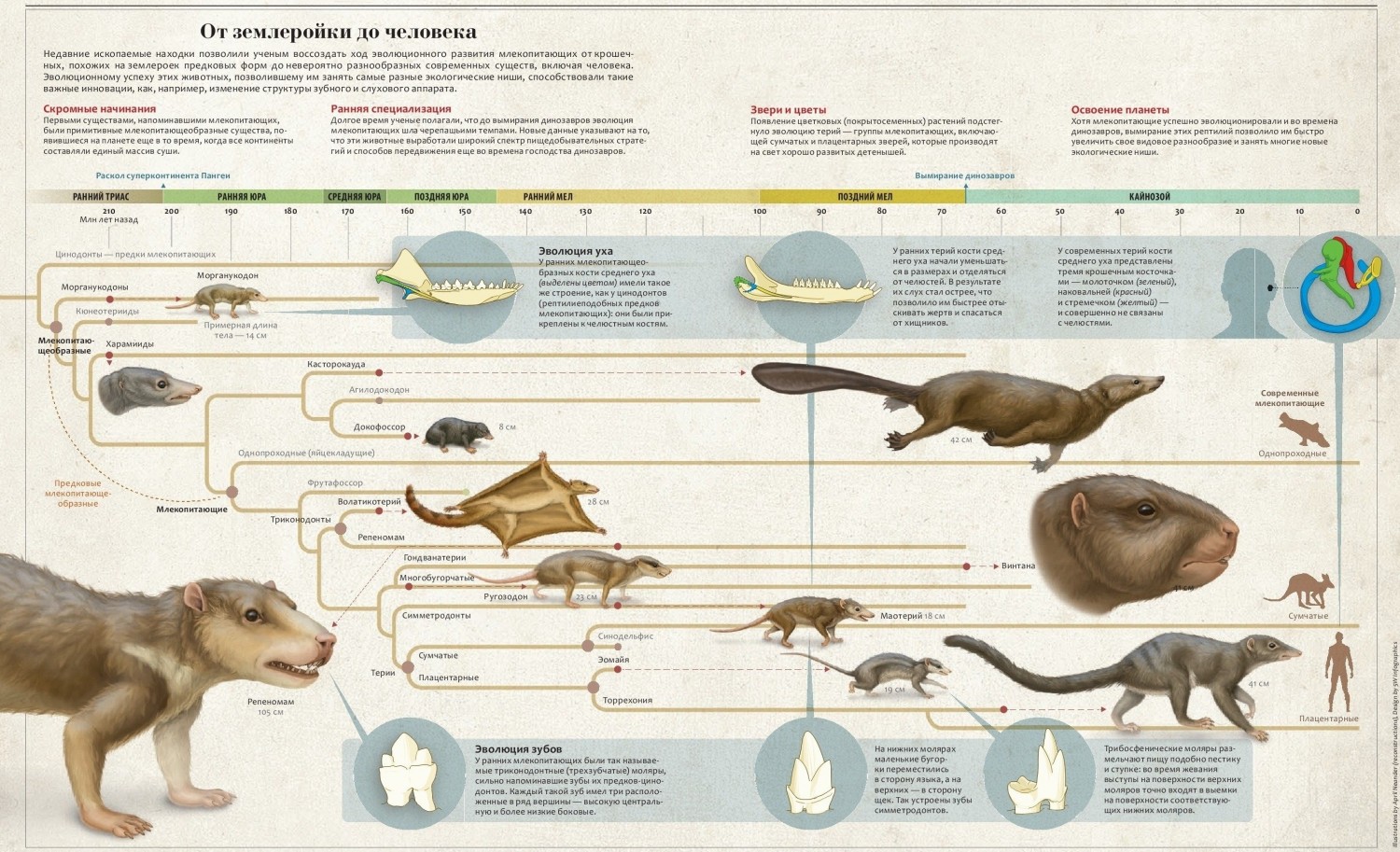
Млекопитающеобразные (продолжение)
В целом скелетные пропорции ранних млекопитающеобразных достаточно близки к таковым самых примитивных современных сумчатых и плацентарных, чтобы предполагать и физиологическое сходство, включая интенсивность метаболизма и температуру тела. Судя по всему, температурные показатели была ниже, чем у наиболее примитивных из современных млекопитающих, однопроходных, у которых они составляют всего лишь 30-32 °С. В любом случае, эти животные были эндотермными и поддерживали относительно постоянную температуру.
Современные мелкие млекопитающие поддерживают устойчивую температуру тела благодаря эффективной теплоизоляции. Специализация зубной системы и наличие вторичного неба позволяет предполагать, что эндотермия и шерстный покров были характерны уже для продвинутых цинодонтов. Наличие волос требует также присутствия сальных и потовых желез для их смазывания и контроля потерь тепла. Эмбриологическое сходство потовых и молочных желез позволяет сделать вывод, что развитие последних также могло быть тесно связано с эволюцией шерстного покрова. Наличие их у однопроходных, сумчатых и плацентарных говорит о появлении этих структур у общих предков трех групп, живших, по всей вероятности, ранее конца триаса.
Высказано предположение, что эволюция млечных желез была необходима для развития точной зубной окклюзии, характерной для позднетриасовых млекопитающеобразных. У типичных рептилий молодые особи должны быть способны к самостоятельному питанию с самого момента вылупления, поэтому от их зубной системы сразу же требуется функциональность. Так как только что вылупившиеся животные малы, зубы выпадают и сменяются несколько раз, прежде чем будут достигнуты взрослые размеры. Так как зубы неоднократно сменяются и их относительное расположение по мере роста челюстей изменяется, специфический прикус установиться не может. Появившись у базальных млекопитающеобразных, окклюзия позволила качественнее обрабатывать пищу в ротовой полости, а значит извлекать из нее максимум доступной энергии.
У предков млекопитающих вскармливание потомства материнским молоком, по-видимому, позволяло задерживать прорезывание зубов до достижения довольно значительного размера особи. Ко времени появления единственной генерации коренных позади взрослых предкоренных животное уже было довольно крупным. Становление специфического прикуса облегчалось также развитием дефинитивного челюстного сустава млекопитающих между чешуйчатой и зубной костями и более прочным прикреплением коренных зубов к челюсти несколькими корнями. Эти факторы, по всей видимости, были особенно важны для осуществления контролируемого поперечного движения челюстей при их смыкании.
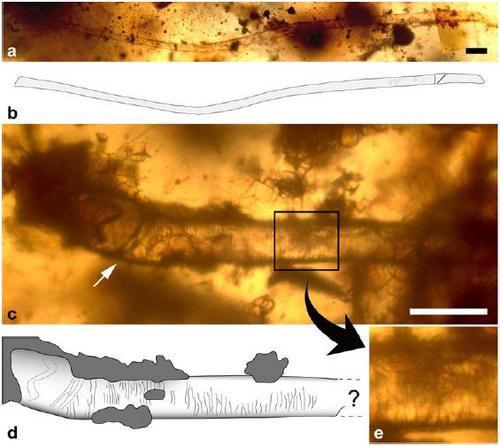
Частичные волоски, сохранившиеся в янтаре из середины мела (около 100 млн. лет назад) региона Пуату-Шаранта, Франция: a — первый фрагмент; b — прорисовка первого фрагмента; c — второй фрагмент; d — прорисовка второго фрагмента; e — крупный план второго фрагмента, демонстрирующий поверхность кутикулы. Данные образования определенно не являются протоперьями динозавров, однако точная их принадлежность неизвестна. В слое выше того, из которого происходит янтарь, обнаружено сумчатообразное Arcantiodelphys, а на соседнем участке аналогичного возраста найдены остатки различных многобугорчатых, эвтриконодонтов, спалакотероидов и дриолестоидов. Исходя из современного уровня знаний, волоски могли принадлежать любому из них. В любом случае, кутикула (защитный внешний слой волоса, образованный перекрывающимися, похожими на черепицу клетками) весьма напоминает таковую современных млекопитающих, что свидетельствует о том, что данная морфологическая особенность существует уже очень давно. Комочек янтаря, содержащий волосы, также сохранил жесткую внешнюю оболочку личинки мухи, в связи с чем высказано предположение, что волоски, возможно, происходят от уже умершего зверька, используемого мухами в пищу и для откладывания в него яичек.
Постоянная температура, поддерживаемая внутри тела, и эволюция млечных желез стали предпосылкой совершенствования размножения. Вероятно, базальные млекопитающеобразные размножались подобно современным однопроходным, т. е. вылуплялись из яиц на весьма незрелой, практически эмбриональной стадии развития. Несколько удивляет то, что примитивные млекопитающие и их непосредственные предки должны были рождаться или вылупляться совершенно беспомощными. Главное отличие рептилий от амфибий — их активное, почти взрослое состояние при появлении на свет. В то же время млекопитающеобразные, если судить по современным сумчатым и однопроходным, весили при рождении менее грамма, были слепыми и полностью зависимыми от взрослых особей в смысле защиты, обогрева и пищи.
Незрелость древнейших млекопитающих при рождении можно объяснить мелкими размерами взрослых особей, а также физиологическими ограничениями, обусловленными интенсивностью энергообмена. Мыши или землеройке на единицу веса тела требуется приблизительно в 100 раз больше пищи, чем слону. Потребность в пище и кислороде у теплокровных млекопитающих массой менее 2-3 г теоретически приближается к бесконечности. Это физиологическое ограничение препятствует существованию более мелких млекопитающих, но некоторые землеройки близки к указанному пределу. Став взрослыми, такие мелкие формы вынуждены питаться почти непрерывно, и голодание в течение всего нескольких часов приводит к их гибели. Для их детенышей физиологически невозможно поддерживать высокую температуру тела за счет собственной метаболической активности. Фактически они этого и не делают.
Детеныши мелких землероек и грызунов не эндотермны, а эктотермны. В поддержании высокой температуры тела они полностью зависят от своих родителей. Голая кожа позволяет им поглощать тепло более эффективно. Гнездо, как и тело родителей, обеспечивает теплоизоляцию. Обычно около 90 % энергии пищи, потребляемой мелкими млекопитающими, используется на поддержание высокой температуры тела. Когда детеныши избавлены от этой необходимости, почти все, что они съедают, может использоваться для роста и развития.
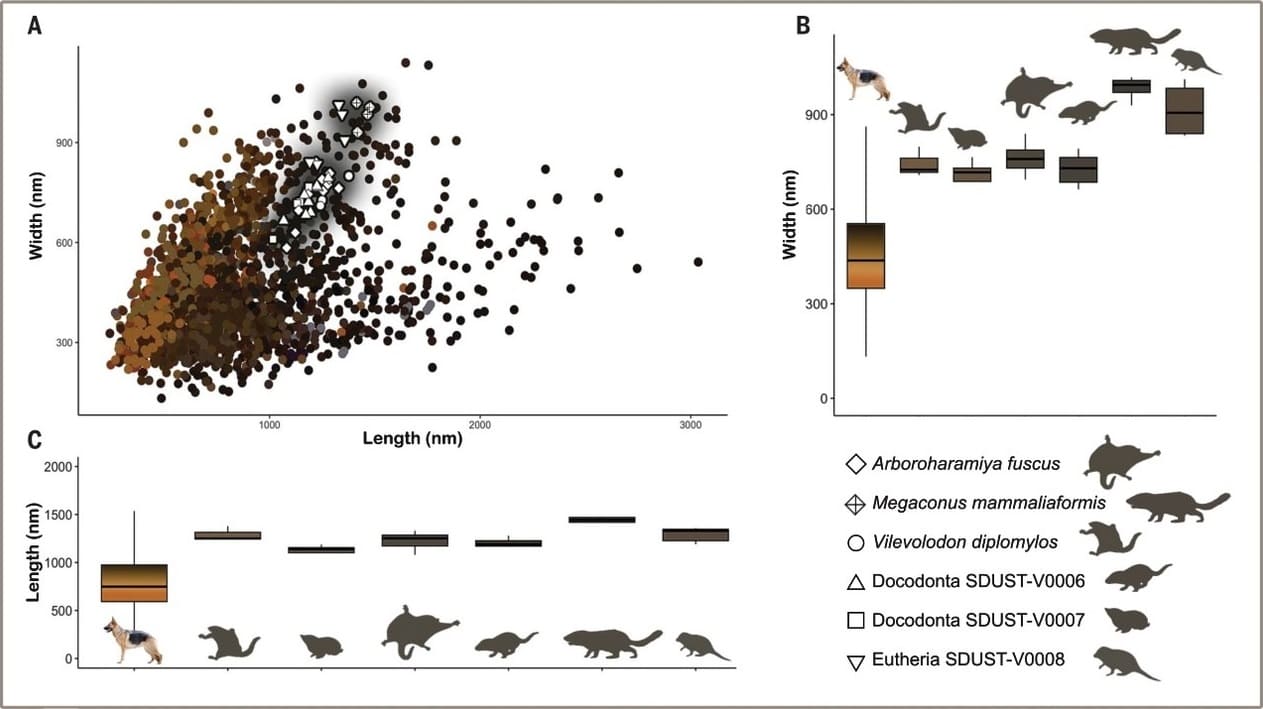
Сравнение меланосом современных млекопитающих с сохранившимися меланосомами мезозойских млекопитающеобразных из юры Китая, позволило восстановить их прижизненный окрас. Анализ отпечатков шерсти арборохарамии (Arboroharamiya fuscus) выявил высокую концентрацию меди, связанной с эумеланином, что указывает на равномерный темно-коричневый окрас без признаков цветовых узоров. Точно такой же результат был получен для вилеводона (Vilevolodon diplomylos), мегакона (Megaconus mammaliaformis), а также неопределенных докодонтов и эвтериев. Все они имели темный шерстный покров. Его окраска имеет решающее значение для эволюции поведения, физиологии и экологических предпочтений. Исследователи предположили, что изученные ими юрские млекопитающеобразные вели ночной образ жизни и потому обладали неприметной расцветкой. Более яркий и разнообразный окрас у млекопитающих мог развиться уже после вымирания динозавров в конце мела.
Достаточно высокая интенсивность метаболизма, компенсирующая быструю потерю тепла при мелких размерах тела, подразумевала высокую эффективность пищевого поведения, основанного на лучшей интеграции сенсорной информации и двигательной функции, чем у терапсидных предков. Совершенствование интеграции приводило к дальнейшему развитию высших мозговых центров. Таким образом, увеличение мозга базальных млекопитающеобразных по сравнению с цинодонтами сходного размера, видимо, связано с интеграцией возросшей слуховой, тактильной и ольфакторной информации и необходимостью координации усложнившейся локомоторной и пищевой активности, связанных с усилившимися метаболическими потребностями.
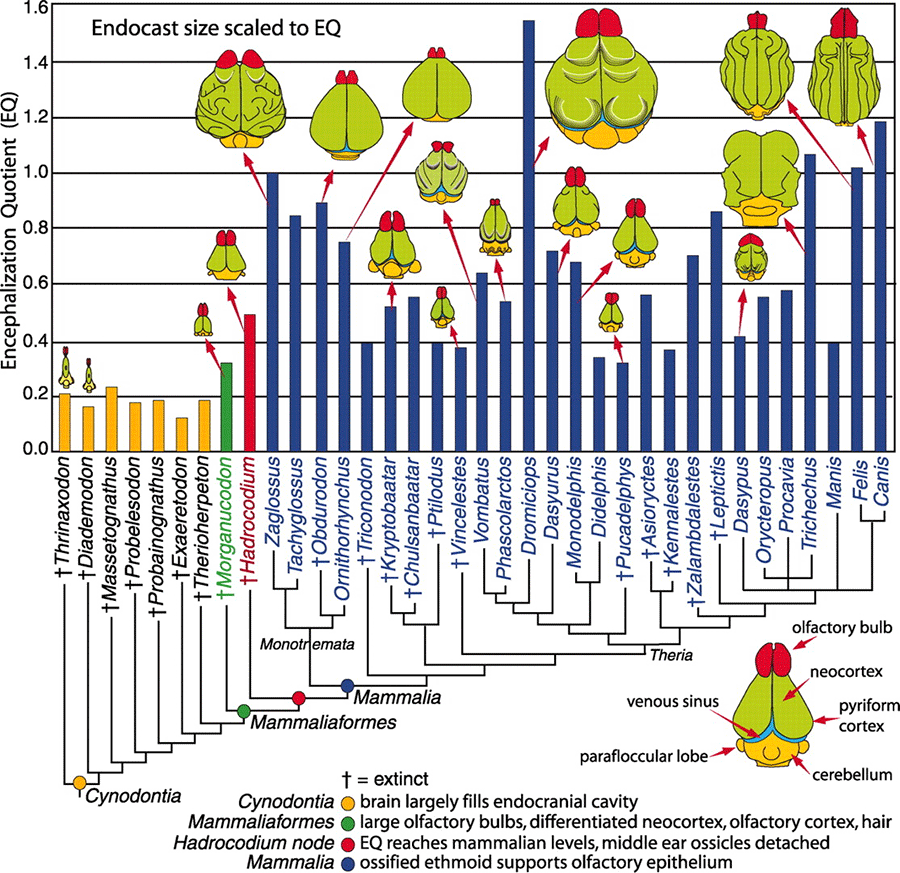
Изучение эндокастов показало, что головной мозг базальных цинодонтов Thrinaxodon и Diademodon был еще вполне "рептильный" по своему размеру и строению. В то же время, мозг млекопитающеобразных Morganucodon и Hadrocodium по своим размерам уже приближается к показателям, известным для примитивных кайнозойских млекопитающих. При этом Morganucodon стоит ближе к примитивным цинодонтам, а Hadrocodium настолько близко подошел к млекопитающим, насколько это возможно, оставаясь формально за пределами этой группы.
У базальных цинодонтов коэффициент энцефализации варьирует от 0,16 до 0,23. Их небольшие обонятельные луковицы и хрящевые турбиналии свидетельствуют о слабом развитии обонятельного эпителия. Однако обоняние, которое очень важно для самых примитивных млекопитающих, возможно, было весьма существенным уже для ранних цинодонтов. Об этом можно судить по их крупным и сложно организованным носовым полостям. Передний мозг маленький и узкий, не подразделенный на отделы, без признаков наличия неокортекса. Средний мозг и эпифиз не закрыты сверху полушариями переднего мозга. Мозжечок шире переднего мозга, спинной мозг тонкий. Эти и другие "рептильные" особенности мозга и черепа цинодонтов свидетельствуют о том, что у них, по сравнению с млекопитающими, было слабое обоняние и не слишком совершенные зрение, слух, осязание и координация движений.
Коэффициент энцефализации у млекопитающих и их родственников. Оранжевые столбики — цинодонты, зеленый — Morganucodon, красный — Hadrocodium, синие — "настоящие" млекопитающие. Вымершие формы отмечены крестиками. Отделы мозга: красным цветом показаны обонятельные луковицы, зеленым — кора полушарий переднего мозга, в том числе неокортекс и обонятельная кора, желтым — мозжечок.
Мозг Morganucodon гораздо больше похож на мозг млекопитающего. По объему он в полтора раза превосходит мозг базальных цинодонтов — коэффициент энцефализации составляет 0,32. Сильнее всего увеличились обонятельная луковица и обонятельная кора. Это со всей очевидностью указывает на развитое обоняние. Полушария переднего мозга стали выпуклыми благодаря развитию неокортекса; они закрывают средний мозг и эпифиз при взгляде сверху, как у млекопитающих. Передний мозг Morganucodon шире мозжечка, хотя мозжечок тоже заметно вырос по сравнению с базальными цинодонтами.
Увеличение мозжечка говорит об улучшенной координации движений. На это же указывает и более толстый, чем у базальных цинодонтов, спинной мозг. Расширение диапазона движений в суставах конечностей и их поясов у млекопитающеобразных по сравнению с цинодонтами должно было происходить одновременно с усложнением системы обратной связи, обеспечиваемой интероцепторами мускулатуры, что приводило к совершенствованию интегративных центров мозжечка.
Развитие неокортекса у млекопитающеобразных было связано в первую очередь с совершенствованием соматосенсорных функций. Значительная часть неокортекса у примитивных млекопитающих, таких как
Didelphis, — это так называемая соматосенсорная кора, которая отвечает за сбор и анализ сигналов, приходящих от многочисленных механорецепторов, рассеянных по всему телу. Особенно много таких рецепторов приурочено к волосяным фолликулам.
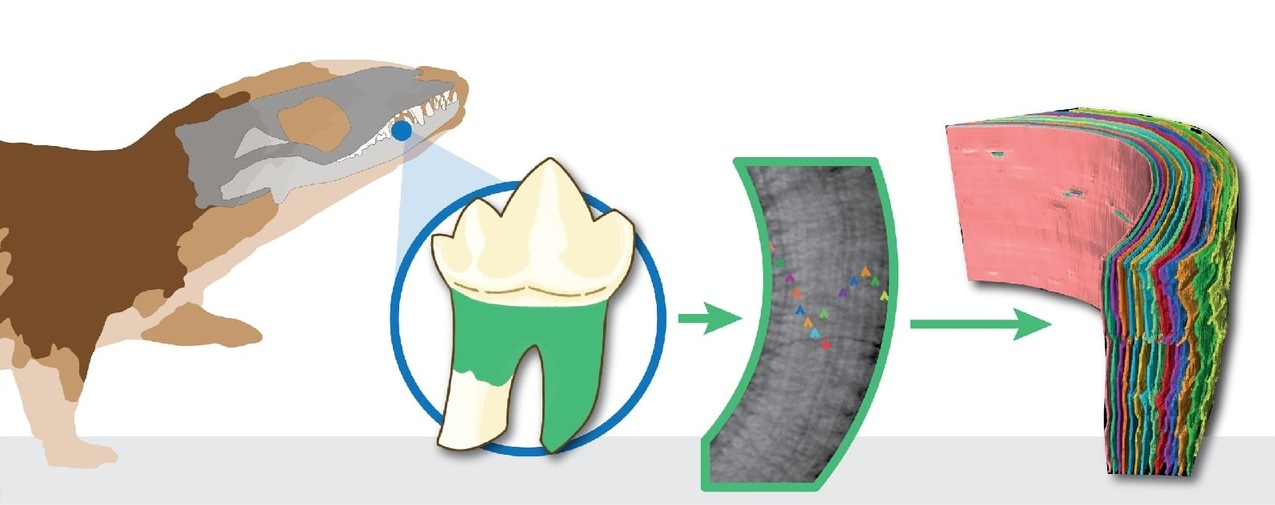
Микрофотография слоистой структуры зубной ткани
Morganucodon.
Судя по всему, волосы выполняли сначала осязательную функцию, обеспечивая поступление тактильных стимулов со всей поверхности тела. Для терморегуляции они стали использоваться позже, когда у предков млекопитающих начала развиваться теплокровность. Волосы, безусловно возникли еще до появления настоящих млекопитающих. Для ряда базальных млекопитающеобразных достоверно установлено наличие уже хорошо развитого волосяного покрова, состоявшего, как у современных зверей, из осевых волос и подшерстка. Существует предположение, что появление неокортекса у млекопитающеобразных было тесно связано с развитием волосяного покрова и осязания.
Головной мозг Morganucodon, базального представителя млекопитающеобразных, иллюстрирует первый этап прогрессивной эволюции мозга в ходе становления млекопитающих. На этом этапе увеличение мозга было обусловлено развитием обоняния, осязания и координации движений. Изменения в строении внутреннего уха свидетельствуют также о возможном улучшении слуха.
Hadrocodium, продвинутый представитель млекопитающеобразных и ближайший родственник настоящих млекопитающих, иллюстрирует второй этап развития мозга. Коэффициент энцефализации у Hadrocodium равен 0,5, то есть мозг увеличился еще в полтора раза по сравнению с Morganucodon и достиг размеров, характерных для некоторых настоящих млекопитающих. Мозг вырос в основном за счет обонятельных луковиц и обонятельной коры. Таким образом, второй этап прогрессивной эволюции мозга тоже был связан с развитием обоняния.
Погрызы, оставленные древним млекопитающим на шейном ребре завропода-мамменчизаврида из поздней юры Синьцзян-Уйгурского автономного района, Китай. Судя по всему, они возникли в результате обкусывания парными резцами мягких тканей.
В прошлом полагали, что ключевые признаки млекопитающенобразных, в том числе и их теплокровность, возникли примерно в одно и то же время. Однако в дальнейшем было показано, что несмотря на внешнее сходство, сложно устроенный мозг и продвинутые формы поведения, по уровню активности эти животные заметно уступали мелким млекопитающим типа землероек. Рентгеновские исследования микроскопической структура зубов и костей демонстрирует, с какой скоростью росло животное, что оно ело и какие проблемы с питанием вставали перед ним в детстве или в то время, когда у животного рос тот или иной набор зубов. Об этом можно узнать, измеряя толщину, плотность и состав слоев эмали и других зубных тканей. Согласно результатам исследований, у Morganucodon и Kuehneotherium из ранней юры Великобритании эмаль нарастала с той же скоростью, как и у современных им завропсид. Это говорит о том, что по темпам роста и продолжительности жизни базальные млекопитающеобразные сравнимы с рептилиями. Так, Morganucodon жил до 14 лет, а Kuehneotherium — до 9. Ранее же предполагалось, что мелкие млекопитающеобразные могли жить не более 3 лет.
Помимо анализа зубов, был оценен диаметр кровеносных сосудов в длинных костях, указывающий на максимально возможный кровоток, по которому можно судить об образе жизни и активности. Оказалось, что в бедренных костях Morganucodon диаметр сосудов выше, чем у ящериц того же размера, но намного ниже, чем у современных млекопитающих. Это предполагает, что ранние млекопитающеобразные были активнее мелких рептилий, но не настолько проворными, как современные млекопитающие.
Подобное открытие стало значительной неожиданностью. Долгое время считалось, что теплокровность и связанный с ней высокий уровень активности в любое время суток были одной из самых древних и важных особенностей млекопитающих, благодаря чему эти животные занимали те экологические ниши, на которые не могли претендовать холоднокровные рептилии. Однако необходимо отметить что это не обязательно говорит в пользу того, что Morganucodon и Kuehneotherium не являлись теплокровными. Но это свидетельствует о том, что древние родичи современных млекопитающих существенно уступали им в уровне активности. Как и когда метаболизм млекопитающеобразных приблизился к современному, еще предстоит выяснить.
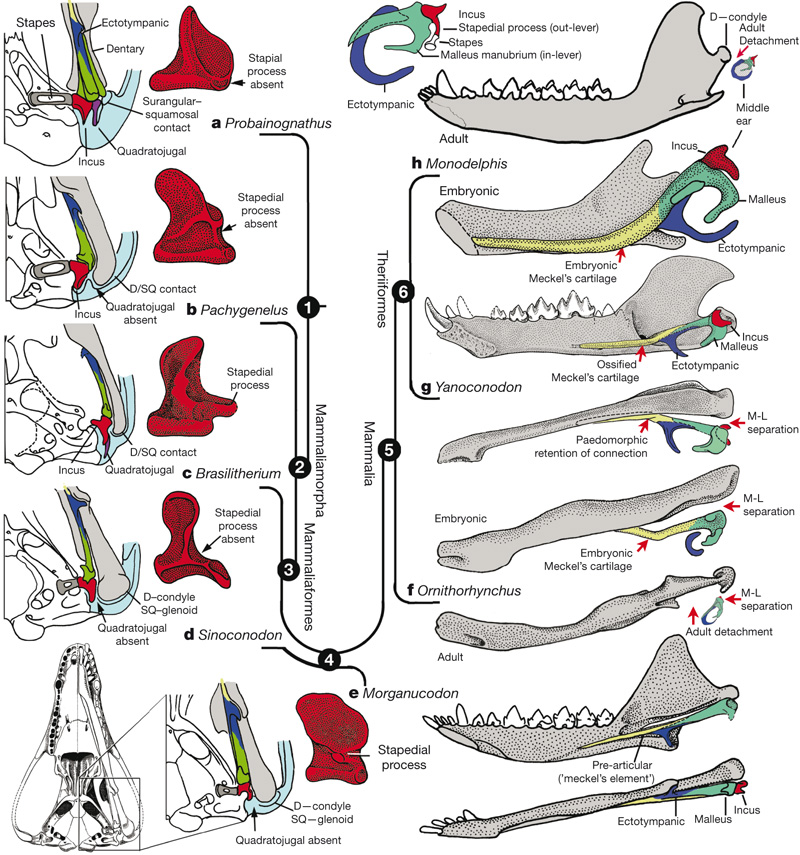
Развитие костей среднего уха у млекопитающих и их ближайших родственников.
У цинодонтов имеется только одна слуховая косточка — стремечко, а молоточек и наковальня входят в состав нижней челюсти. То же самое наблюдается и у таких базальных млекопитающеобразных, как у Morganucodon. У Hadrocodium молоточек и наковальня уже отделены от нижней челюсти, что считается одним из главных определительных признаков млекопитающих.
У современных млекопитающих в ходе эмбрионального развития эти косточки сначала, как и у далеких предков, прилегают к нижней челюсти, а затем отделяются от нее. Отделение происходит в 2 этапа: сначала слуховые косточки отделяются от боковой поверхности нижней челюсти, но остаются связаны с ней спереди посредством меккелева хряща. На втором этапе исчезает и это переднее соединение: меккелев хрящ у взрослых млекопитающих рассасывается. На основании этих фактов была выдвинута гипотеза, согласно которой в эволюции среднего уха млекопитающих должна была быть промежуточная стадия — примерно такая же, как в эмбриональном развитии современных зверей.
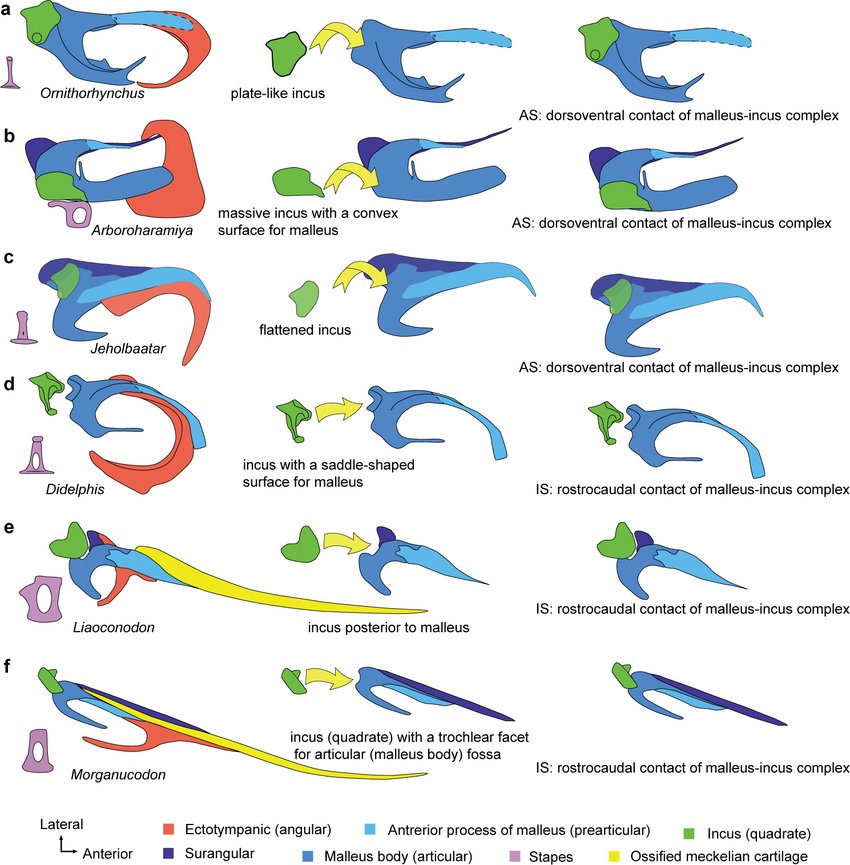
Находка хорошо сохранившихся остатков эвтриконодонта Yanoconodon блестяще подтвердил эту догадку. Его молоточек и наковальня уже отделены от нижней челюсти с боков, но еще прикреплены к ней спереди посредством окостеневшего меккелева хряща. Примечательно, что эволюция слуховой области, в процессе которой элементы нижней челюсти постепенно переходят в череп, формируя крошечные кости среднего уха, происходила самостоятельно, как минимум, в 3 разных линиях млекопитающеобразных.
Все же существуют сомнения в том, что отделение молоточка и наковальни от нижней челюсти было связано с радикальным улучшением слуха, потому что строение внутреннего уха у Hadrocodium такое же, как у Morganucodon. Судя по особенностям эмбрионального развития Didelphis, это важное изменение черепного строения может быть просто побочным следствием разрастания обонятельной коры переднего мозга. Также кажется очевидным, что переход нижнечелюстных элементов в ушную область каким-то образом связано с особенностями питания. Однако заметно более крупные размеры улитки у млекопитающеобразных все же предполагают повышение остроты слуха и более широкий диапазон восприятия частот по сравнению с поздними цинодонтами.
Схемы строения слухового аппарата различных млекопитающеобразных.
Третий этап прогрессивной эволюции мозга соответствует переходу от высших ранних млекопитающеобразных, таких как Hadrocodium, к настоящим млекопитающим. На этом этапе обоняние становится еще более тонким, о чем свидетельствуют специфические изменения решетчатой кости: на ней образуются костные носовые раковины, поддерживающие разросшийся обонятельный эпителий.
По-видимому, потребность в тонком чутье являлась главным стимулом развития мозга в ходе становления млекопитающих. У млекопитающих, как известно, обоняние развито намного лучше, чем у всех остальных наземных позвоночных. Скорее всего, это изначально было связано с приспособлением к ночному образу жизни. К концу триаса - началу юры синапсиды окончательно проиграли завропсидам конкуренцию за "дневные" ниши, и выжить удалось только тем из них, кто сумел "уйти в ночь", выработав совершенное обоняние для ориентации в потемках.
Ночью или в темных норах и расселинах также требовалась высокая острота зрения. Это косвенно подтверждается тем, что в сетчатке примитивных современных млекопитающих над колбочками преобладают палочки, отвечающие за сумеречное зрение. Кроме того, в связи с переходом к ночному образу жизни у млекопитающеобразных ослабилось цветовосприятие, поскольку они потеряли 2 из 4 генов светочувствительных белков — опсинов.
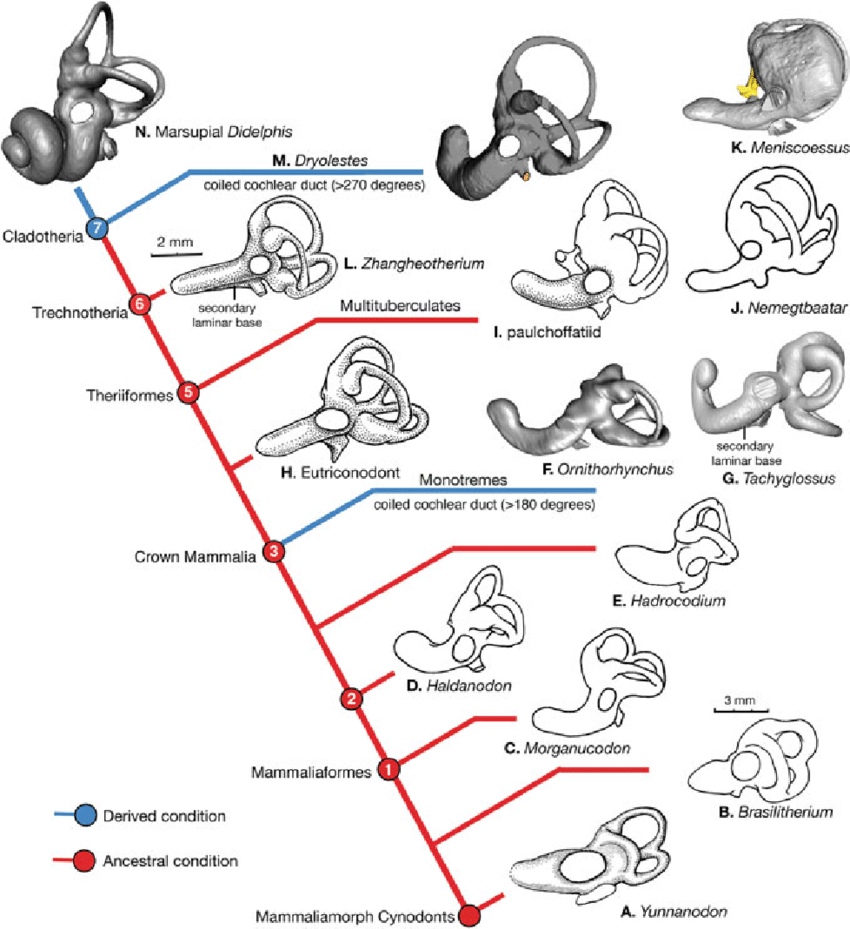
Эволюция улитки и полукружных каналов млекопитающеобразных.
Ранее предполагалось, что оба опсиновых гена были утрачены практически сразу и очень давно, еще до разделения млекопитающих на однопроходных и териевых. Однако один из потерянных генов обнаружился в геноме Ornithorhynchus. Это значит, что гены были потеряны не сразу, а по очереди, и не так быстро. Общий предок всех современных млекопитающих еще имел 3 опсина, а общий предок териевых — уже только 2. Похоже, некоторые австралийские сумчатые имеют полноценное цветное зрение, но ни одного из двух потерянных генов в их геномах обнаружить не удалось, несмотря на целенаправленные поиски. Значит, если у них действительно есть цветное зрение, оно приобретено ими вторично и на иной генетической основе.
Как это в принципе может происходить, отлично показывает пример обезьян. У общего предка обезьян Старого Света, который жил 30-40 млн. лет назад, один из двух сохранившихся опсиновых генов подвергся дупликации, и естественный отбор быстро "настроил" получившиеся копии на разные длины волн. Для этого потребовалось зафиксировать всего 3 мутации (аминокислотные замены). В итоге зрение у обезьян стало трихроматическим, что дало им возможность отличать спелые плоды от зеленых и свежую листву (наиболее питательную) от старой (у многих тропических растений молодые листья имеют красноватый оттенок). Параллельно и совершенно независимо сформировалось цветное зрение у некоторых обезьян Нового Света. У них недостающий третий опсин возник как аллельный вариант одного из двух старых опсиновых генов.

Реконструкции внешнего облика мегазостродона (Megazostrodon rudnerae), касторокауды (Castorocauda lutrasimilis), псевдотрибоса (Pseudotribos robustus), ругозодона (Rugosodon eurasiaticus), юрамайи (Juramaia sinensis), амблотерия (Amblotherium gracilis) и фрутафоссора (Fruitafossor windscheffeli). Автор — Г. Угуэто.
Как выяснилось, уменьшение числа опсинов сопровождалось увеличением числа обонятельных рецепторов, и наоборот. В отличие от цветов спектра, для различения многочисленных запахов 3-4 генами не обойтись: на каждую летучую молекулу нужен свой рецептор. Было установилено, что многочисленные гены обонятельных рецепторов млекопитающих возникли в результате множественных дупликаций из исходного небольшого набора. Судя по всему, большинство дупликаций обонятельных генов произошло примерно в то же время, что и утрата 2 опсиновых генов.
Исследования показали, что у общего предка амфибий и амниот было примерно 100-110 генов обонятельных рецепторов. Изначально большинство этих генов находилось на одной хромосоме, но время от времени они перепрыгивали на другие хромосомы. Этот процесс "рассеивания" обонятельных генов по хромосомам, по-видимому, шел уже у первых наземных позвоночных и практически прекратился к моменту расхождения линий однопроходных и териевых. В результате у всех млекопитающих обонятельные гены присутствуют почти на всех хромосомах. У человека, например, их нет только на 2 хромосомах: 20-й и Y-хромосоме. Высказано предположение, что рассеивание обонятельных генов по хромосомам облегчило их последующую множественную дупликацию.
У общего предка синапсидных и диапсидных число обонятельных генов осталось прежним (около сотни). У общего предка однопроходных и териевых их было уже около 330, у общего предка плацентарных и сумчатых их число возросло примерно до 670. Общий предок мышей и собак имел около 740 генов обонятельных рецепторов. Практически все дупликации обонятельных генов в эволюции тетрапод были приурочены к отрезку эволюционного древа, заключенному между общими предками диапсидных и синапсидных и плацентарных и сумчатых. В целом дупликации, по сравнению с "рассеиванием", позже начались и позже закончились.
Морганукодон (Morganucodon watsoni) и кюнеотерий (Kuehneotherium praecursoris) в естественном окружении. Автор — Дж. Сиббик.
Учитывая разрешающую способность примененных методик, можно заключить, что в пределах этой разрешающей способности период массовых дупликаций обонятельных генов в точности совпадает с периодом утраты опсиновых генов. Первый опсиновый ген был потерян на отрезке между общим предком диапсидных и синапсидных и общим предком однопроходных и териевых, то есть на ранних этапах становления млекопитающих. Второй опсиновый ген был потерян на отрезке между общим предком однопроходных и териевых и общим предком сумчатых и плацентарных, то есть на ранних этапах эволюции териевых. Именно к этим двум отрезкам приурочены и дупликации обонятельных генов.
Примечательно также, что восстановление цветного зрения у обезьян Старого Света сопровождалось утратой значительной части обонятельных генов (или превращением их в неработающие псевдогены). Очевидно, развитие зрения и обоняния происходило в противофазе. Когда древние млекопитающие перешли к ночному образу жизни, роль зрения уменьшилась, а роль обоняния возросла. Когда обезьяны вернулись к дневной жизни и стали снова полагаться в основном на зрение, их обоняние ослабло.
Еще одна любопытная деталь состоит в том, что одно из семейств обонятельных генов (известное как семейство №7; у приматов это самое многочисленное семейство обонятельных генов) подвергалось усиленной дупликации уже после разделения плацентарных и сумчатых. О некоторых рецепторах этого семейства известно, что они реагируют на половые феромоны.
Слева: следовые отпечатки раннего млекопитающего из поздней юры Патагонии, Аргентина.
Справа: современные следовые отпечатки мыши в песчаных дюнах штата Джорджия, США.
Может возникнуть резонный вопрос: неужели для улучшения зрения или обоняния достаточно добавить новый рецептор? А откуда возьмутся новые мозговые структуры, которые должны обрабатывать сигналы от этого нового рецептора? Судя по всему, новых мозговых структур для этого не требуется — имеющиеся в наличие структуры используют какие-то обобщенные, универсальные алгоритмы интерпретации поступающих сигналов. По-видимому, мозг в процессе индивидуального развития автоматически обучается различать сигналы, приходящие от разных рецепторов, и интерпретировать их именно как разные сигналы. Ничего не меняя в структуре мозга, можно добавить в сетчатку новый опсин, и мозг сам разберется, что делать с новым типом сигналов.
Своей внешностью и образом жизни базальные млекопитающеобразные напоминали современных насекомоядных. Их рацион состоял, по-видимому, из насекомых и других мелких животных. Небольшие размеры облегчали преследование крошечной добычи и позволяли держаться вблизи от земли в гуще растительности, где обеспечивалась относительная безопасность от нападения хищных динозавров, которые в то время являлись доминантами сухопутной биоты. Традиционно было принято считать, что ранние млекопитающеобразные были нетребовательными ночными зверьками, обитавшими среди скал и густой растительности. По мере поступления новых находок оказалось, что при сохранении довольно примитивного уровня общей организации среди них имеется немало довольно специализированных животных с роющей, полуводной или планирующей специализацией. Размножение происходило посредством откладывания яиц.
Вверху: троодон (Troodon formosus) с пойманным млекопитающим. Автор — Н. Тамура.
Внизу: череп с нижней челюстью альфадона (Alphadon halleyi) в отрыгнутом содержимом желудка предположительно троодона из позднего мела местонахождения Эгг-Маунтин в штате Монтана, США.
Исследования показывают, что по меньшей мере до средней юры для млекопитающеобразных были характерны относительно медленные темпы индивидуального развития и высокая продолжительность жизни. Судя по всему, свойственный современным мелким млекопитающим жизненный цикл, отличающийся быстрым ростом и низкой продолжительностью жизни, возник у их предков не ранее среднеюрского времени. Почему это произошло, остается неясным. Возможно, сокращение жизненного цикла у ранних млекопитающих стало следствием ускорившегося метаболизма и развития теплокровности на фоне освоения новых экологических ниш.
Ссылки
Марков А. Новая палеонтологическая находка проливает свет на раннюю эволюцию млекопитающих. — 2007 (Элементы.Ру)
перейтиМарков А. Обоняние и цветное зрение в эволюции млекопитающих развивались в противофазе. — 2008 (Элементы.Ру)
перейтиМарков А. Рост мозга у древних млекопитающих был связан с развитием обоняния. — 2011 (Элементы.Ру)
перейтиMammaliaformes (Paleobiology Database)
перейти
Литература
Кэрролл Р. Палеонтология и эволюция позвоночных. В 3 т. Т. 2 — М.: Мир, 1993
Основы палеонтологии. В 15 т. Т. 12: Земноводные, пресмыкающиеся, птицы / Гл. ред. Орлов Ю. А. — М.: Наука, 1964
Ромер А. Ш. Палеонтология позвоночных. — М. — Л.: Государственное научно-техническое издательство нефтяной и горно-топливной литературы, 1939
Россолимо О. Л., Павлинов И. Я., Крускоп С. В., Лисовский А. А., Спасская Н. Н., Борисенко А. В., Панютина А. А. Разнообразие млекопитающих: В 3-х ч. — М.: изд-во КМК, 2004