Млекопитающеобразные — Mammaliaformes
Домен Ядерные (Eukaryota)
Царство Животные (Animalia)
Подцарство Настоящие многоклеточные (Eumetazoa)
Клада Двусторонне-симметричные (Bilateria)
Клада Вторичноротые (Deuterostomia)
Тип Хордовые (Chordata)
Клада Черепные (Craniata)
Подтип Позвоночные (Vertebrata)
Инфратип Челюстноротые (Gnathostomata)
Клада Эвгнатостоматы (Eugnathostomata)
Клада Конечноротые (Teleostomi)
Клада Эвтелеостоми (Euteleostomi)
Клада Лопастеперые (Sarcopterygii)
Клада Хоановые (Choanata)
Клада Четвероногообразные (Tetrapodomorpha)
Надкласс Четвероногие (Tetrapoda)
Клада Амниоты (Amniota)
Клада Синапсидообразные (Synapsidomorpha)
Клада Млекопитающеобразные (Mammaliaformes)
Млекопитающеобразные (Mammaliaformes Rowe, 1988) — клада, включающая млекопитающих и их ближайших ископаемых родственников. Известны начиная с позднего триаса.
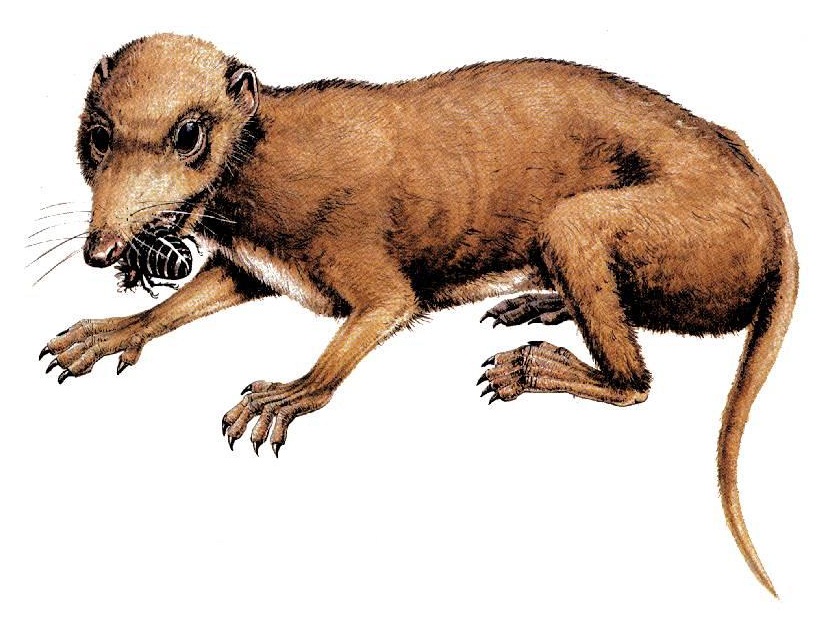
Реконструкция внешнего облика мегазостродона (Megazostrodon rudnerae).
Т. Роу полагал, что при отнесении организмов к млекопитающим (Mammalia) используются дополнительные критерии, из-за чего эта группа приобрела неоправданно широкий объем. Введенное им определение Mammaliaformes обозначило кладу, включающую потомков последнего общего предка морганукодонов (Morganucodonta) и млекопитающих. Собственно млекопитающие (Mammalia) в рамках данной концепции рассматриваются как краун-группа, включающая потомков последнего общего предка однопроходных, сумчатых и плацентарных. Более обширная клада Mammaliamorpha была предложена Роу для обозначения потомков последнего общего предка тритилодонтовых (Tritylodontidae) и краун-группы млекопитающих.
Млекопитающеобразные сформировалась в результате радиации пробайногнатов (Probainognathia) — мелких хищных и насекомоядных цинодонтов, отличавшихся от других терапсид относительно крупным мозгом и, по всей вероятности, более интенсивным обменом веществ. Помимо собственно млекопитающих, млекопитающеобразные включают ряд мезозойских родов и отрядов.
Принадлежность остатков к данной кладе основывается преимущественно на строении челюстей и зубов. По уровню организации эти животные занимают промежуточное положение между цинодонтами и настоящими млекопитающими. Основные черепные признаки маммальной организации — в первую очередь, развитие костного среднего уха и зубной системы, а также особенности строения боковой стенки черепа — развивались постепенно, причем в разных филетических линиях — в разной последовательности.
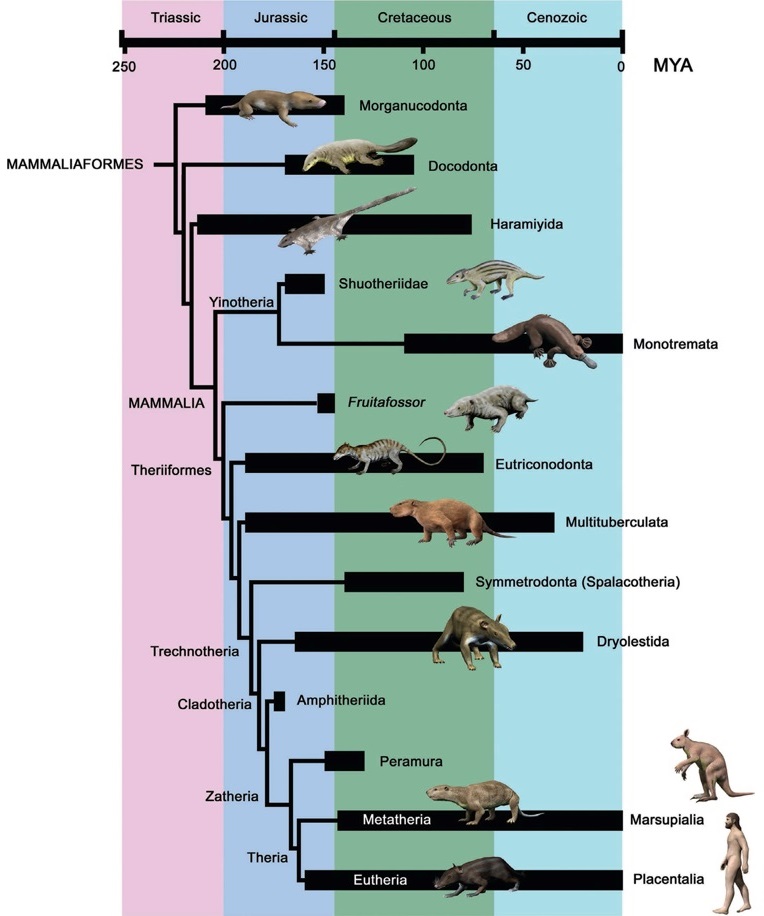
Кладограмма, отображающая родственные связи млекопитающеобразных ( по Луо с соавт., 2015 г.).
Макросистематика базальных млекопитающеобразных далека от ясности. Кроме того, что из-за небольшого количества ископаемых остатков основания классифицирования более чем скудны, еще и конкурируют разные классификационные подходы. Принимая во внимание последовательность становления маммалийного уровня организации и развития ключевых признаков, выделяются несколько град и клад.
Л. Чжэси, С. Келян-Яворовская и Р. Чифелли (2002 г.) предложили считать млекопитающими группу, включающую общего предка Sinoconodon и ныне живущих млекопитающих, а также всех потомков этого общего предка.
Большинство базальных млекопитающеобразных известно по изолированным зубам, фрагментам черепа; лишь для некоторых известно строение посткраниального скелета. Судя по реконструкциям, это были мелких или средних размеров — от землеройки до крысы — коротконогие длиннохвостые животные, покрытые короткой шерстью. Голова с сильно вытянутой мордой, глаза от небольших до довольно крупных, ушная раковина зачаточная.
Для черепа характерны незамкнутая сзади глазница и довольно крупная (в 3-4 раза больше, чем у цинодонтов эквивалентного размера) мозговая камера. По внутренним поверхностям верхнечелюстных и носовых костей проходят продольные гребни, указывающие места прикрепления обонятельных раковин, или турбиналий (у продвинутых цинодонтов соответствующие гребни четко выражены лишь на носовых костях, а турбиналии не окостеневали). В отличие от терапсид главный челюстной сустав сформирован чешуйчатой и зубной костями, тогда как сочленовная и квадратная остаются частью челюстного аппарата.

Разрозненные зубы и мелкие фрагменты костей — то, что обычно сохраняется от базальных млекопитающеобразных.
Отличающиеся по размеру и функциям зубы (гетородонтия) позволили млекопитающеобразным более эффективно измельчать пищу. За исключением очень примитивного Sinoconodon, резцы, клыки и предкоренные сменяются только один раз, а коренные не сменяются совсем. У коренных имеются 2 корня. Для большинства групп характерен специфический тип окклюзии (прикуса), при котором нижняя челюсть движется и медиально, и вверх. Смыкание зубов происходит на обеих сторонах черепа неодновременно.
Движение челюстей у базальных млекопитающеобразных, по всей вероятности, было очень похоже на известное для некоторых примитивных современных млекопитающих, например Didelphis. Изучив характер жевания последнего, возможно реконструировать его особенности у ранних млекопитающих. Ветви челюсти у Didelphis в симфизе соединены непрочно, что позволяет им двигаться относительно независимо друг от друга. Одна из самых характерных черт — то, что прикус осуществляется в каждый момент только одной ветвью челюсти. Если смотреть спереди, при этом она движется сначала латерально, затем медиально и слегка вперед, когда зубы смыкаются. При открывании рта они движутся прямо вниз. Таким образом, окклюдирующие зубы при открывании и закрывании рта описывают траекторию, близкую к треугольнику. Поперечное их движение во время смыкания и независимость двух ветвей челюсти относятся к важнейшим признакам базальных млекопитающеобразных, которые отличают их от всех цинодонтов.
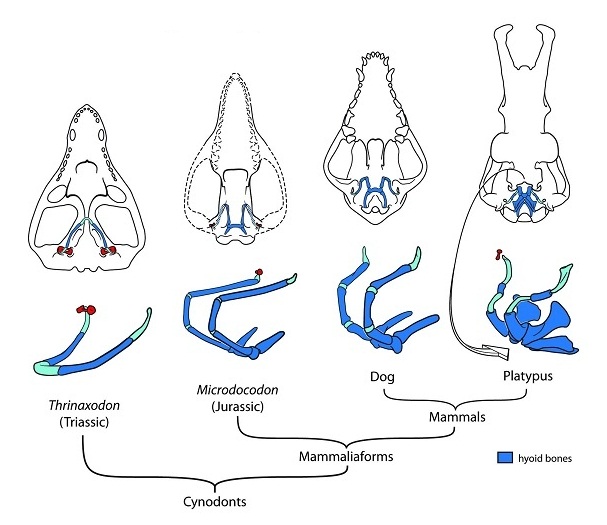
Схема развития костей подъязычного аппарата у млекопитающих и их ближайших родственников.
Некоторые из этих признаков известны как у растительноядных, так и у хищных цинодонтов, но все они одновременно ни у одного из известных цинодонтов не встречаются. И у Tritylodontidae, и у хищных цинодонтов число смен зубов уменьшено, но коренные сменяются хотя бы один раз. У Tritylodontidae щечные зубы многокоренные, но клыки и передние щечные зубы утрачены. У Gomphodontia и Tritylodontidae наблюдается окклюзия, но исключительно как результат движений нижней челюсти в вертикальной плоскости, тогда как уже у самых ранних млекопитающеобразных она включает и поперечный компонент.
В строении зубов Galesauridae и Chiniquodontidae меньше признаков млекопитающих, чем у Tritylodontidae, но нет и специализаций, отсутствующих у ранних млекопитающих. Хотя все щечные зубы сменяются более одного раза, у Thrinaxodon и Probainognathus некоторые из них похожи по строению на предкоренные и коренные. Они несут следы износа, но на нижней челюсти расположены медиальнее, чем на верхней, так что ни форма, ни размещение верхних и нижних зубов никак специфически не связаны.
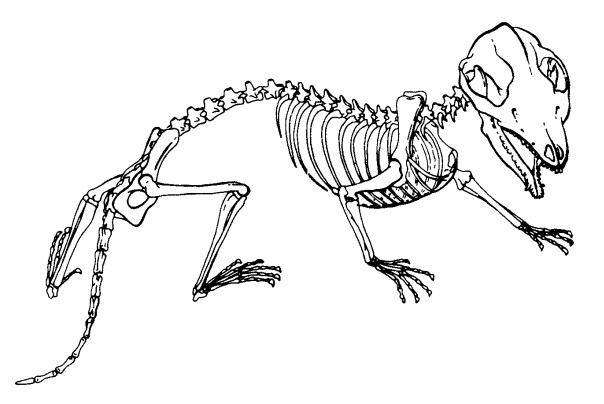
Прорисовка скелета базального млекопитающеобразного, реконструированного на основании несочлененных костей Morganucodon и неполного скелета Megazostrodon. Длина до основания хвоста 10 см.
Возможно, отсутствие специфической окклюзии у хищных цинодонтов связано с сохранением примитивной непрерывной смены зубов. Поскольку зубы сменялись неоднократно и многие из них какое-то время не были полностью функциональны, не возникало и сильного давления отбора в сторону установления точной окклюзии. За малыми исключениями, у млекопитающих коренные не сменяются вне зависимости от продолжительности жизни, а более передние зубы сменяются только один раз. У Chiniquodontidae иногда заметны следы изнашивания между латеральной поверхностью нижнего и медиальной верхнего коренных, но оно, по-видимому, не было важным компонентом жевания. У Morganucodonta изнашивание коренных зубов уже обусловлено их интенсивными жевательными движениями.
Развитие сложного рисунка из бугорков на жевательной поверхности коренных зубов происходило параллельно в разных линиях и было связано с переходом к всеядности и растительноядности. Зубные коронки при этом расширялись вбок, так как более широкие зубы лучше подходят для измельчения пищи. В связи с этим изменилось и основание зуба — у него развились двойные корни. Проведенный биомеханический анализ показал, что зубы с несколькими корнями лучше выдерживают механические нагрузки, в отличие от зубов с одним корнем, в том числе давление при смыкании верхней и нижней челюстей.
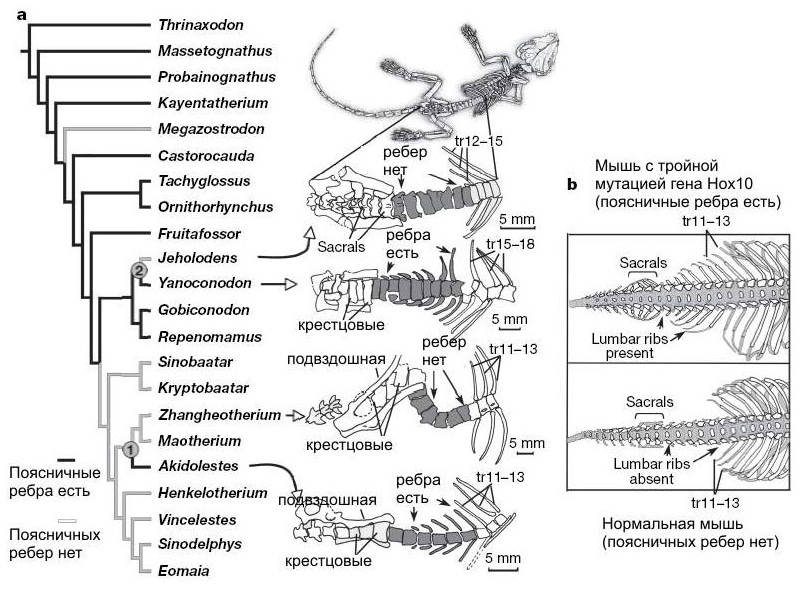
Параллельная эволюция признака "наличие / отсутствие поясничных ребер" у мезозойских млекопитающих.
Продвижение пищи кзади и ее глотание млекопитающеобразным обеспечивают подвижный язык и мускулистая глотка. Они же помогают всасывать жидкости, в частности, молоко. Диапазон движений мышц языка и глотки зависит от костей, к которым они крепятся, — подъязычного (гиоидного) аппарата, связывающего заднюю часть глотки с отверстиями пищевода и гортани. Подъязычный аппарат включает в себя мелкие кости, залегающие в мышечной толще и подвижно соединенные друг с другом. У цинодонтов гиоидный аппарат был малоподвижен и не позволял глотать небольшие порции. Современные же млекопитающие способны жевать пищу и проглатывать ее по одному маленькому кусочку за раз вместо того, чтобы глотать огромные куски или целую добычу, как рептилии.
Позвонки отличаются от типичных для терапсид и напоминают свойственные млекопитающим строением комплекса атлант - эпистрофей, расширением шейного неврального канала и выделением грудного и лишенного ребер поясничного отделов. Конфигурация невральных остистых отростков показывает, что изгибание позвоночника происходит преимущественно в вертикальной плоскости. Поясничные ребра многократно исчезали и вновь появлялись в разных эволюционных линиях млекопитающеобразных. Возможно, данное явление объясняется простотой "кодирования" этого признака, то есть тем, что для его изменения достаточно всего одной или немногих мутаций. Подобным образом могли редуцироваться и "лишние" фаланги пальцев в различных группах.
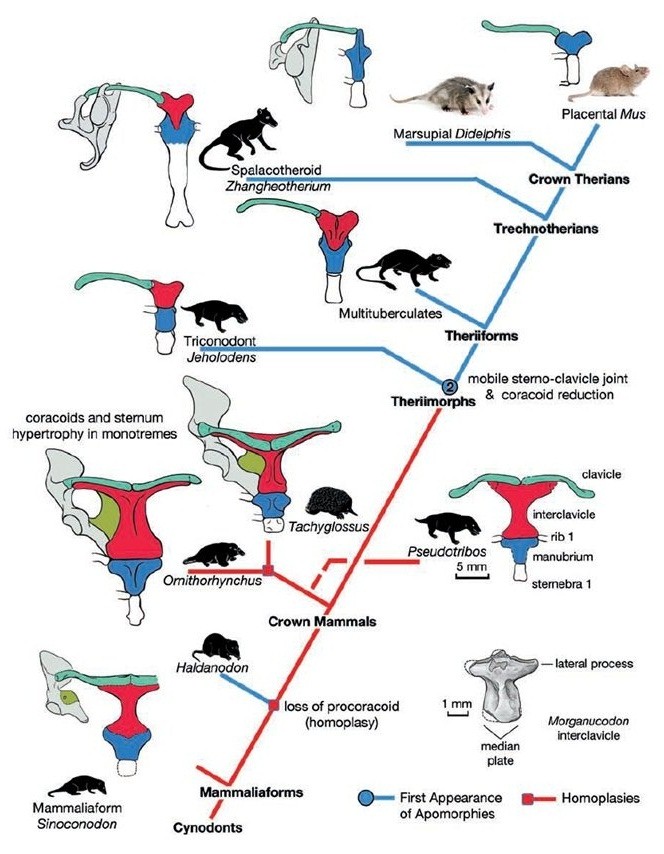
Эволюция костей плечевого пояса млекопитающеобразных.
Конечности у базальных млекопитающеобразных расположены по бокам туловища. Хотя они и не выдаются столь сильно, как у цинодонтов, но и не находятся под телом, как это характерно для типичных млекопитающих. Тем не менее, строение суставов конечностей демонстрирует возможность совершения ими движений в довольно широком диапазоне, что обеспечивало этим животным достаточную подвижность.
Пяточные шпоры выявлены у представителей отрядов Docodonta, Haramiyida, Eutriconodonta, Multituberculata, Spalacotheroidea и Monotremata. Возможно, шпоры изначально имелись у всех или почти всех базальных млекопитающеобразных и млекопитающих дотериевого уровня с расставленной постановкой конечностей. По-видимому, они исчезают одновременно с приобретением полностью парасагиттальной постановки конечностей. Высказано предположение, что шпора будет более эффективно работать у стопоходящего животного с расставленными конечностями, а при парасагиттальной постановке во время локомоции может травмировать соседнюю конечность.
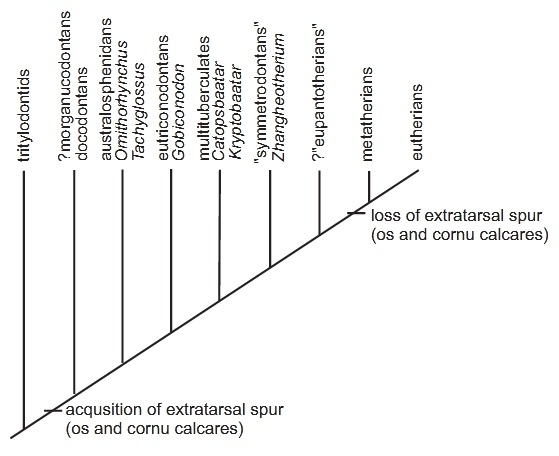
Кладограмма, иллюстрирующая распространение пяточных шпор среди млекопитающеобразных.
За исключением таранной и пяточной костей, характер расположения тарсалий и карпалий при переходе от цинодонтов к млекопитающим прослеживается лишь отчасти из-за мелких размеров этих элементов и их тенденции расчленяться при захоронении. Число централий запястья уменьшено с 2 до 1. У терапсид V-я дистальная карпалия сливается с IV-й; похожие изменения происходят и в предплюсне. I-й, или большой, палец задней конечности базальных млекопитающеобразных, возможно, несколько противопоставлялся остальным. Этот признак, а также наличие хорошо развитых когтей обычно ассоциируются с древесным образом жизни. Однако мелкие размеры могли развиться в любой сложной для локомоции среде, заставляющей часто хвататься, цепляться и карабкаться, когда функциональные различия между наземным и древесным образом жизни невелики.

Этот скелет, возможно принадлежавший Sinosauropteryx, содержит в районе пищеварительной системы три челюстные кости. Две из них идентифицированы как принадлежащие Zhangheotherium, а третья — Sinobaatar. Таким образом, млекопитающеобразные входили в рацион этого мелкого хищного динозавра. При этом известно, что на лодыжках у Zhangheotherium имелись шпоры.