Строение и функции
Рост ореопитека в вертикальном положении достигал 1,1-1,2 м, а вес составлял 25-35 кг. Внешне он походил на невысокого, но коренастого шимпанзе (Pan troglodytes) с длинными руками. Полагают, что тело этого примата занимало более или менее вертикальное положение. Вероятно, он мог кормиться как на деревьях, так и на земле. У ореопитека, как и у многих других миоценовых обезьян, был ярко выражен половой диморфизм — самцы имели более крупные размеры, чем самки.

Реконструированный череп с нижней челюстью ореопитека (Oreopithecus bambolii).
Череп ореопитека сравнительно небольшой и низкий, округлой формы. Его лицевой отдел довольно грацилен, с умеренно выступающими вперед челюстями. Предчелюстные кости короткие и тонкие. Носовые кости короткие и выступающие. Надглазничные валики хорошо выражены. Межглазничная область широкая. Мозговая коробка относительно небольшая. Сагиттальный и затылочный гребни хорошо развиты. Верхняя челюсть массивная, с выраженными клыковыми ямками. Небо сравнительно узкое. Горизонтальная ветвь нижней челюсти утолщена, особенно в области коренных зубов. m3 не виден при рассматривании челюсти сбоку. Поверхность ветви равномерно выпукла в обоих направлениях. Восходящая ветвь высокая и широкая, а угловой отдел резко выступает назад; такое строение нижней челюсти типично для специализирующихся на листоядении тонкотелых обезьян (Colobinae).
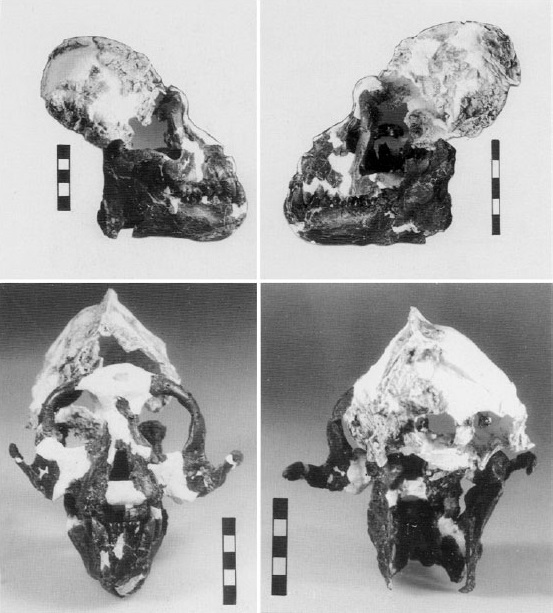
Реконструированный череп с нижней челюстью ореопитека (Oreopithecus bambolii).
Полукружные каналы внутреннего уха в целом демонстрируют типичное для человекообразных обезьян строение. Передний и боковой каналы имеют размеры примерно как у человекообразных, а задний несколько увеличен. Это говорит о сильных угловых движениях головы в сагиттальной плоскости, поскольку размер канала увеличивается у быстрых и подвижных видов для большей стабильности, а уменьшается у медленных. Например, у человека передний и задний каналы имеют увеличенные размеры для стабилизации при ходьбе, а вот боковой — маленький, так как боковые движения головы слабые.
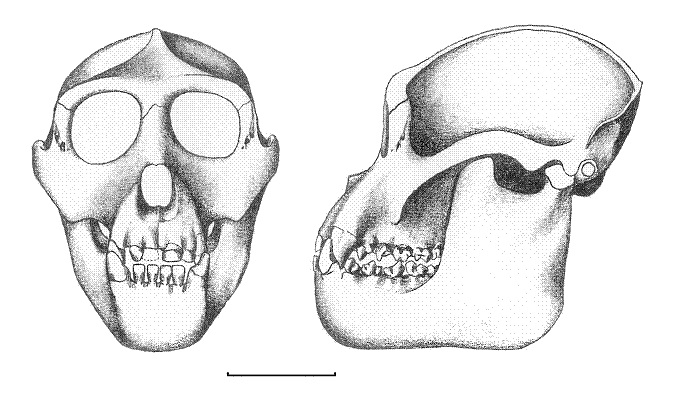
Прорисовки реконструированного черепа с нижней челюстью ореопитека (Oreopithecus bambolii).
Зубы относительно небольшие. Они демонстрируют множество примитивных черт, присущих скорее мартышковым, чем человекообразным обезьянам, а также несколько особенностей, встреченных только у ореопитека. I1 с заметным воротничком на внутренней стороне и выпуклой передней поверхностью. I2 уже и более уплощенн, чем I1. Верхний клык слегка выше Р3. С небольшим размером клыка связано отсутствие характерной для гоминид диастемы — промежутка между резцом и клыком. Верхние предкоренные с 2 бугорками. Коронки верхних коренных почти квадратной формы. Внутренние бугорки несколько смещены назад по отношению к наружным. Паракон соединен с метаконом валиком; от протокона к метакону проходит валик, отграничивающий переднюю часть зуба от гипокона. Имеется ясный или намеченный метаконуль; к нему или к его месту проходит от гипокона невысокий валик. Коронки верхних коренных окружены воротничком, особенно ясным снаружи.
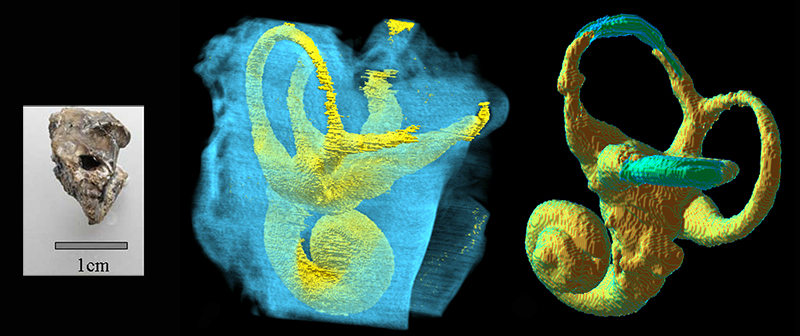
Компьютерная реконструкция внутреннего уха ореопитека (Oreopithecus bambolii).
Из нижних резцов i2 имеет более выпуклую переднюю поверхность, чем i1. Клык массивнее резцов, но почти не возвышается над соседними зубами. С небольшим размером нижнего клыка связано отсутствие характерной для гоминид диастемы между резцом и клыком на верхней челюсти. Оба нижних предкоренных с 2 корнями и с 2 бугорками. На m1 и m2 внутренние бугорки несколько смещены назад по отношению к наружным. На переднем крае m1 обычно имеется небольшой параконид, он иногда встречается на m2 и его никогда нет на m3. Позади метаконида расположен мезостилид. На середине заднего края m3 имеется гипоконулид. Мета- и гипоконид связаны невысоким валиком. В отличие от всех других приматов, на нижних коренных есть центроконид. Несмотря на то, что коронки щечных зубов несут высокие эмалевые гребни, в целом слой эмали достаточно тонок.

Фрагменты верхних челюстей самца (слева) и самки (справа) ореопитека (Oreopithecus bambolii).
Несмотря на умеренно выраженный прогнатизм и сравнительно мелкие зубы, в целом жевательный аппарат ореопитека был весьма мощным. Сильные жевательные мышцы крепились к саггитальному гребню и развитым скуловым дугам. Зубочелюстной аппарат ореопитека демонстрирует высокий уровень приспособленности к измельчению листьев; при случае также могли поедаться плоды, ягоды и, возможно, мягкие части водных растений. На это указывают не только детали строения зубов, но и их небольшие относительно тела размеры, форма и расположение в челюстях.
Хотя половой диморфизм по размерам клыков выражен вполне отчетливо, по сравнению с мартышковыми, дриопитеками, шимпанзе и другими обезьянами сопоставимого размера клыки самцов ореопитека увеличены умеренно. Это говорит о пониженной внутривидовой и особенно межсамцовой агрессии, примерно на уровне современного бонобо (Pan paniscus) — самой миролюбивой среди современных человекообразных обезьян.

Неполная нижняя челюсть ореопитека (Oreopithecus bambolii).
Грудная клетка ореопитека довольно широкая. Поясничных позвонков 5, как у сиаманга (Symphalangus syndactylus), а не 4 как у современных гоминид. Таз короткий и широкий, с поперечно ориентированными крыльями подвздошных костей. Интермембральный и брахиальный индексы очень высокие и сравнимы с таковыми у орангутанов (Pongo). Ладьевидная кость запястья сходна с таковой орангутанов, поскольку центральная кость к ней не прирастает, что говорит о небольших размерах и малой подвижности большого пальца. Фаланги пальцев как верхних, так и на нижних конечностей тонкие, удлиненные и искривленные. Изгиб фаланг кисти, хотя и превышает индивидуальный максимум человека, все же несколько меньше чем у горилл — наиболее "прямопалых" современных обезьян, находится в самых нижних пределах изменчивости обыкновенного шимпанзе, на нижней границе вариативности бонобо и далеко не достигает минимума орангутанов. Бедренная кость укороченная и прочная, с большой относительно шейки головкой. Коленный сустав отличается высокой подвижностью. Предплюсна сужена. Большой палец стопы далеко отведен от остальных, более коротких и прямых, чем пальцы современных человекообразных обезьян.
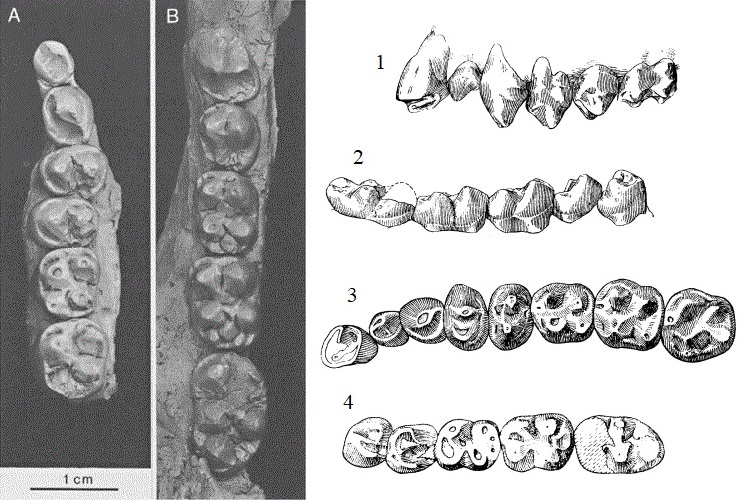
Зубной материал ореопитека (Oreopithecus bambolii) из Баччинелло, Италия. Слева: жевательные поверхности I2-M2 самки подросткового возраста (А) и Р3-М3 взрослого самца (В). Справа: прорисовки верхних (1, 3) и нижних (2, 4) зубов.
Короткое и широкое туловище, значительная длина верхних конечностей по сравнению с нижними, повышенная подвижность суставов и некоторые особенности их строения (например, глубокая суставная ямка лопатки, сходный с гоминидным локтевой сустав), приспособленная к сильным хватательным движениям длинная и узкая кисть с длинными изогнутыми пальцами, грацильная ступня, а также строение полукружных каналов внутреннего уха — все перечисленные признаки позволяют реконструировать ореопитека как брахиатора, способного ловко лазать по древесным стволам и раскачиваться на ветвях. В этом отношении он мог несколько походить на орангутанов, но значительно уступал сверхспециализированным в брахиации гиббонам, поскольку кости его рук весьма тяжелые.
Все же, по мнению некоторых исследователей, ряд особенностей в строении позвоночника, таза и конечностей указывает на то, что ореопитек обладал зачатками прямохождения и мог жить на земле. Свидетельствующие об этом анатомические черты были отмечены уже более полувека назад, однако длительное время они интерпретировались в качестве адаптаций, возникших для вертикального восхождения и спускания по стволам деревьев. Считалось, что эти черты развились из задатков, полученных ореопитеком еще от общего предка всех гоминоидов и сохранившихся в его морфологии. Таким образом, даже если у ореопитека действительно начала формироваться бипедальность, то происходило это несколько иначе, чем у предков человека.


Этапы портретной реконструкции ореопитека (Oreopithecus bambolii).
Поясничные позвонки ореопитека имеют размеры тел спереди большие, чем сзади, а сверху — меньшие, чем снизу. Это говорит о наличии поясничного лордоза и о повышенной нагрузке на крестец. Такой вариант характерен для человека и нетипичен для обезьян. В ходе исследования поясницы был выявлен и другой признак, прежде считавшийся характерным исключительно для гоминид: особое строение V-го поясничного позвонка, который смягчает толчки и удары, возникающие при ходьбе на двух ногах. Вместе с тем, размеры верхних и нижних суставных поверхностей поясничных и крестцовых позвонков, а также ширина позвоночных дуг сходны с таковыми горилл и шимпанзе. Наиболее человеческой чертой выглядит увеличение ширины между суставными отростками от первого до последнего поясничных позвонков, но оно не такое уж существенное и в той же мере встречается у обезьян. Подобно гориллам и шимпанзе, крестец ореопитека узкий, длинный, вытянутый, постепенно сужающийся книзу, тогда как австралопитеков и людей крестец очень широкий и короткий, с мощным верхом, резко сужающийся вниз. Следовательно, нагрузки на поясницу и крестец ореопитека были примерно как у современных человекообразных обезьян, а поясничный лордоз отсутствовал.
Поясничные позвонки ореопитека имеют размеры тел спереди большие, чем сзади, а сверху — меньшие, чем снизу. Это говорит о наличии поясничного лордоза и о повышенной нагрузке на крестец. Такой вариант характерен для человека и нетипичен для обезьян. В ходе исследования поясницы был выявлен и другой признак, прежде считавшийся характерным исключительно для гоминид: особое строение V-го поясничного позвонка, который смягчает толчки и удары, возникающие при ходьбе на двух ногах. Вместе с тем, размеры верхних и нижних суставных поверхностей поясничных и крестцовых позвонков, а также ширина позвоночных дуг сходны с таковыми горилл и шимпанзе. Наиболее человеческой чертой выглядит увеличение ширины между суставными отростками от первого до последнего поясничных позвонков, но оно не такое уж существенное и в той же мере встречается у обезьян. Подобно гориллам и шимпанзе, крестец ореопитека узкий, длинный, вытянутый, постепенно сужающийся книзу, тогда как австралопитеков и людей крестец очень широкий и короткий, с мощным верхом, резко сужающийся вниз. Следовательно, нагрузки на поясницу и крестец ореопитека были примерно как у современных человекообразных обезьян, а поясничный лордоз отсутствовал.
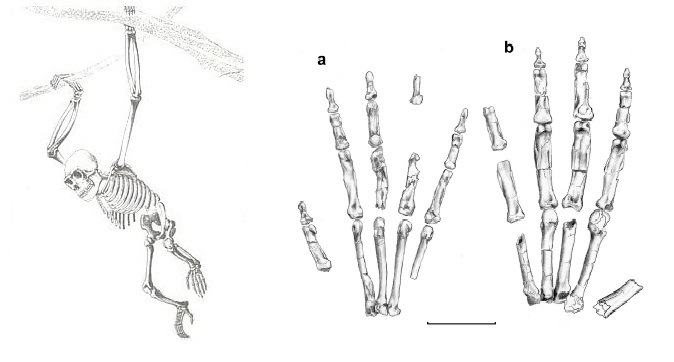
Слева — рисунок реконструированного скелета ореопитека (Oreopithecus bambolii); справа — прорисовки скелета кисти Oreopithecus (a) и Dryopithecus (b).
Таз ореопитека шире, чем у гиббоновых. На нем развиты передняя нижняя подвздошная и седалищная ости (обе отсутствуют у обезьян и имеются у людей), расстояние от ушковидной поверхности до вертлужной впадины уменьшено, а седалищная кость укорочена. Лобковый симфиз тоже короткий, а за счет тонкой и прямой формы нижней лобковой ветви в наибольшей степени соответствует варианту Люси, самки афарского австралопитека (Australopithecus afarensis), а не шимпанзе и орангутанов. Впрочем, что касается таза, его форма у ореопитека своеобразна настолько, чтобы можно было утверждать разные вещи. С одной стороны, подвздошные кости действительно широкие и низкие, несколько похожие на человеческие. С другой стороны, пропорции всего таза крайне вытянутые, сопоставимые с орангутаньим состоянием. На выпрямленную осанку ореопитека указывает и сантиметровый отросток седалищной кости, к которому крепилась сухожильная связка. Этот отросток имеется также у австралопитеков и людей, а у человекообразных обезьян он меньше или вообще отсутствует.
Таз ореопитека шире, чем у гиббоновых. На нем развиты передняя нижняя подвздошная и седалищная ости (обе отсутствуют у обезьян и имеются у людей), расстояние от ушковидной поверхности до вертлужной впадины уменьшено, а седалищная кость укорочена. Лобковый симфиз тоже короткий, а за счет тонкой и прямой формы нижней лобковой ветви в наибольшей степени соответствует варианту Люси, самки афарского австралопитека (Australopithecus afarensis), а не шимпанзе и орангутанов. Впрочем, что касается таза, его форма у ореопитека своеобразна настолько, чтобы можно было утверждать разные вещи. С одной стороны, подвздошные кости действительно широкие и низкие, несколько похожие на человеческие. С другой стороны, пропорции всего таза крайне вытянутые, сопоставимые с орангутаньим состоянием. На выпрямленную осанку ореопитека указывает и сантиметровый отросток седалищной кости, к которому крепилась сухожильная связка. Этот отросток имеется также у австралопитеков и людей, а у человекообразных обезьян он меньше или вообще отсутствует.
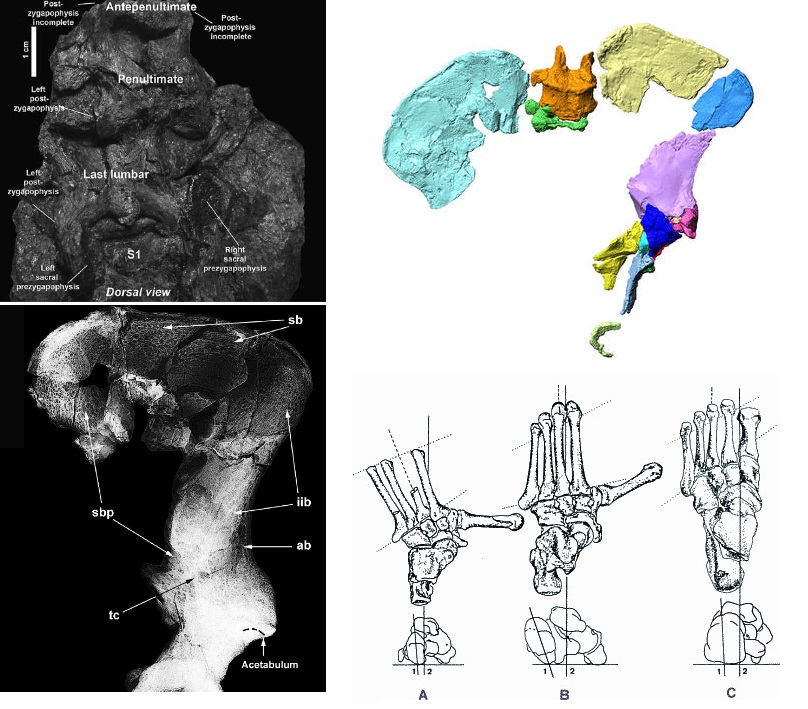
Вверху слева — крестец ореопитека (Oreopithecus bambolii); вверху справа — компьютерная реконструкция таза ореопитека (Oreopithecus bambolii); внизу слева — трабекулярная структура таза ореопитека (Oreopithecus bambolii); внизу справа — прорисовки скелета стопы ореопитека (Oreopithecus bambolii) (А), обыкновенного шимпанзе (Pan troglodytes) (В) и человека разумного (Homo sapiens) (С).
В качестве доказательства прямохождения ореопитека также приводится трабекулярная структура таза. В зависимости от нагрузок мелкие перегородочки внутри костей — стенки ячеек губчатой ткани — выстраиваются во вполне определенной последовательности и с неслучайной плотностью. Эти трабекулы располагаются вдоль преобладающих нагрузок и плотнее в тех местах, где нагрузка больше. В тазовых костях ореопитека трабекулярная структура больше напоминает человеческую, чем обезьянью. Так, сильно укреплены область ушковидной поверхности по направлению к лобковой кости, подвздошная кость спереди по направлению к седалищной кости, в том числе область передней нижней подвздошной ости, а также — задневерхний край подвздошного гребня. Усиленная полоса кости проходит у ореопитека между передней и задней частями подвздошной кости от большой седалищной вырезки до передней нижней подвздошной ости. Все эти особенности у австралопитеков и людей выражены ярче, но у гиббонов и крупных человекообразных обезьян по большей части отсутствуют вовсе. Частичное исключение составляют лишь гориллы с их большим весом, из-за которого область подвздошно-крестцового сустава в скелете этих обезьян оказывается усиленной.
Нижние конечности сравнительно недлинные, но сильные. На бедренных костях мыщелки образуют большой угол с диафизом, что также характерно для людей. Стоит отметить, что у орангутанов и гиббонов угол тоже есть, но у них внутренний мыщелок очень большой, а у людей и ореопитеков мыщелки одинакового размера. Бедро и голень при передвижении двигались в плоскости и по траектории, подобных таковым у людей и австралопитеков. Коленный сустав расположен вертикально, что обычно считается важным признаком способности к прямохождению. Особенности строения таза и костей нижних конечностей предполагают наличие достаточно развитой мускулатуры, отвечающей за передвижение на двух ногах в выпрямленном положении.

Пластическая реконструкция ореопитека (Oreopithecus bambolii), выполненная на основе представлений о его наземном двуногом образе жизни.
Наибольшей оригинальностью отличается стопа ореопитка, которая ориентирована пальцами наискось в стороны, а не вперед, как у других приматов. Внутренняя часть стопы, судя по деталям строения суставов, была предназначена держать большую нагрузку, а вот наружная не усилена должным образом. У современных обезьян усилена именно боковая сторона — для лазания по деревьям и ходьбе вразвалку по земной поверхности. Пяточный бугор ориентирован вертикально — как у человека, а не наискось, как у обезьян. Отличают ореопитека также укороченные пропорции плюсневых костей и в целом относительно небольшие размеры стопы, что также характерно для прямоходящего человека и гориллы с ее огромной массой; однако ореопитек имел небольшие размеры. Однако, своеобразнее всего ориентация большого пальца стопы: он противостоит прочим пальцам под углом около 100°. Таким образом, стопа по форме несколько напоминала птичью лапу: отведенный далеко в сторону большой палец, направленная вперед группа из II-V-го пальцев и отставленная назад пятка. Благодаря такому строению стопы ореопитек в принципе мог сохранять равновесие, передвигаясь по болотистой почве (например, в зарослях тростника), хотя и, вероятно, шаркающими короткими шажками.
Некоторые специалисты считают, что ореопитек вполне мог комбинировать древесный и наземный образ жизни. Вместе с тем нельзя отбрасывать вариант, что он действительно был двуногим прямоходящим (или, по крайне мере, склонным к прямохождению) приматом, однако передвигался таким образом не по земле, а по способным его выдержать ветвям деревьев, наподобие гиббоновых (Hylobatidae), совмещая такой тип передвижения с более характерной для него брахиацией. Выпрямленное положение тела и высвобожденные передние конечности предоставляли ореопитеку дополнительные возможности, востребованные в его своеобразной среде обитания — например, срывать плоды с высоких кустов. Эти прогрессивные особенности, как предполагается, смогли возникнуть в условиях эндемичной островной экосистемы при ограниченном количестве ресурсов, но и при почти полном отсутствии хищников.
Таким образом, исследования ореопитека делают его своеобразной моделью, на которой можно проверить целый ряд гипотез, относящихся к развитию двуногого прямохождения независимо от линии Homo, что в тоже время поможет лучше понять предпосылки и развитие двуногости у наших собственных предков.
Эволюция и вымирание
Форма коренных зубов ореопитека позволила некоторым авторам заключить, что его дальним предком мог являться апидий (Apidium) — один из раннеолигоценовых (около 34 млн. лет назад) приматов египетского Файюма. По мнению ряда исследователей, к ореопитеку наиболее близки ранние гоминиды дриопитеки (Dryopithecus) — более древние, широко распространенные европейские формы. Два этих рода имеют довольно много общих черт в строении краниального и посткраниального скелета, причем некоторые из них довольно древние и возникли еще до разделения гоминидов на линии гоминин (Homininae) и понгин (Ponginae).
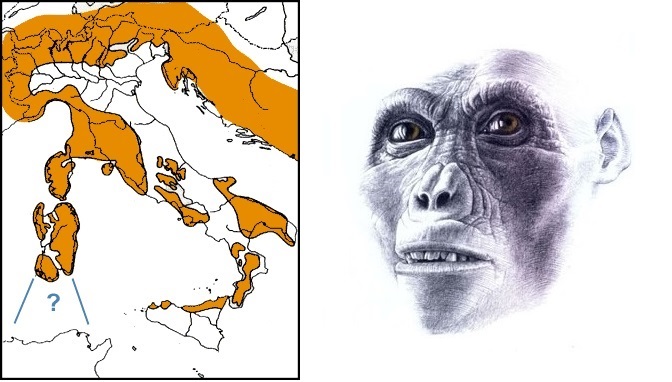
Слева — карта Италии в позднемиоценовое время (около 9 млн. лет назад); знаком вопроса обозначен возможный более ранний сухопутный путь, по которому гипотетически мог проникнуть африканский предок ореопитека. Справа — портретная реконструкция ореопитека (Oreopithecus bambolii).
Вполне возможно, что один из видов дриопитека или другого близкого к нему раннего гоминоида, оказавшись изолированным в Тоскано-Сардинийском географическом регионе, стал предковым для рода Oreopithecus. Последний возник около 9 млн. лет назад, во времена начальной фазы всемирного похолодания, постепенно превращающего климат его мест обитания из тропического в умеренный, напоминавший современный среднеевропейский. Существует также интересное предположение, что ореопитек появился от дриопитекообразного предка в результате закрепившейся неотенической мутации.
Представляется вероятным, что предки ореопитека, изолированные на окруженном морем острове или их группе, имели более мелкие размеры тела и были брахиаторами. Местообитания ореопитеков представляли собой влажные леса, перемежающиеся с кустарниковыми зарослями и болотистыми топями. Ограниченность источников питания, столь типичный для островных сообществ фактор, сделала практически невозможным проникновение сюда типичных для миоценовой Европы хищных (в фауне Гаргано присутствовали лишь мелкая и крупная рыбоядные выдры и средних размеров всеядный медведь; имелись также хищный еж, совы и крокодил). Таким образом, отсутствие крупных хищников избавило обезьян от серьезных естественных врагов и дало им потенциальную возможность спуститься с деревьев на землю, а значит получить доступ к более обильным пищевым ресурсам. Помимо этого, передвижение по земле требует меньшего расхода энергии и гораздо более безопасно, чем прыжки по деревьям, чреватые падениями и травмами. Как возможное следствие перехода к наземному образу жизни, у ореопитека наблюдается значительное увеличение размеров тела. Кроме того, по сравнению с предками изменилась его анатомия — переход брахиации к наземному передвижению способствовал развитию бипедальности и высвободил передние конечности, что сулило новые эволюционные выгоды.

"Прямоходящие" ореопитеки (Oreopithecus bambolii) в естественном окружении. Автор — М. Антон.
На протяжении примерно 2 млн. лет существования вид Oreopithecus bambolii практически не изменился, что свидетельствует об относительной стабильности среды его обитания. Позже, около 7-6,5 млн. лет назад, уровень моря значительно понизился (видимо, вследствие оледенения на полюсах планеты), и Тосканский регион соединился с материком. Находясь в эволюционном тупике, ореопитек вымер вместе с другими эндемиками, не выдержав конкуренции со стороны видов-вселенцев и давления новых хищников. Хотя ореопитек оказался бесплодной ветвью на генеалогическом древе приматов, он все равно остается своеобразной и очень примечательной вехой их эволюционного развития.