История изучения
Название "оранг-утан" малайского происхождения и переводится как "лесной человек" (примечательно, что местные земледельцы — батаки — называют так не только обезьян, но и примитивные племена лесных собирателей, например, кубу). Используемое иногда название "орангутанг" ошибочно, так как в переводе значит "человек-должник". Сообщения местных жителей о волосатых двуногих существах, известных как "седапа" или "оранг-пендек" ("маленький человек"), по-видимому, частью основаны на наблюдениях спустившихся на землю орангутанов и гиббонов, а частью на бурном воображении.
Местные жители издревле охотились на орангутанов ради полученя мяса, использовали части их тела в магических ритуалах и традиционной медицине, а порой приручали их и содержали в качестве домашних питомцев. Недюжинная сообразительность этих обезьян была давно подмечена населением региона. Так, согласно одному из поверий, орангутаны умеют хорошо разговаривать, но не делают этого при людях, чтобы те не заставили их работать.В некоторых народных сказках орангутаны спариваются с людьми и похищают их. Есть даже истории о том, как самки орангутангов соблазняли охотников.
Судя по всему, в печатной литературе слово "орангутан" впервые употребил голландец Н. Тульп в 1641 г.; правда, он обозначил им ангольского шимпанзе. Если учесть, что европейцы впервые попали на Калимантан на 100 лет раньше, малайские сказания о "лесных людях", вероятно, уже успели к этому времени широко среди них распространиться. Кроме того, мертвые орангутаны или части их тел наверняка доставлялись в страны Западной Европы суднами Ост-Индской компани. Другой голландец, Я. Бонтиус, бывший врачом на Яве, в 1631 г. употребил наименование "орангутан" в отношении волосатых человекоподобных существ, лишенных речи и живущих в лесу (вероятно, имелись в виду психически неполноценные одичавшие люди). В дальнейшем это название использовалось в нескольких работах по зоологии Ост-Индии на немецком языке.
Современное родовое название Pongo восходит к английскому моряку Э. Баттэллу, взятого в плен португальцами в Анголе. В 1613 г. он под этим названием крупную африканскую обезьяну — вероятнее всего, гориллу. Данное наименование происходит от слова "мпонго", которым в западноафриканских диалектах называли человекообразных обезьян. Таким образом, до конца XVIII в. орангутанами и понго зачастую именовали всех без разбора крупных приматов.

Старинные изображения орангутанов: 1758 г. (слева), XVIII в. (в центре), 1876 г. (справа)
В 1749 г. письменный отчет Бонтиуса вошел в «Естественную историю» К. Линнея, присвоившего "орангутану" латинское название Homo Sylvestris. В 1763 г. К.Э. Хоппиус переименовал данный таксон в Simia pygmaeus. Наконец, в 1799 г. Б.Ж. Ласепед под названием Pongo borneo описал настоящие образцы орангутана с Калимантана. В 1827 г. французский натуралист Р. Лессон описал орангутана с Суматры в качестве самостоятельного вида Pongo abelii.
На протяжении XIX и начала ХХ вв. исследователи активно охотились на орангутанов с помощью огнестрельного оружия. На основе добытых образцов были выделены многочисленные новые виды этих обезьян, что сильно запутало их таксономию. Высказывалось даже предположение, что самцы с валиками на щеках и без них относятся к разным видам. В 1926 г. Стайлз свел все виды орангутанов в единственный — Pongo pygmaeus, с подвидами Pongo pygmaeus pygmaeus и Pongo pygmaeus abelii. С этими выводом в 1929 г. согласилась Международная комиссия по зоологической номенклатуре. В 1996 г. после секвенирования митохондриальной ДНК ранг суматранского подвида вновь повысился до видового. В 2017 г. был окончательно идентифицирован третий вид орангутанов — Pongo tapanuliensis. Он тоже живет на Суматре, но теснее связан не со своим суматранским, а калимантанским сородичем.
Начиная с XIX в., появляются сотни научных трудов об орангутанах со всевозможными анатомическими и физиологическими подробностями — и зачастую с умозрительными построениями, основанными на этих характеристиках, поскольку ни один из тех авторов, по-видимому, не наблюдал этих приматов на воле. Исследованиями орангутанов в природе впервые занялась Б. Хариссон в середине ХХ в. Другая выдающаяся исследовательница, продолжившая работу на этом поприще в 1970-х гг. — Б. Гальдикас, ставшая ярой защитницей орангутанов и среды их обитания. Поведение этих приматов в дикой природе также было описано Шаллером (1961 г.), Йошибой (1964 г.) и Давенпортом (1967 г.).
Помимо этого, наряду с прочими человекообразными обезьянами, орангутаны подверглись многим лабораторным исследованиям с целью изучения их интеллекта и коммуникативных способностей. В отличие от довольно редких в природе, в неволе у них зафиксированы многочисленные случаи использования инструментов. Обезьяны также продемонстрировали способность решать сложные задания, например, открывать запертый на пряжки ящик. В рамках изучения коммуникации орангутанов обучали языку жестов и графическим символам. В начале 2011 г. было объявлено об секвенировании генома этих приматов. Исследователи выражают надежду, что добытые данные поспособствуют не только сохранению орангутанов, но и лечению генетических заболеваний человека.
Строение и функции
Орангутаны представляют собой крупных обезьян с ярко выраженным половым диморфизмом, самцы у них значительно крупнее самок. Высота сидя взрослых самцов составляет 95-100 см, самок — 70-80 см; высота в выпрямленном положении — 120-150 см (до 180 см) у самцов и 100-120 см (до 127 см) у самок. Масса тела взрослых самцов составляет 50-90 кг (в среднем 75 кг). Огромный размер и своеобразный внешний вид служат самцу для устрашения конкурентов, если те предпринимают попытки посягнуть на его территорию и полномочия. Самки примерно вдвое легче и весят около 30-50 кг (в среднем 38). Орангутаны с Калимантана и Суматры примерно равны по размерам и массе, но максимальные показатели зафиксированы у калимантанских форм. В неволе орангутаны сильно жиреют — их масса составляет в среднем 65 кг у самок и 144 кг у самцов, хотя последние могут весить и 200 кг.

Самка калимантанского орангутана (Pongo pygmaeus).
Голова орангутанов крупная, округлая. Лицевая часть широкая, несколько выдвинутая вперед и имеет сферическую форму. Лицо голое, широкое. Глаза маленькие, близко посаженные. Ушные раковины маленькие, без мочек, неоволошенные. Нос небольшой, без расширенных крыльев. Рот выступает, губы тонкие. Они способны сильно вытягиваться, особенно нижняя. Цвет лица серый, коричневатый или почти черный, у молодых животных слегка розоватый. Спереди на мощной шее имеется парный гортанный мешок, служащий для усиления голоса. Он тянется от нижней челюсти (с ответвлениями к ушам) до грудины (с ответвлениями до подмышечных впадин). Мешок имеет различный размер в зависимости от возраста и пола. Мешок взрослых самцов достигает нескольких литров в объеме и похож на подвес, тогда как у самок он развит слабее. У новорожденных мешок отсутствуеь, становясь заметным на первых месяцах жизни.
У взрослых самцов по бокам головы развиваются упругие наросты в виде полукруглых валиков до 10 см шириной и до 20 см длиной. Они представляют собой локализованные отложения подкожного жира, связанные соединительной тканью и покрытые грубой кожей со скудными и неравномерно расположенными волосками. Сверху валики сходятся на лбу, а снизу сливаются с мешком-резонатором. Со стороны кажется, будто лицо обезьяны окантовано толстой складчатой кожей. Валики продолжают расти после достижения полового созревания и наибольшего размера достигают у старых животных. Кроме того, с возрастом у самцов появляются желтоватые борода и усы, растущие больше не посредине над высокой верхней губой, а по ее бокам. У взрослых самок тоже имеется борода, но она развита не столь сильно.

Портреты взрослых самца и самки калимантанского орангутана (Pongo pygmaeus).
Телосложение орангутанов массивное и довольно неуклюжее. У них сильно развита мускулатура, обычно имеется большой круглый живот. Эти животные прекрасно приспособлены к древесному образу жизни. Их мощные передние конечности сильно удлинены, при выпрямленном положении тела доходят почти до щиколоток, а их размах у крупных особей может достигать 225 см. В целом, длина передних конечностей составляет 200 % длины туловища. Задние конечности короткие, порядка 116 % длины туловища, и относительно слабые. Кисть длинная, составляя 53 % длины туловища. Ступня составляет 62 % длины туловища.
Волосяной покров лохматый и длинный. У взрослых особей на плечах и верхних частях рук он настолько значителен, что свисает лохмами длиной более 40 см. Волосы тонкие, достаточно жесткие и относительно редкие: их плотность составляет около 160 на кв. см. на макушке, 175 на спине и 105 на груди. Пигментный слой в волосках волнистый и неровный. Шерсть несколько темнеет с возрастом: ее окрас меняется от ярко-оранжевого у молодых животных до красновато-рыжего у большинства взрослых, а также коричневатого или темно-шоколадного у некоторых особей. Кожа серовато-черная, грубо пористая. Седалищных мозолей обычно нет, они встречаются лишь изредка и имеют небольшие размеры. Выраженность ладонного дерматоглифического рисунка на 70 % больше, чем подошвенного. Рисунок подошвенного возвышения большого пальца более интенсивный, чем у возвышения мизинца, но на ладони оба возвышения равны.
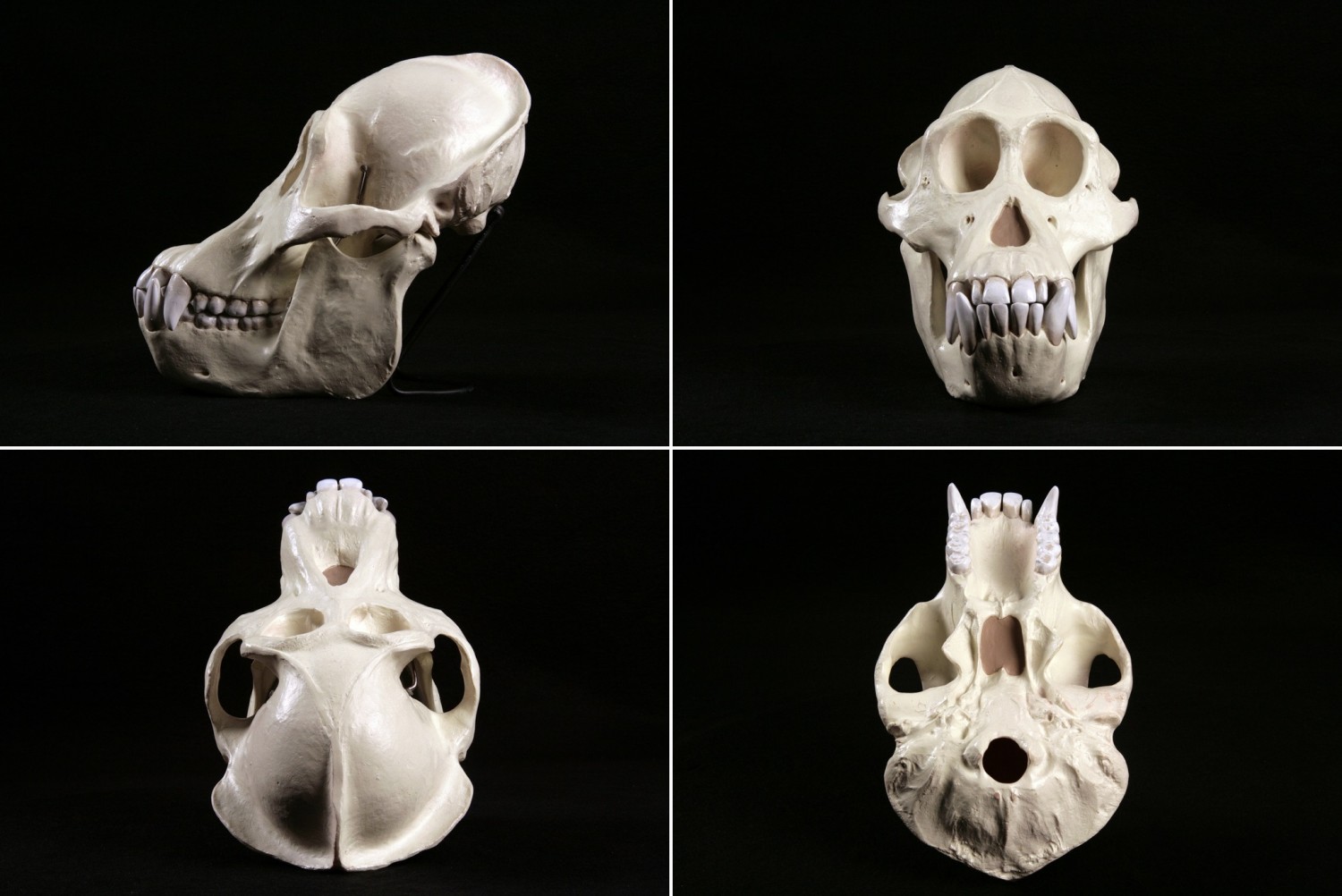
Череп калимантанского орангутана (Pongo pygmaeus).
Череп орангутанов довольно высокий, с вогнутым лицевым профилем. Верхняя и нижняя челюсти массивные, сильно выдающиеся вперед, с крупными зубами. Носовые кости сильно редуцированы в 3 % случаев. Скуловые дуги взрослых самцов очень мощные. Глазницы расположены близко друг к другу, обычно их высота больше ширины. Слезно-решетчатый шов в глазнице всегда присутствует. В отличие от горилл и шимпанзе, надбровные дуги развиты умеренно. Лоб, в отличие от большинства человекообразных обезьян, высокий и выпуклый, круто поднимающийся над глазницами. Мозговая коробка высокая и округлая. У большинства взрослых самцов сильно развит сагиттальный гребень, к которому прикрепляются массивные височные мышцы, а также лямбдоидальный гребень. У взрослых самок сохраняется округлый череп и узкий лицевой отдел молодых особей. Нижняя челюсть короткая и глубокая. Характерные очертания черепа орангутанов могут быть обусловлены сильным развитием гортанного мешка.
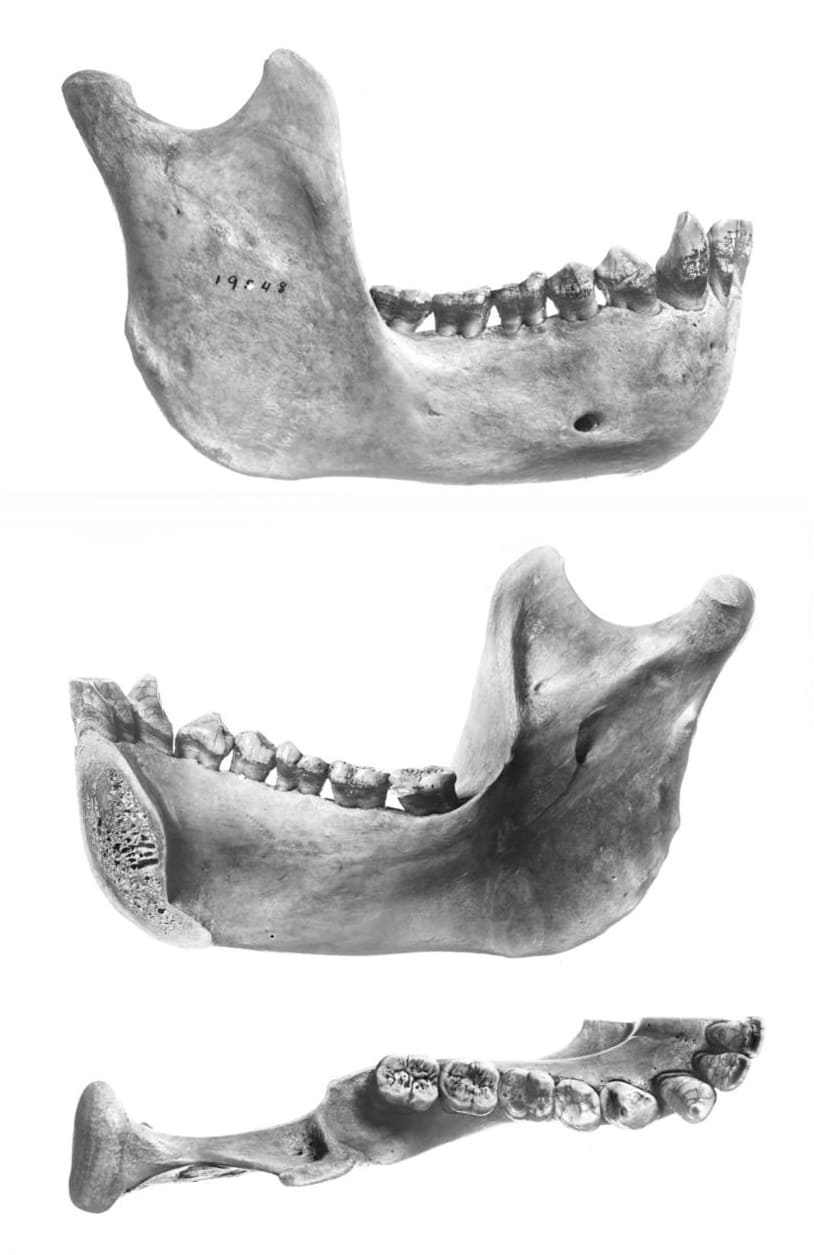
Правая ветвь нижней челюсти орангутана.
Как и у других обезьян Старого Света, зубная формула составляет I2/i2, C1/с1, P2/р2, M3/m3 = 32. Внутренние резцы широкие и лопатообразные, особенно верхние; наружные резцы маленькие и изначально заостренные. Клыки большие и толстые у самцов, гораздо мельче у самок. Предкоренные двухбугорчатые, за исключением р1, который несет уменьшенный метаконид, особенно выраженный у самцов. р1 самцов также несколько крупнее, чем у самок. Их жевательная поверхность покрыта сложным рисунком мелких бороздок и морщин, который стирается с возрастом. По меньшей мере, один лишний (IV-й) коренной зуб выявлен у 25 % самцов и 14,5 % самок и почти в 1,5 раза чаще встречается в нижней челюсти, чем в верхней. Мощный зубочелюстной аппарат орангутанов с равным успехом справляется как с мягкой, так и жесткой пищей и представляет собой отличное приспособление для срывания фруктов, веток с гнездами термитов, сдирания с деревьев коры, перемалывания твердых семян, раскалывания раковин и орехов.

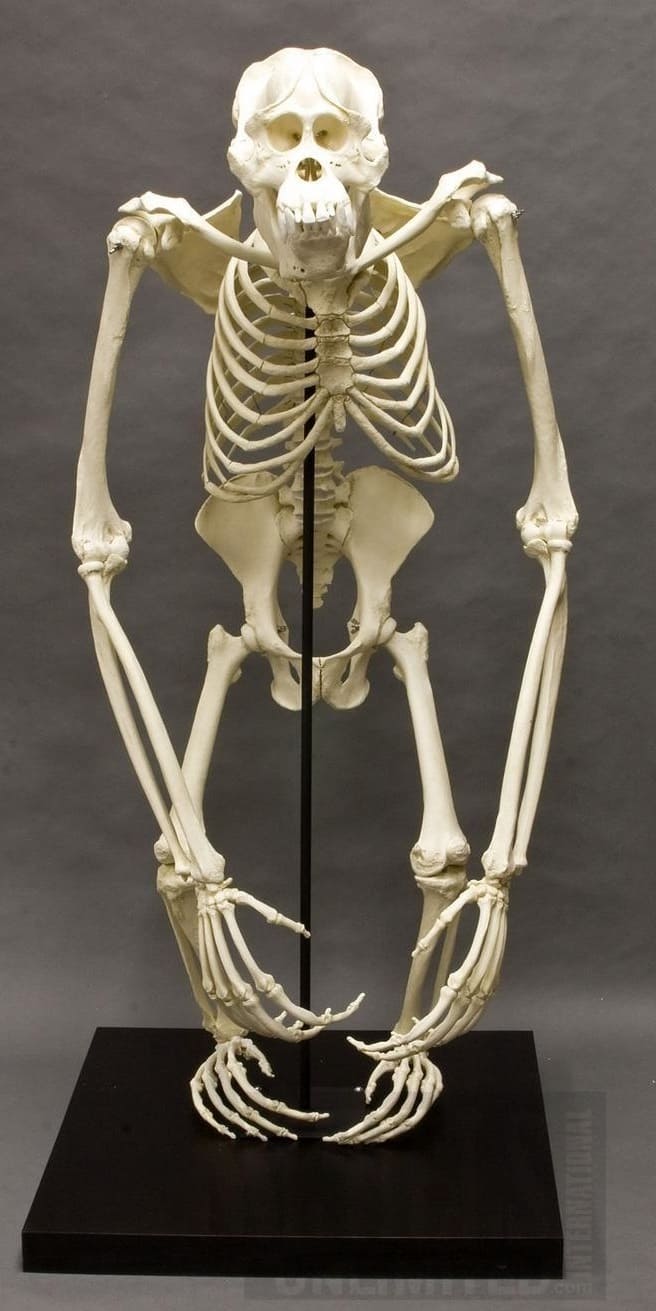
Смонтированные скелеты орангутанов; справа — скелет самца суматранского орангутана (Pongo abelii).
Позвонков 7 шейных, 12 грудных, 4 поясничных, 4-5 крестцовых и 2-3 копчиковых. Ширина грубной клетки больше, чем глубина. Лопатка расположена дорсально. Локтевая и лучевая кости длиннее плечевой, интермембральный индекс варьирует в пределах 135-150. Круральный индекс составляет 85-94 (в среднем 92). Свободный скелет конечности сходен с таковым других человекообразных обезьян, за исключением следующих особенностей: плечевая кость обычно с надрохлеарным отверстием; бедренная кость обычно не имеет ямки для круглой связки; в запястье присутствует центральня кость, имеет тенденцию сливаться с ладьевидной костью только в пожилом возрасте; пяточная кость сравнительно короткая; предплюсна короче III-й плюсневой кости; общая длина фаланг III-го пальца стопы составляет более 75 % общей длины предплюсны и плюсны.
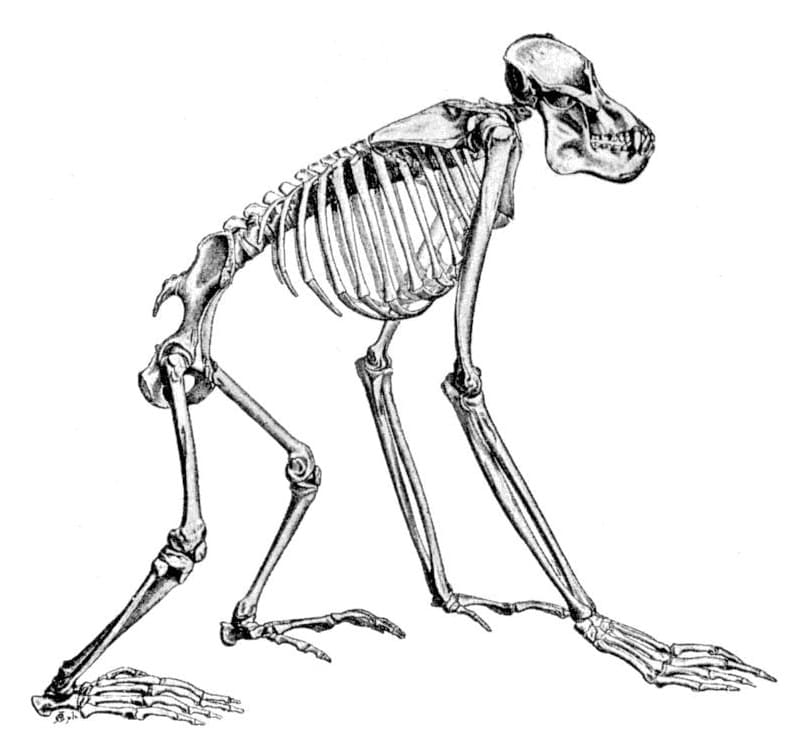
Прорисовка скелета калимантанского орангутана (Pongo pygmaeus).
Кисть и стопа внешне похожи. Метаподии и фаланги изогнутые. Кисть удлиненная и широкая. Ее I-й палец развит слабо и почти неспособен к манипуляциям, остальные пальцы длинные и сильные. При передвижении по деревьям пальцы кисти со II-го по V-й схватывают ветви подобно мощному крюку. Также эти пальцы способны надежно удерживать предметы небольшого диаметра, упираясь своими кончиками во внутреннюю часть ладони, создавая таким образом захват с двойной блокировкой. Пищевой объект во время кормления захватывается одной рукой, обычно зажимаясь между II-V-м пальцами и ладонью, поскольку I-й палец слишком короткий для эффективного манипулирования. Запястье может отводиться в несколько большей степени, чем у других человекообразных обезьян, а приводиться намного дальше — в среднем на 98 °. Пястно-фаланговые суставы пальцев кисти с II-го по V-й способны разводиться не более чем на 19 °.
В связи с жизнью на деревьях I-й палец стопы редуцирован и зачастую лишен ногтевой фаланги (отсутствует у 75,5 % самок и 46,3 % самцов), но может поворачиваться и противопоставляться остальным пальцам. Другие пальцы развиты хорошо. Стопа держится в согнутом положении и способна к схватыванию, не уступая по цепкости кисти. Длинный сгибатель I-го пальца стопы находится в зачаточном состоянии или отсутствует. Мускулатура данного пальца крайне сложна, дополнительная пара коротких сгибателей начинается у основания I-й пястной кости и прикрепляется к основанию ногтевой фаланги; присутствует глубокая головка короткого сгибателя I-го пальца.
Благодаря большой подвижности запястных и плечевых суставов, при лазании по ветвям орангутан может выворачиваться под самыми разными углами. Тазобедренный сустав также практически универсален. Обезьяна способна вытягивать ногу вниз, назад, вперед, в сторону под прямым углом и почти вертикально вверх, значительно превосходя в этом отношении человека.

Муляжи левых кисти и ступни суматранского орангутана (Pongo abelii).
Желудок продолговатый. Кишечный тракт длинный, приблизительно в 6 раз превышающий длину тела. В общих чертах кишечник сходен с таковым человека, однако его слизистая оболочка имеет многочисленные и хорошо развитые поперечные складки. Червеобразный отросток очень длинный, спиралевидной формы, но с возрастом становится относительно короче. Легкие не подразделяются на доли, с рудиментарными бороздами.
Дуга аорты дает начало безымянной и левой подключичной артериям. Безымянная артерия дает левую общую сонную артерию, а затем разделяется на правую общую сонную и правую подключичную артерию. Такая картина наблюдается у 90 % орангутанов. У человека ситуация, когда 3 артерии выходят из аорты, встречается лишь в 10 % случаев. Подключичные артерии развиты намного сильнее общих сонных артерий.
Головной мозг сравнительно большой и похож на человеческий. Его объем достигает в среднем 424 (от 320 до 540) куб. см у самцов и 366 (от 276 до 494) куб. см у самок. Орангутаны хорошо различают синий, зеленый, желтый и красный цвета. В ходе экспериментов они постоянно выбирают цветные блоки вместо серых. Одлна особь продемонстрировала значительное предпочтение красному цвету в сравнении с темно-коричневым и желтому в сравнении с оранжевым.
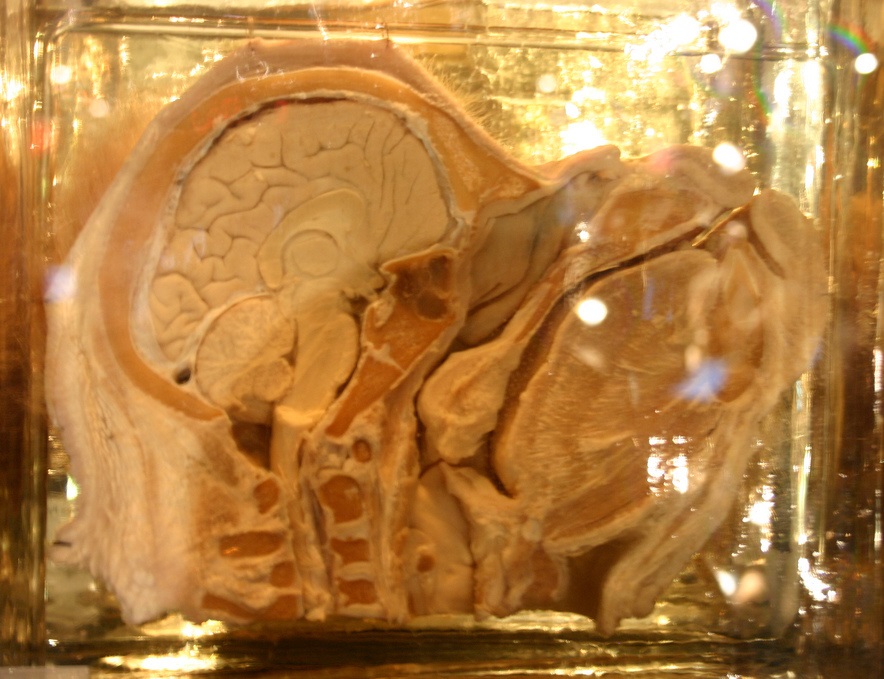
Срединный срез головы орангутана.
Установлено, что метаболизм орангутанов примерно на 30 % ниже, чем рассчитанное исходя из массы их тела значение. Согласно расчетам, на протяжении дня среднестатистический орангутан потребляет от 1100 до 2000 кал. Для сравнения: человек, который не обременен даже легкой физической работой, как правило, сжигает в день на 500-1000 кал больше. Вероятно, столь низкий уровень обмена веществ выработался у орангутанов из-за их неспешного образа жизни и сезонного минимума пищевых ресурсов.
У орангутанов выявлены группы крови А, В и АВ (отсутствует группа О) и другие компоненты крови человека. Подобно гориллам и шимпанзе, в кариотипе орангутанов 48 хромосом (в отличие от людей, у которых их 46). Основное количество плечей хромосом — 72. 26 аутосом метацентрические, 20 — акроцентрические (16 больших и 4 малых). Х-хромосома метацентрическая, Y — метацентрическая; акроцентры обладают спутниками.
Малярийный плазмодий орангутанов Plasmodium pitheci морфологически отличается от плазмодия человека и других обезьян. Наиболее частыми паразитами орангутангов являются иинфузория Balantidium coli и нематоды рода Strongyloides, из числа которых Strongyloides fuelleborni и Strongyloides stercoralis обычно встречаются у молодых особей. 19 % родов паразитических гельминтов, выявленных у орангутанов и не встречающихся среди гоминоидов в целом, являются общими с человеком, 14 % — с шимпанзе (52 % общие для человека и шимпанзе). Некоторые вши, общие для человека и шимпанзе, отсутствуют у орангутанов. У 13 % взрослых орангутанов наблюдается кариес зубов, 61 % имеют абсцессы, 34 % обладают зажившими переломами костей. Из прочих заболеваний зафиксированы стеоартрит, бластомикоз, аппендицит, стрептококовое воспаление гортанного мешка, ангина и ряд других.