx inexpectatus; D, G — Puma concolor. Длина масштабных линеек — 5 и 2 см.
I3 увеличен относительно I2 в большей степени, чем у Acinonyx jubatus. Как и у Puma concolor, I3 имеет два небольших бугорка, один дальний и один среднеязычный, что увеличивает первичный центральный бугор. Верхний клык демонстрирует тенденцию к уменьшению, не зазубрен и отделен от I3 короткой диастемой. Клык меньше относительно щечных зубов, чем у Puma concolor, но не столь редуцирован, как у Acinonyx jubatus. P2 небольшой, однокорневой. P3 — относительно простой зуб с двумя бугорками, основным и меньшим последующим. У видов рода Miracinonyx, так же как у Puma и Acinonyx, присутствует задне расположенный цингулюм, но у Acinonyx он более острый. У представителей рода Miracinonyx наружный край смещен вперед, этот край несет небольшой бугорок — тот же самый бугорок подвержен индивидуальной изменчивости у Puma concolor. У P4 эктопарастиль отсутствует, а протокон редуцирован относительно такового у Puma concolor, но не настолько, как у Acinonyx jubatus. Однако, в отличие от последнего, протокон направлен несколько вперед, а не назад. Развитие эктопарастиля является индивидуально изменчивым и у Puma concolor.
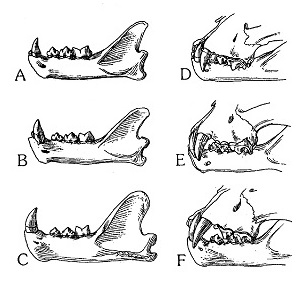
Прорисовки нижних челюстей и характера смыкания зубов: A, D — Acinonyx jubatus; B, E — Miracinonyx trumani; C, F — Puma concolor.
Нижние зубы относительно более узкие, чем у Puma concolor, но не настолько, как у Acinonyx jubatus. Вполне возможно, что у видов рода Miracinonyx присутствовала некоторая внутривидовая изменчивость в форме нижних предкоренных. Например, имеются бланкские и ирвингтонские образцы с относительно широкими, или, наоборот, сравнительно узкими предкоренными. P3 у представителей Miracinonyx имеет хорошо заметный цингулюм. Эти две черты строения отсутствуют либо слабо развиты у Puma concolor и более выражены у Acinonyx jubatus. Оба предкоренных имеют более высокие коронки относительно их продольного размера, чем у Puma concolor. Подобно многим кошкам, р4 и m1 слегка перекрываются.
Покок (1916 г.) отметил, что Acinonyx jubatus уникален среди ныне живущих кошек специфическом набором черт зубного строения, выражающемся в чрезвычайном перекрытии зубных рядов, действующих, подобно ножницам. Эта специализация зубного аппарата включает уменьшение верхних и нижних клыков, сокращение верхней и нижней заклыковых диастем, сужение предкоренных, отсутствие протокона. В этом отношении представители рода Miracinonyx почти идентичны Acinonyx jubatus и отличаются от Puma concolor, имеющей значительную заклыковую диастему.
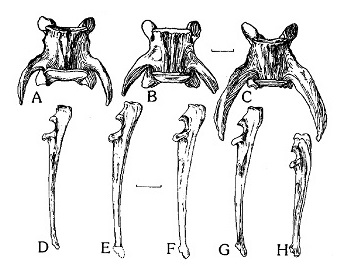
Прорисовки VII-х поясничных позвонков и локтевых костей: A, D — Acinonyx jubatus; B, G — Miracinonyx trumani; C, H — Puma concolor; E — Acinonyx pardinensis; F — Miracinonyx inexpectatus. Длина масштабных линеек — 2 и 5 см.
Расчетная длина предхвоствого позвоночного столба Miracinonyx inexpectatus, включая крестец, составляет 106 см. Поясничный отдел длиннее относительно грудного отдела, чем у Puma concolor. Нужно учитывать, что соотношение общей длины поясничной области к грудной составляет приблизительно 0,78 у Puma concolor, 0,90-1,00 у представителей рода Miracinonyx и у современного Acinonyx jubatus. У последнего длинная поясница функционирует при сгибании спины во время резкого броска, увеличивая длину шага и скорость. Невральные дуги позвонков поясничной области расширены передне-задне, как у Puma concolor, в отличие от узких дуг Acinonyx jubatus. В дополнение к разделению сравнительно длинной поясничной области, VII-й позвонок Acinonyx jubatus и видов рода Miracinonyx имеет специфические поперечные фасетки. Оценочная длина хвоста составляет 85-90 см, или 74-79 % длины предхвостового позвоночного столба. Это преполагает относительную длину хвоста, скорее подобную гепардам (79 %) чем пумам (90 %).
В ископаемом материале представлены, по меньшей мере, 17 ребер, включая I-е. Отдельные фрагменты демонстрируют некоторые патологии. Наличествуют 4 элемента грудины.
Лопатка несколько длиннее, чем у Puma concolor. Как и у Acinonyx jubatus, у Miracinonyx имеется небольшое углубление вдоль боковой границы сутавной впадщины для надостной мышцы. Однако, в отличие от Acinonyx jubatus, ямка крепления большой круглой мышцы выглядит более узкой. Акромион, продолговатый отросток в верхней части лопатки, не выступает над сутавной ямкой, как у Puma concolor.
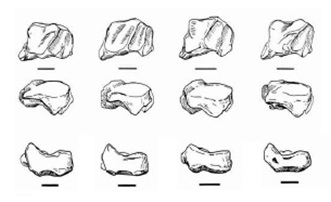
Прорисовки лопаток дальне и спереди, а также левых ладьевидных костей (внизу), слева направо: Miracinonyx inexpectatus, Miracinonyx trumani, Acinonyx jubatus, Рuma concolor. Длина масштабной линейки — 10 см.
Плечевая кость примерно на 20-25 % длиннее, чем у современных Puma concolor и Acinonyx jubatus и подобна таковой вымершего Acinonyx pardinensis. По форме она занимает промежуточное положение между плечевыми костями Puma concolor и Acinonyx jubatus. Например, подобно Puma concolor, ближняя часть костного стержня сжата срединно-наружно, дельтопекторальный гребень небольшой, наружный гребень для крепления плечелучевой мышцы уменьшен и срединный надмыщелок небольшой. Ближняя эпифизарная линия очевидна. У Miracinonyx trumani передне-срединная граница головки суставного конца кости больше ориентирована по аналогии с Miracinonyx inexpectatus, чем с Puma concolor. Однако, ее дальний конец менее сжат срединно-наружно и энтепикондилярное отверстие больше, чем у Acinonyx pardinensis и Acinonyx jubatus.
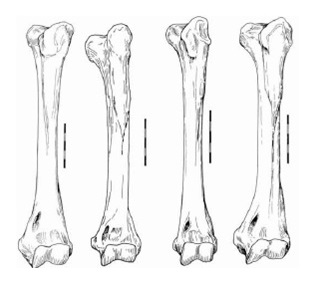
Прорисовки левых плечевых костей спереди, слева направо: Miracinonyx inexpectatus, Miracinonyx trumani, Acinonyx jubatus, Рuma concolor. Длина масштабной линейки — 5 см.
Лучевая кость составляет почти 90 % длины плечевой кости, как и у Puma concolor. Соотношение длин лучевой и плечевой костей напоминает таковое у современного Acinonyx jubatus. Ближняя суставная поверхность ориентирована под небольшим углом к стержню в дальней проекции, как у Puma concolor и Acinonyx pardinensis. Тело кости тонкое, как у Acinonyx jubatus. Имеется ближне-эпифизарная линия. Бугор двуглавой мышцы плеча соразмерен с таковым у Puma concolor и Acinonyx pardinensis, где он составляет 10 % полной длины тела кости в отличие от 5 % у Acinonyx jubatus. Суставная фасетка для соединения с локтевой костью имеет эллиптическую форму. Ее длинная ось ориентирована перпендикулярно к стержню, как у Puma concolor, а не так, как у гепардов Старого Света.
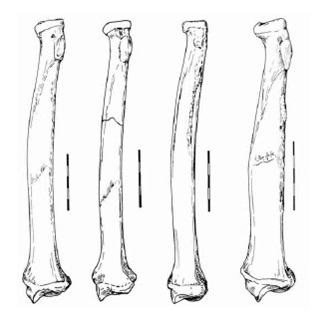
Прорисовки правых лучевых костей ближне, слева направо: Miracinonyx inexpectatus, Miracinonyx trumani, Acinonyx jubatus, Рuma concolor. Длина масштабной линейки — 5 см.
Локтевая кость более массивная, чем у Acinonyx jubatus. Средний гребень, который служит для прикрепления квадратного пронатора, является крупным и шиловидный отросток хорошо развит. Сочленовная фасетка для соединения с лучевой костью также больше. Локтевая кость, которая морфологически сходна с таковой у псовых, заметно отличается от кости Puma concolor своей удлиненностью и отчетливо вогнутым задним краем.
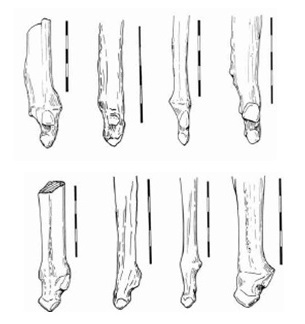
Прорисовки локтевых костей дальне радиально (вверху) и срединно (внизу), слева направо: Miracinonyx inexpectatus, Miracinonyx trumani, Acinonyx jubatus, Рuma concolor. Длина масштабной линейки — 10 см.
Запястные суставы по сравнению с таковыми Puma concolor являются на удивление небольшими. Это предполагает, что у бегающих кошек рода Miracinonyx запястье относительно общих размеров тела было относительно тонким, подобно тому, что наблюдается у Acinonyx jubatus. Дальняя суставная поверхность трапециевидной кости разделена гребнем у Puma concolor и обоих видов Miracinonyx, тогда как у Acinonyx jubatus является гладко вогнутой. С передней стороны ладьевидная кость срединно сжата в большей степени, чем у других кошек. Крючковидная кость запястья в передней части слегка клиновидной формы, будучи более широкой дальне, чем ближне. V-я плюсневая кость отличается от таковой Puma concolor ее относительно тонкими ближним и дальним концами.

Слепок костей левой кисти мирациноникса (Miracinonyx trumani).
Представители рода Miracinonyx отличаются от Puma concolor наличием относительно большей поверхности на II-й пястной кости, которая сильнее изогнута. Ближняя суставная поверхность трапецевидной кости у Miracinonyx и Puma является вогнутой, тогда как у Acinonyx jubatus она плоская или немного выпуклая. Гороховидная кость несколько удлинена и больше похожа на таковую Acinonyx jubatus, чем Puma concolor. Однако суставная поверхность клиновидной кости имеет яйцевидную форму, как у Puma concolor, а не треугольную, как у Acinonyx jubatus. Гороховидная кость имеет углубление на срединной поверхности головки, которой нет ни у Puma concolor, ни у Acinonyx jubatus. Все пястные кости на 20-25 % длиннее и имеют менее выраженные площадки для крепления мышц (например, разгибателей запястья), чем у Puma concolor.
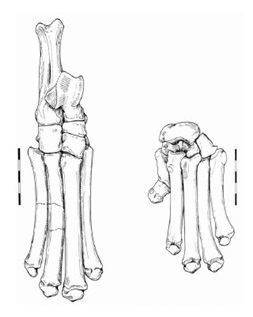
Прорисовки неполного скелета правой стопы (слева) и левой кисти (справа) Miracinonyx inexpectatus. Длина масштабной линейки — 5 см.
Пястные кости Miracinonyx c относительно узкими ближним и дальним концами. В этом отношении они близки к пястным костям Acinonyx jubatus, сильно сжатым с боков. I-я пястная кость рудиментарна, как у всех кошек. II-я пястная кость с небольшим ближним углублением для лучевой артерии. Ближний конец III-й пястной кости выступает над IV-й костью больше, чем у Acinonyx jubatus и меньше, чем у Puma concolor. Эта черта также характерна для ближнего конца IV-й пястной кости, а у представителей рода Miracinonyx там имеются отметины на местах прикрепления III-малоберцовой и короткой малоберцовой мышц. Также виды Miracinonyx имеют общую с Puma concolor и Herpailurus yagourundi форму ближнего конца IV-й пястной кости с заметным при виде спереди зубцом. Херрингтон (1986 г.) использовал эту общую особенность, объединяя этих кошек как монофилетичную группу в пределах Felidae.
Длина бедренной кости превышает длину бедра Puma concolor на 20-25 %. Ее тело прямое при виде сверху, как у Puma concolor. Тело кости наклонено вперед в большей степени, чем у Acinonyx pardinensis и Acinonyx jubatus. Суставная поверхность дальней головки кости более ровная, чем у Acinonyx jubatus. Коленная чашечка имеет слабовыпуклую поверхность для соединения с бедром, как у Puma concolor.
Голень примерно на 25-30 % длиннее, чем у Puma concolor. Отношение голени к длине бедра составляет приблизительно 0,95 у Miracinonyx inexpectatus и Miracinonyx trumani, в отличие от 0,90 у Puma concolor и 1,0 у гепардов Старого Света. Суставная поверхность квадратной кости относительно больше, чем у Acinonyx jubatus, где она сжата с боков. Кроме того, гребень голени прямой при виде сбоку, как у Puma concolor, вместо ближне вогнутого, как у Acinonyx. Углубления для длинного разгибателя пальцев и задней большеберцовой мышцы по степени развития сходны с таковыми у Puma и более глубокие, чем у Acinonyx. В дальней проекции суставная поверхность иззубрена вдоль заднего края, как у Puma concolor; у гепардов Старого Света этот край является плоским.
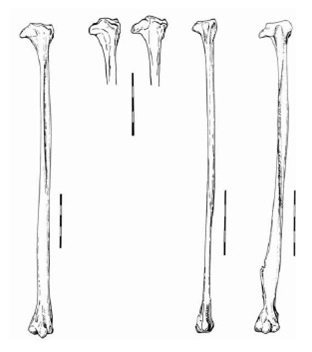
Прорисовки малых берцовых костей снаружи, слева направо: Miracinonyx inexpectatus, Miracinonyx trumani, Acinonyx jubatus, Рuma concolor. Длина масштабной линейки — 5 см.
В сравнении с Puma concolor тело малой берцовой кости более бугристо с наружной стороны. У головки таранной кости имеется ладьевидная суставная поверхность, которая пролегает срединно, как у Puma concolor, но не как у Acinonyx. Средняя пяточная поверхность округлая, с небольшим ближним расширением, как у Puma concolor. Боковая пяточная суставная поверхность вогнута меньше, чем у Acinonyx jubatus и подобна таковой у Puma concolor. Относительная длина шейки таранной кости, измеренной вдоль ее задне-срединной поверхности, не настолько большая, как у Acinonyx jubatus, но больше, чем у Puma concolor. В сочленении таранная кость почти соприкасается с задним краем пяточной кости, как у Puma concolor. У Acinonyx таранная кость не проходит как далеко, учитывая обширное пяточно-ладьевидное сочленение в суставе.
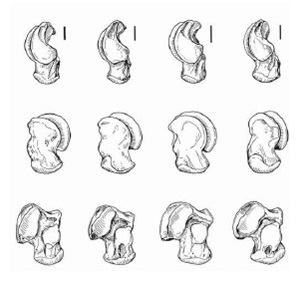
Прорисовки левых таранных костей снаружи (вверху), изнутри (середина) и снизу (внизу), слева направо: Miracinonyx inexpectatus, Miracinonyx trumani, Acinonyx jubatus, Рuma concolor. Длина масштабной линейки — 10 см.
Пяточная кость подобна таковой Puma concolor, а не Acinonyx jubatus. Поддерживающая структура относительно крупная и имеет развитый задне-срединный гребень, ограничивающий углубление для сгибающего мускула. Дальне костный стержень немного выгнут наружу, поддерживая сухожилие длинной малоберцовой мышцы. Имеется небольшая дальняя поверхность для контакта с ладьевидной костью; поверхность ориентирована вертикальнее, чем у Acinonyx. Вымерший Acinonyx pardinensis имеет необычно большую ладьевидную поверхность полулунной формы, редуцированную у современного Acinonyx jubatus. У обоих видов Miracinonyx срединная и наружная поверхности суставной фасетки отделены гребнем.
У ладьевидной кости умеренно развита бугристость, расширяющая ее ближне в большей степени, чем у современного Acinonyx jubatus. В этом отношении она аналогична Puma concolor и Acinonyx pardinensis. Виды Нового Света также имеют более глубокую и вогнутую суставную поверхность головки таранной кости. Однако ладьевидная кость Puma concolor отличается от таковой и Miracinonyx, и Acinonyx относительно большей ближне-дальней сжатостью. В отличие от Acinonyx, у видов рода Miracinonyx и у Puma concolor есть небольшая, вертикально ориентированная поверхность для сочленения с пяточной костью. У Acinonyx эта поверхность относительно больше и расположена более горизонтально (особенно у Acinonyx pardinensis) и имеет, таким образом, большое значение в распределении нагрузки между двумя костями предплюсны.
Ближняя поверхность кубовидной кости слабовыпуклая, прямоугольной формы и имеет маленькую фасетку для сочленения с ладьевидной костью. Эта особенность также характерна для Puma, но не для Acinonyx. Однако Acinonyx pardinensis демонстрирует большое количество вогнутых поверхностей ладьевидной кости. Перифериферически кубовидная кость Miracinonyx аналогична таковым Puma и Acinonyx в отношении наличия двух фасеток: большей для IV-й пястной кости и гораздо меньшей для V-й пястной кости.
Наружная часть клиновидной кости более широка ближне, чем дальне, придавая предплюсневой кости несколько трапециевидную форму при виде сверху. Углубление для сухожилия длинной малоберцовой мышцы является рельефным и относительно большим, чем у Acinonyx jubatus. Средняя и наружная части клиновидной кости более сходны с таковыми у Puma concolor, чем у Acinonyx jubatus; ближне и дальне они более овальные, чем у Acinonyx jubatus.
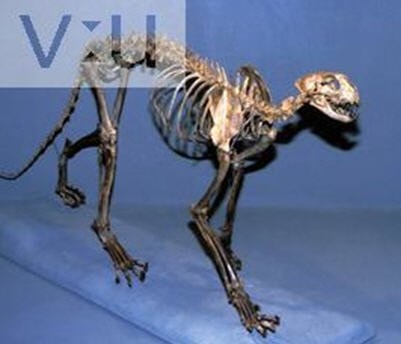
Реконструированный скелет Miracinonyx.
Плюсневые кости на 20-27 % длиннее, чем у Puma concolor; различия в длине наиболее выражены для II-й и V-й костей. В результате Miracinonyx походит на Acinonyx pardinensis наличием боковых плюсневых костей (II-й и V-й), которые больше, чем срединные кости (III-я и IV-я). У Puma concolor, обоих видов Miracinonyx и Acinonyx jubatus длина боковых элементов составляет приблизительно 88 % длины срединных. Однако у Puma concolor плюсневые кости имеют тенденцию к большему дальнему расширению по отношению к средней ширине костного стержня, чем таковые Miracinonyx и Acinonyx.
Фаланги представлены ближними, средними и когтевыми элементами. Средние и когтевые фаланги указывают на то, что когти у Miracinonyx inexpectatus и Miracinonyx trumani могли полностью втягиваться. Средние фаланги имеют вогнутость вдоль одной стороны тела кости, а для когтевых фаланг характерно увеличение нависающего отростка для присоединения мышцы глубокого сгибателя пальцев (разгибателя когтей). В отличие от этого, когтевые фаланги современного Acinonyx pardinensis могут сгибаться только частично, и точки прикрепления сгибающей мышцы на них уменьшены по сравнению с таковыми у других кошек. Одна из сохранившихся когтевых фаланг Miracinonyx inexpectatus намного больше, чем другие и, вероятно, представляет собой когтевую фалангу I-го пальца кисти.
По сравнению с Acinonyx jubatus, виды рода Miracinonyx имеют сильнее развитую заднюю конечность с большими супинаторными возможностями. Малая берцовая кость у Miracinonyx является более мощной, не соединенной с голенью, и размеры ложбинок для прикрепления мышц свидетельствуют о том, что представители рода Miracinonyx были более мускулистыми, чем гепарды. У Acinonyx jubatus большая и малая берцовые кости срослись, ограничивая, таким образом, двигательный момент в лодыжке. Подвижная малая берцовая кость позволяет задней конечности лучше приспосабливаться к неровностям почвы при пересеченном рельефе и играет важную роль в преодолении подъема.