Происхождение и ископаемые находки
Орангутаны (Pongo) — единственный современный род подсемейства понгин (Ponginae), которое отделилось от гоминидного ствола в раннем миоцене между 19,3 и 15,7 млн. лет назад. Предполагается, что эволюционные предшественники орангутанов не были приспособлены к жизни на деревьях, а передвигались по земле на всех четырех конечностях. В позднем миоцене на территории северной Индии и Пакистана уже обитали сивапитеки (Sivapithecus) — среднего размера полудревесные обезьяны, ранее считавшиеся предками орангутанов. Согласно современным представлениям, более вероятными претендентами на роль предков этих приматов являются кхоратпитеки (Khoratpithecus), обнаруженные в позднемиоценовых отложениях Таиланда. В достаточно близком родстве со всеми ними состояли и очень крупные гигантопитеки (Gigantopithecus) из позднего миоцена - среднего плейстоцена Южной и Юго-Восточной Азии.
Примечательно, что по сравнению с шимпанзе и людьми, ответвившимися от общего ствола человекообразных значительно позже (примерно 6 млн. лет назад), геном орангутанов эволюционировал гораздо медленнее. Исследование ДНК орангутанов показывает, что частота ее перестроек вдвое меньшая, чем у шимпанзе или у человека — соответственно 0,0027 и 0,0058 событий на ген за 1 млн. лет. Это значит, что люди и шимпанзе в ходе эволюции потеряли либо приобрели больше генов, чем орангутаны.
Неожиданной при анализе генома орангутана оказалась также низкая активность Alu-элементов (коротких повторяющихся нуклеотидных последовательностей, способных копироваться и встраиваться в различные участки ДНК). Эти элементы играли важнейшую роль в эволюции приматов. У человека, например, насчитывается 5000 специфических для него Alu-элементов, у шимпанзе — 2300. У орангутанов таких видоспецифических элементов всего 250. Возможно, этим отчасти объясняются низкие темпы реорганизации их генома. Таким образом, орангутаны должны являться гораздо более точным приближением к общему предку человекообразных обезьян, чем шимпанзе — по крайней мере, генетически.
Некоторые местонахождения ископаемых остатков орангутанов (Pongo).
На сегодняшний день орангутаны сохранились лишь в некоторых районах Калимантана и Суматры. Остатки плейстоценового возраста из южного Китая, северного Вьетнама, Лаоса, Камбоджи, Таиланда, полуостровной части Малайзии, острове Ява и, по некоторым данным, Сулавеси указывают на то, что раньше этот род имел гораздо более обширное распространение. Ареал орангутанов значительно уменьшился к концу плейстоцена, скорее всего, из-за сокращения лесной среды обитания во время последнего ледникового максимума. Более молодые остатки, обнаруженные на севере Вьетнама, а также традиционные верования в Камбодже и полуостровной Малайзии позволяют предположить, что орангутаны все еще обитали на материковой части Юго-Восточной Азии в раннеисторическое время.
В настоящее время более или менее надежно установлены 5-6 ископаемых видов орангутанов. Степень их родства с современными видами пока невыяснена, не установлена и видовая принадлежность многих находок. Последние представлены в основном разрозненными зубами, возраст которых устанавливается главным образом на основе корреляций фауны и палеомагнитного анализа. Вероятно, все зубы орангутанов, продававшиеся ранее в аптеках Шанхая, Пекина, Гонконга и Манилы, имеют ископаемое происхождение. Наблюдается отчетливая тенденции к уменьшению размера зубных рядов на протяжении плейстоцена. Однако на эту общую тенденцию, по-видимому, повлияли различные внешние факторы, которые привели к пространственно-временной неоднородности размеров зубов орангутанов.

Коренной зуб орангутана (Pongo sp.) из плейстоцена Китая.
Самым древним и крупным известным видом орангутанов является Pongo weidenreichi Hooijer, 1948, известный по зубам из ранне- и среднеплейстоценовых отложений ряда пещерных местонахождений (Лугундон, Хошантун, Цюэцюэ и др.) в провинциях Хубэй, Гуанси, Гуандун и Юньнань на юге Китая. Находки из среднеплейстоценовых слоев многочисленных пещер (Кванси, Там-Ганг, Там-Па-Лой и др.) на севере Вьетнама иногда относят к указанному виду, а порой выделяют в самостоятельный вид Pongo hooijeri Schwartz, Long, Cuong, Kha et Tattersall, 1995, таже отличающийся крупными размерами. В позднем плейстоцене в Китае и Вьетнаме указанный вид (или виды) сменил менее крупный Pongo devosi Schwartz, Long, Cuong, Kha et Tattersall, 1995, описанный из вьетнамской пещеры Ханг-Хум. Статус ряда таксонов, описанных по коренным зубам из среднего и позднего плейстоцена пещер северного Вьетнама — Pongo ciochoni из Ланг-Транг, Pongo fromageti из Там-Ом и Pongo kahlkei Schwartz, Long, Cuong, Kha et Tattersall, 1995 из Там-Хуен — нуждается в уточнении.

Реконструкция внешнего облика орангутана Хойера (Pongo hooijeri). Автор — Х. Нунду.
В Зондском архипелаге окаменелые кости орангутанов найдены на Калимантане и Суматре за пределами современного ареала, а также на Яве и Сулавеси. В некоторых из этих мест обезьяны жили еще несколько тысяч лет назад. На Яве орангутаны присутствовали, по меньшей мере, уже в среднем плейстоцене (Pongo sp. из Тринила, зубы которого не превосходят по размерам таковые современных форм) и продолжили свое существование в позднем плейстоцене как Pongo javensis Drawhorn, 1995. Последний более миниатюрен (по крайней мере, в отношении зубов), чем современные ему виды материковой Азии, Суматры и Калимантана.
Два вида орангутанов происходят из позднего плейстоцена Падангского нагорья на Суматре. Относительно крупный Pongo duboisi Drawhorn, 1995 известен из Лида Айер и Джамбу, в то время как немного более мелкий Pongo palaeosumatrensis Hooijer, 1948 найден в несколько более молодых отложениях в Сибрамбанге. Зубы последнего на 16 % больше, чем у современных орангутанов, наружные резцы менее редуцированы, а клыки и I-е нижние предкоренные крупнее; в целом в размерах зубов наблюдается ярко выраженный половой диморфизм. Эти древние суматранские виды, возможно, представляют сменяющих друх друга представителей эволюционной линии Pongo abelii, или же представляют более древнюю линию орангутанов, широко распространенную в Сундаланде и сестринскую по отношении к общей линии Pongo pygmaeus и Pongo abelii.
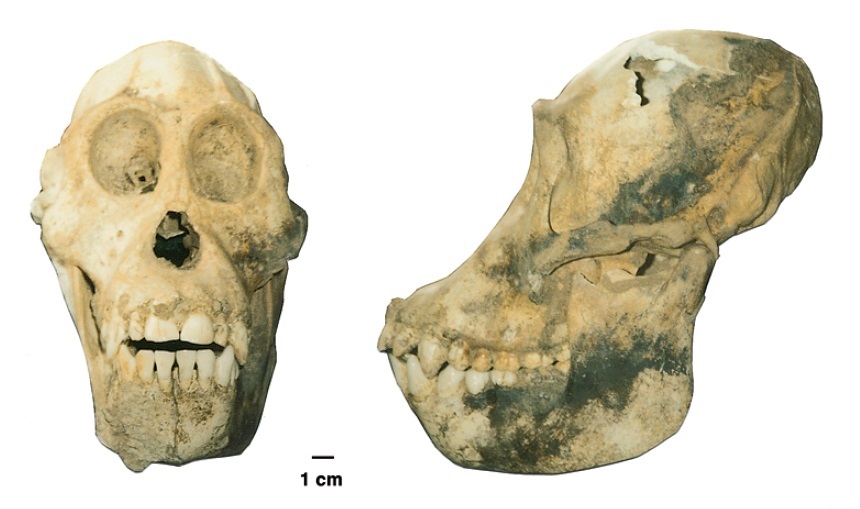
Череп от полного скелета взрослой особи орангутана (Pongo sp.) из позднеплейстоценовых отложений пещеры в провинции Хоабинь во Вьетнаме. По сравнению с современными видами, череп больше относительно туловища, крупнее зубы, интермембральный и брахиальный индексы выше. Там же собраны фрагментарные остатки молодых особей.
Наиболее многочисленные находки совершены в пещерных отложениях Калимантана и Суматры возрастом около 40 тыс. лет. Судя по сохранившимся зубам и фрагментам скелета, здешние орангутаны были примерно на треть крупнее современных. В пещере Нио из Саравака на Калимантане преобладают остатки самок и детенышей, по-видимому, представлявших для первобытных охотников более легкую добычу. А в одной пещере к северу от Нио обнаружены признаки того, что уже в позднем палеолите эти обезьяны содержались в качестве домашних животных.
Современный состав
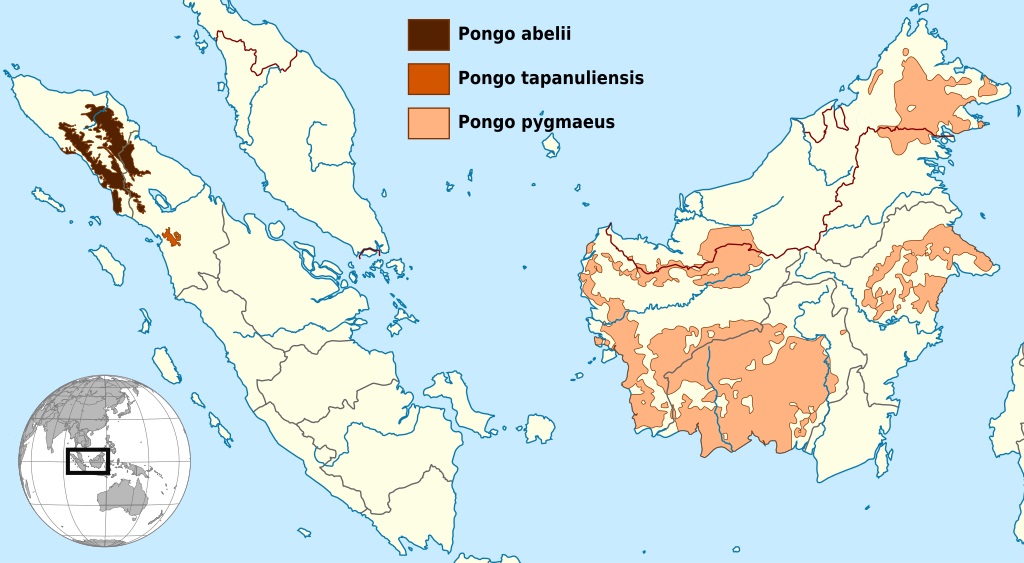
Современный ареал орангутанов (Pongo).
В настоящее время признается существование 3 современных видов орангутанов.
Суматранский орангутан (Pongo abelii Lesson, 1827) — вид, распространенный на севере Суматры.
Тапанульский орангутан (Pongo tapanuliensis Nater, Mattle-Greminger, Nurcahyo, Nowak, Manuel, Desai, Groves, Pybus, Sonay, Roos, Lameira, Wich, Askew, Davila-Ross, Fredriksson, Valles, Casals, Prado-Martinez, Goossens, Verschoor, Warren et Si, 2017) — второй суматранский вид, обитающий несколько южнее предыдущего.
Калимантанский орангутан (Pongo pygmaeus (Lacépède, 1799) Rothschild, 1904) — типовой вид, населяющий Калимантан.

Портреты самцов орангутана, слева направо: суматранский (Pongo abelii), тапанульский (Pongo tapanuliensis) и калимантанский (Pongo pygmaeus).
Генетические сравнения показывают, что общая линия Pongo tapanuliensis и Pongo pygmaeus отделилась от линии Pongo abelii около 3,4 млн. лет назад, а линии Pongo tapanuliensis и Pongo pygmaeus разошлись порядка 670 тыс. лет назад. Полагают, что современный ареал Pongo tapanuliensis максимально близок к тем территориям современной Индонезии, на которые предки орангутанов изначально пришли из материковой Азии. Популяция Pongo tapanuliensis стала более изолированной от популяции Pongo abelii после извержения вулкана Тоба, которое произошло около 75 тыс. лет назад. Однако спорадические контакты между ними продолжились и прекратились лишь 20-10 тыс. лет назад.

Степень родства ныне живущих видов орангутанов (Pongo).
Судя по результатам генетического анализа, предок Pongo pygmaeus мигрировал по Сундаленду из Суматры на Калимантан около 400 тыс. лет назад во время оледенений, когда уровень моря был ниже и острова соединялись сухопутным мостом. Тем не менее, и после разделения между популяциями Калимантана и Cуматры сохранялся очень слабый поток генов. Несмотря на то, что сейчас на Суматре живет гораздо меньше орангутанов, чем на Калимантане, генетически суматранские орангутаны гораздо разнообразнее своих калимантанских сородичей. В связи с этим можно предположить, что изначально суматранская популяция была гораздо больше, чем популяция Калимантана. Нуклеотидные последовательности калимантанского и суматранского орангутанов совпадают на 99,7 % (совпадение последовательностей орангутанов и человека — 97,4 %).