Строение и функции
Размеры продиноцеров небольшие для диноцератов, но весьма крупные для современных им млекопитающих: длина их черепа составляет 30-50 см, общая длина — 200-290 см, высота тела в наивысшей точке достигает 125 см, а вес оценивается в 150-300 кг.
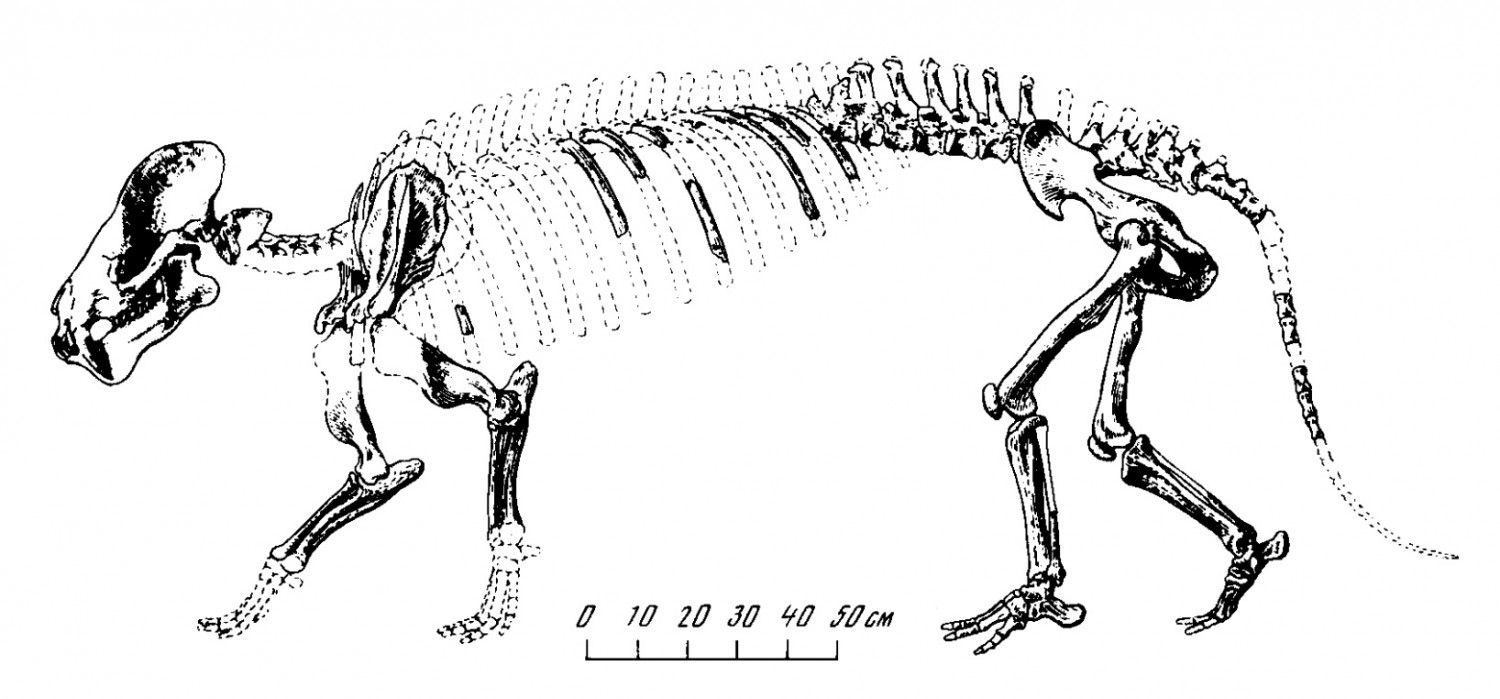
Частично реконструированный скелет продиноцера (Prodinoceras martyr) из гашатия местонахождения Наран-Булак в аймаке Умнеговь, Монголия. Палеонтологический музей им. Ю.А. Орлова РАН, Москва.
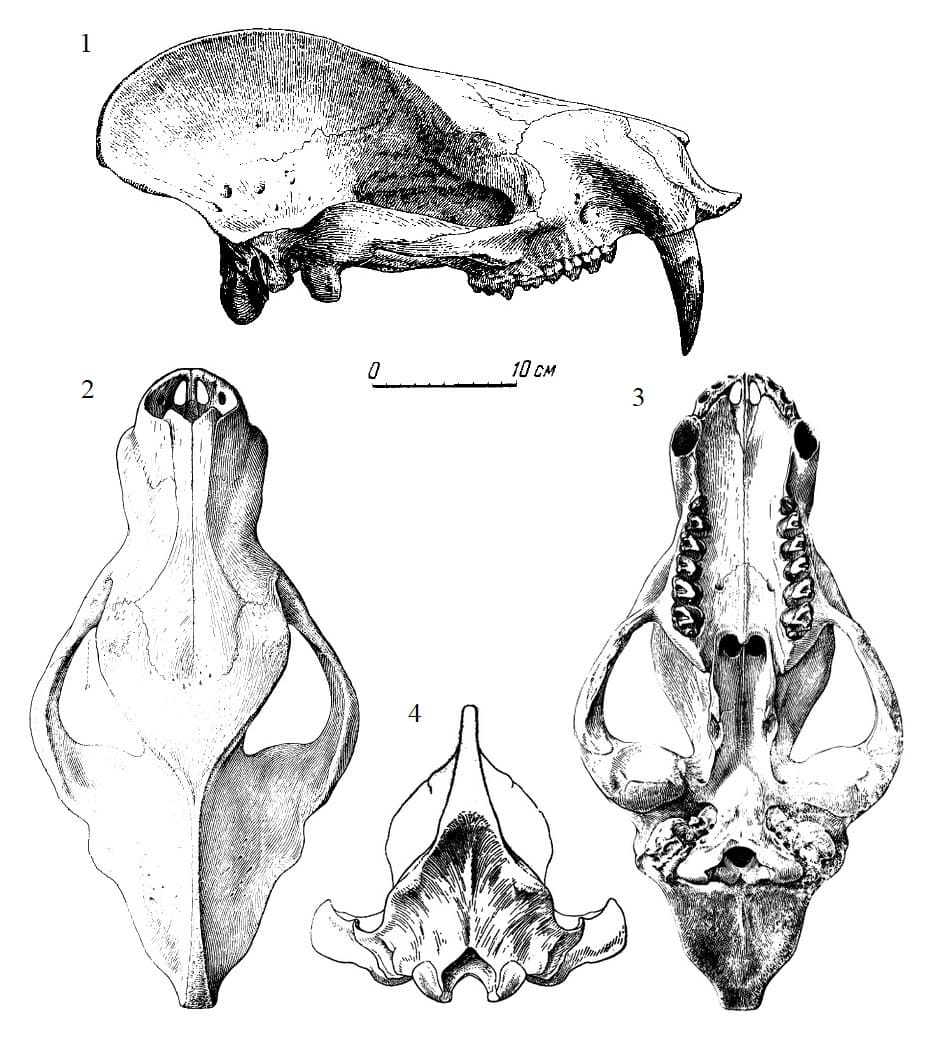
Череп крупный, удлиненный и высокий, наклоненный при жизни вперед. По форме он напоминает черепа других примитивных эвтериев, например арктоционий (Arctocyonia). Лицевой отдел почти равен по длине мозговому. Носовые кости очень длинные, почти равные половине длины всего черепа. Они простираются назад значительно дальше уровня задних верхних коренных зубов. Спереди носовые кости плотно сочленяются с предчелюстными костями и немного выступают за их края, прикрывая переднее носовое отверстие. Носовые кости толстые и массивные, особенно в передней части. Имеются большие обонятельные камеры.
Лобные кости много короче, чем носовые; в направлении назад они суживаются, выклиниваясь к сагиттальному гребню. Лобная кость плоская сверху, с небольшим вдавлением в области срединного шва заднего края носовых костей и несет здесь большое число отверстий, ведущих в лобные пазухи, которые облегчают кость в этой ее части. Глазничная и височная впадины слиты между собой. Заглазничные отростки лобных костей очень малы, а на скуловых костях отсутствуют. Скуловые дуги широко раздвинуты в стороны и равномерно выпуклы наружу. Такое строение свидетельствует о хорошо развитой височной мускулатуре. На передних наружных краях лобных костей, над глазницами, имеются бугры с шероховатой скульптурой поверхности — вероятно зачатки "рогов", подобных рогам более поздних диноцератов. Возможно, что у живых животных здесь были мозолистые или бородавчатые образования, подобные тем, которые имеются у современных бородавочников (Phacochoerus).
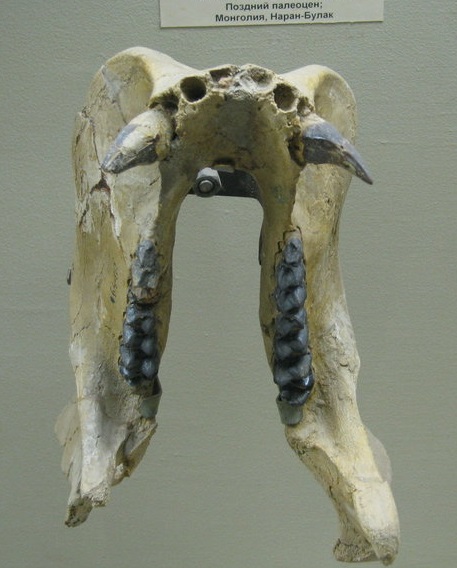
Череп с нижней челюстью и нижняя челюсть взрослого самца продиноцера (Prodinoceras martyr) из гашатия местонахождения Наран-Булак в аймаке Умнеговь, Монголия. Палеонтологический музей им. Ю.А. Орлова РАН, Москва.
Мозговой отдел вытянут, но мозговая коробка небольшая и узкая. При этом наружный объем мозговой коробки довольно большой, вследствие губчатой структуры костей и значительного развития синусов, как у всех диноцератов. Теменные кости очень слабо выпуклы латерально, так что боковые стенки мозговой коробки полого опускаются вниз и наружу. Сагиттальный гребень чрезвычайно развит, относительно сильнее, чем у каких-либо других известных млекопитающих. Поперечные затылочные гребни большие, но не раздвоенные, простые. Затылок треугольный, с сильно выступающими мыщелками.
Небная хоанальная вырезка несет небольшой средний отросток, выступающий назад. Сошник почти полностью окостеневший, делит позади хоанальный вход и опирается на задний средний отросток небных костей своим задним концом. Небные кости по краю хоаны несут выступающий гребень, окаймляющий хоанальный вход. Края хоан окаймлены валиком; на заднем краю костного неба имеется срединный выступ. Слуховой аппарат архаичного типа. Слуховая капсула как таковая не образуется, слуховые кости малы. Наружный слуховой вход широкий и не образует замкнутой трубки.
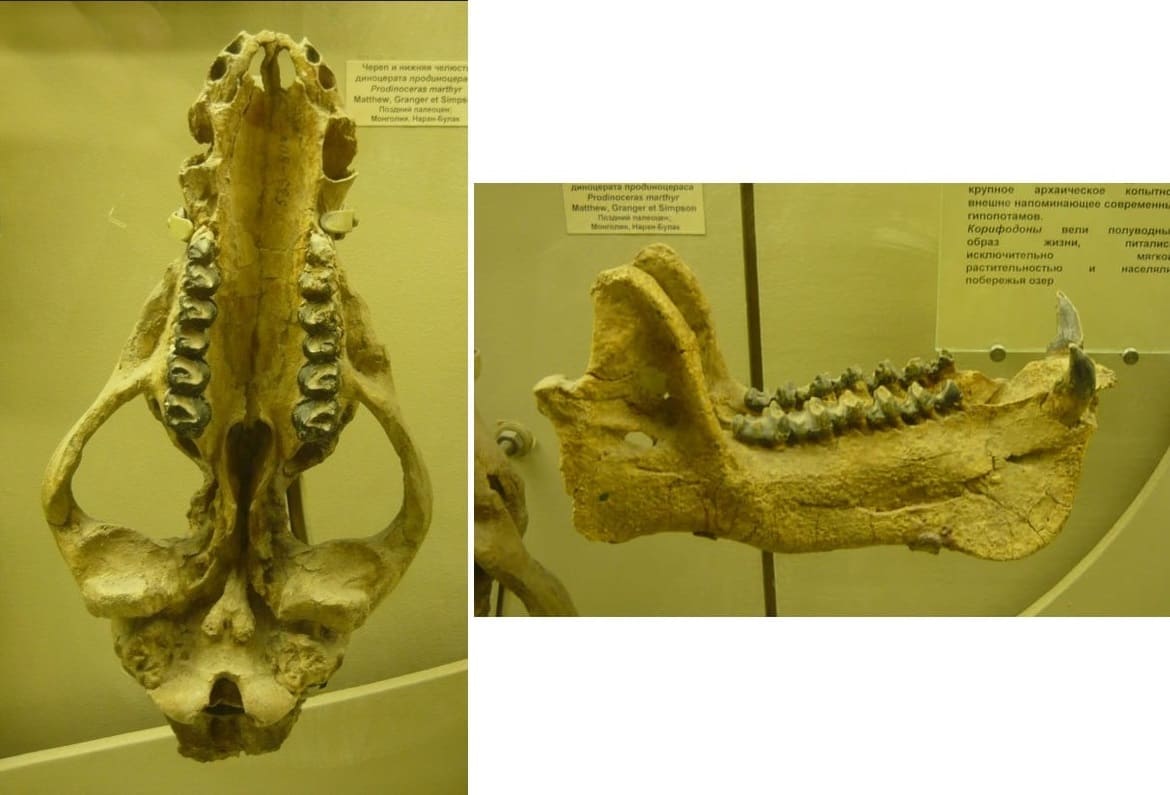
Череп и нижняя челюсть продиноцера (Prodinoceras martyr) из гашатия местонахождения Наран-Булак в аймаке Умнеговь, Монголия. Палеонтологический музей им. Ю.А. Орлова РАН, Москва.
Нижняя челюсть тяжелая с высокой восходящей ветвью, поддерживаемая при жизни сильной жевательной мускулатурой. Суставный валик находится почти на уровне коренных зубов, сочленовные бугорки расположены на ножках и выдвинуты назад, а не вверх. Подобный тип сочленовного аппарата не характерен для растительноядных копытных, а приближается к таковому насекомоядных и некоторых хищных. Столь низкое положение сустава в известной мере связано с саблезубостью, обуславливающей необходимость откидывания нижней челюсти далеко вниз. Действительно, продиноцеры, подобно саблезубым кошкам, обладали способностью чрезвычайно широко раскрывать рот и задействовать свои увеличенные клыки.
Однако необходимость сильно опускать нижнюю челюсть существенно уменьшается благодаря наличию у продиноцеров широко открытой гленоидной впадины. В результате их челюсти в разное время могли не только схватить и удержать вырывающегося противника, но и размельчать мягкую пищу. Строение нижней челюсти вместе с чрезвычайно сильным развитием гребней на черепе, увеличивающих площадь мест прикрепления смыкающей челюсти мускулатуры, свидетельствует об активной работе всего зубочелюстного аппарата.
Прорисовки частично реконструированных черепов продиноцера (Prodinoceras martyr) из гашатия местонахождения Наран-Булак в аймаке Умнеговь, Монголия: 1 — сбоку (образец ПИН 534-47); 2,3 — сверху и снизу (образец ПИН 533-117); 4 — сзади (образец ПИН 534-79).
Ветви челюсти поставлены более или менее параллельно. Массивный нижнечелюстной симфиз длинный и сросшийся. Вблизи симфиза у ряда азиатских видов имеются резко выраженные лопасти, выступающие вниз и в стороны и предохраняющие ложащиеся в них увеличенные верхние клыки. Обнаруженные нижние челюсти по величине и форме относятся к двум различным типам, что объясняется их принадлежностью самцам (более крупные и массивные, с хорошо обособленными лопастями-полуножнами) и самкам (небольшие и относительно грацильные, с значительно менее развитыми лопастями). У молодых, с молочными зубами, лопасти полностью отсутствуют. Наоборот, у американских видов лопасти выражены меньше, сзади они слабо ограничены и челюсть постепенно суживается.
Зубная формула продиноцеров очень архаична для копытных — сохраняются почти все зубы (очевидно, кроме Р1). Зубная формула: I3/i3, C1/c1, P?-3/p4-3, M3/m3. Присутствует полный набор резцов: верхние хорошо развиты, большие; нижние устроены более просто, однолопастные, крупные и заостренные. Коронки резцов уплощенные, суженные к вершине, с зазубренным волнистым краем. Резцы служили лишь для обрывания мягких растений и, возможно, захвата червей, слизняков, насекомых.
Прорисовки зубочелюстного материала продиноцера (Prodinoceras martyr) из гашатия местонахождения Наран-Булак в аймаке Умнеговь, Монголия: 1 — нижняя челюсть (образец ПИН 533-49); 2, 3 — правые верхние (Р2-М3) и левые нижние (р2-m3) щечные зубы (образец ПИН 533-117); 3 — резцы верхней и нижней челюстей при сомкнутом положении челюстей.
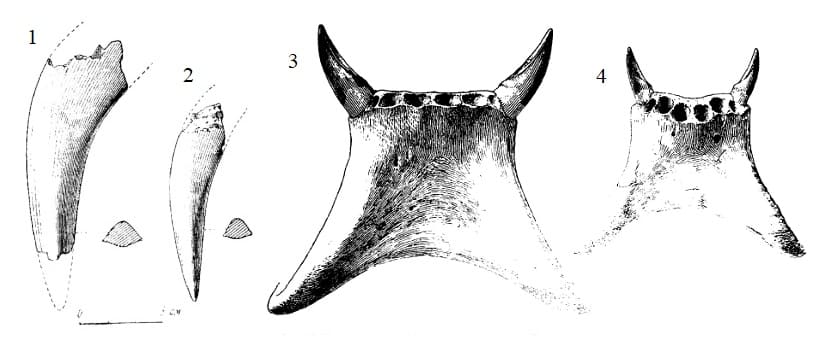
Верхние клыки большие, саблеобразные и направленные вниз, иногда прямые, но чаще загнутые назад; более крупные у самцов. Они сравнительно мало изогнуты, сдавленные — наружная поверхность округленная, внутренняя плоская. Верхние клыки обладают острыми режущими гребнями вдоль переднего и заднего краев, причем передний гребень слабо выражен, а задний слегка зазубренный. Нижние клыки почти трехгранные в сечении, конической формы, много больше резцов, однако гораздо меньше верхних клыков. На верхней и нижней челюсти между клыками и предкоренными имеются более или менее выраженные диастемы.
Предкоренные и коренные зубы низкокоронковые (брахиодонтные) и очень сходны между собой. Они имеют довольно высокие поперечные эмалевые гребни, сходящиеся на верхних зубах V-образно внутрь к протокону. На нижних, пока они не стерты, гребни покрыты мелкими бугорками. Данные гребни служили для раздавливания и разрезания пищи. У молодых имеется pd1 с 1 или 2 корнями, впоследствии он может выпадать, и у взрослых альвеола зарастает. Р2 мало моляризован, Р3-4 сходны с коренными. Энтокониды на р3-m3 небольшие, но четко выраженные; на m1-3 присутствуют бугорки энтоконида и гипоконида; m1-3 несколько вытянуты в переднезаднем направлении. Воротничок более или менее ясно выражен на всех коренных вокруг всего зуба.
Портретная реконструкция продиноцера (Prodinoceras martyr). Автор — Р. Евсеев.
Наличие у самцов продиноцеров гораздо более развитых клыков и нижнечелюстных лопастей, а также относительно большие размеры и массивность их черепов свидетельствуют о довольно ярко выраженном половом диморфизме.
Весь ряд коренных и предкоренных зубов самок при сравнении с одновозрастными челюстями самцов относительно короче чем у последних — приблизительно на длину р2. Нужно полагать, самки были мельче самцов, что явствует из размеров нижних челюстей и чем следует объяснять значительные колебания в размерах костей взрослых животных в одном и том же местонахождении. Однако верхние клыки у самок также довольно длинные, хотя и относительно тонкие — это указывает на то, что у ранних диноцератов клыки несли несколько иную функцию, чем впоследствии, когда они стали настоящим "турнирным" оружием. Впрочем, уже самцы продиноцеров при помощи своих увеличенных верхних клыков вполне могли выяснять между собой отношения.
Нижние челюсти особей продиноцера (Prodinoceras martyr) различного пола и возраста из гашатия местонахождения Наран-Булак в аймаке Умнеговь, Монголия. Палеонтологический музей им. Ю.А. Орлова РАН, Москва.
Весьма оригинальное различие самцов и самок дают резцы нижней челюсти. У самцов с широкой ложкообразной межклыковой частью все резцы поставлены в один прямой ряд и альвеолы их помещаются на одном уровне. В противоположность этому у самок альвеолы располагаются в шахматном порядке, что, вероятно, связано с узостью межклыковой части. Альвеола i3 находится прямо впереди клыка, альвеола i2 — прямо внутрь от альвеолы i3; наконец, альвеола i1, помещена вперед и внутрь от i2; таким образом, альвеолы средних резцов выдвинуты вперед.
С возрастом у продиноцеров происходило удлинение лицевой части черепа, а также удлинение диастемы между клыками и предкоренными, так что у средневозрастных самцов она равна приблизительно длине m1 + m2, а у старых самцов превышает ее. Сообразно с увеличением размера клыков увеличивается и ширина всей носовой области. Чрезвычайно сильно разрастаются все гребни, и весь череп становится более массивным.
Слева: прорисовки верхних клыков продиноцера (Prodinoceras martyr) из гашатия местонахождения Улан-Булак в аймаке Умнеговь, Монголия: 1 — самец (образец ПИН 534-83); 2 — самка (образец ПИН 534-84).
Справа: прорисовки частично реконструированных нижних челюстей продиноцера (Prodinoceras martyr) из гашатия местонахождения Наран-Булак в аймаке Умнеговь, Монголия: 1 — самец (образец ПИН 534-49); 2 — самка (образец ПИН 534-1).
Предохранительные лопасти, отсутствующие у молодых животных, появляются с началом отрастания верхних клыков. Сначала начинает увеличиваться высота передней симфизной части челюсти, которая становится выше задней. Первоначально нижний край челюсти более или менее полого поднимается вперед, но с возрастом появляется обособление лопастей, начинается сравнительно резкое отделение их приблизительно на уровне р3.
Предохранительные лопасти на нижней челюсти развиты очень сильно у взрослых самцов, у самок много слабее, у молодых (возможно, неполовозрелых) со сменяющимися зубами самцов совершенно отсутствуют. Из этого следует, что клыки становятся у самцов саблеобразными и вполне функционирующими только ко времени достижения половой зрелости. Таким образом, верхние саблеобразные клыки диноцератов развились как оружие борьбы между самцами, а в противном случае они появились бы на самых молодых стадиях, и зачатки предохранительных лопастей были бы выражены значительно раньше смены молочных зубов на постоянные.
Прорисовка частично реконструированного скелета продиноцера (Prodinoceras martyr) из гашатия местонахождения Наран-Булак в аймаке Умнеговь, Монголия.
Телосложение продиноцеров коренастое, с наивысшей точкой в пояснице. Шея мощная, относительно длинная. Туловище вытянутое и объемистое. Туловищные позвонки имеют удлиненные тела и большие остистые отростки, что свидетельствует о наличии развитой мускулатуры. Крестец состоит из 3 позвонков. Хвост длинный и толстый, составляющий около трети длины всего позвоночника. Представляется вероятным, что у продиноцеров сохранялся шерстный покров.
Лопатка сравнительно узкая и длинная, не имеет резко выраженной треугольной формы. В плечевом поясе присутствует ключица — элемент, не свойственный копытным. Таз длинный, узкий и выпрямленный. Конечности сильные, пятипалые, относительно короткие, передние короче, чем задние. Кости конечностей почти сплошные, выполнены губчатой массой, что свойственно, впрочем, почти всему скелету продиноцеров, за исключением некоторых частей черепа. Плечевая кость относительно короткая и мощная, с большой артикулярной головкой и хорошо развитыми буграми. Локтевая кость свободная, развита лучше лучевой. Передние конечности были сильно прогнуты в локтевых суставах. Бедренная кость с III-м вертелом. В заплюсне сращены II-я и III-я клиновидные кости; астрагал имеет шейку. Кисть не сохранилась, однако можно предположить, что передняя конечность были полупальцеходящей, задняя — стопоходящей. Подобно медвежьим, такие конечности приспособлены к передвижению по разнообразному грунту. Конечные фаланги в виде округлых наростов.

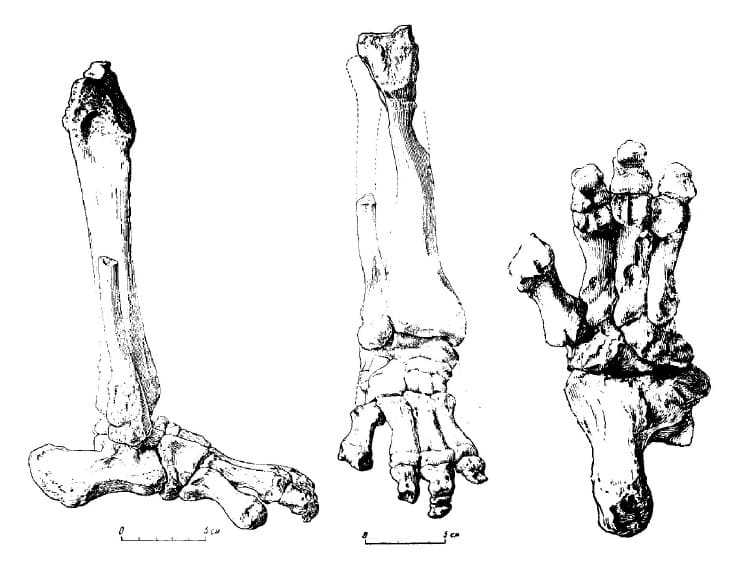
Скелет правой голени и стопы продиноцера (Prodinoceras martyr) (образец ПИН 534-69) из гашатия местонахождения Улан-Булак в аймаке Умнеговь, Монголия; снаружи, спереди и снизу.
Многочисленные гребни и шероховатости на костях передних конечностей указывают на сильное развитие мышц сгибающих, приводящих и поворачивающих их. Мощность мускулатуры плеча и локтя, значительное сгибание этих суставов были, безусловно, связаны с частыми наклонами передней части тела при добывании пищи, которую продиноцеры могли брать прямо с земли или разрывая почву. Строение костей предплечья говорит о том, что оно могло вращаться, хотя и в меньшей степени, чем у медведей. Развитие мускулатуры, вращающей и сгибающей кисть, свидетельствует в пользу того, что продиноцеры обладали способностью хватания при довольно значительной, по сравнению с более поздними диноцератами, подвижности кисти. Строение стопы заставляет считать, что она выдерживала большую нагрузку. Вероятно, эти животные могли подниматься на задние конечности, а также прочно цепляться при лазании по скалам.
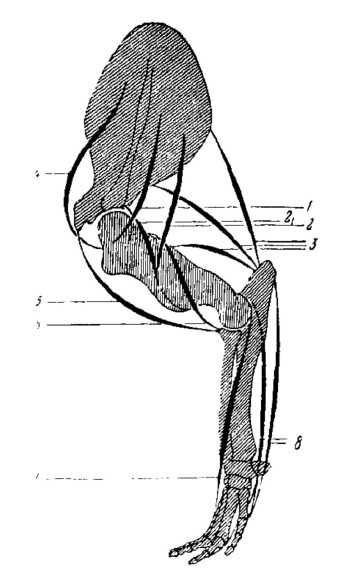
Схема мускулатуры передней конечности продиноцера (Prodinoceras martyr): 1 — m. infraspinatus; 2 — m. deltoideus, лопаточная порция; 21 — m. deltoideus, акромиальная порция; 3 — m. triceps brachii; 4 — m. supraspinatus; 5 — m. biceps brachii; 6 — m. brachialis; 7 — разгибатели кисти; 8 — сгибатели кисти.
Сравнение
Скелет продиноцеров характеризуется весьма специфическими чертами строения, обуславливающими его существенное отличие от прочих известных диноцератов.
Череп сходен с черепами других примитивных эвтериев, с большим сагиттальным гребнем, в противоположность плоскому черепу Gobiatherium или вогнутому в мозговой области у Uintatherium. Рогообразные выросты на верхней стороне отсутствуют, за исключением едва заметных возвышений над заглазничными отростками лобных костей. В противоположность уинтатериям, продиноцеры имеют верхние резцы. Нижние клыки у них большие, конические, а не резцеобразные, как у уинтатериев. От всех прочих родов диноцератов продиноцеры отличаются многими особенностями строения зубов.
В строении посткраниального скелета продиноцеров также отмечается значительное своеобразие. Так, уинтатерии имеют короткие поясничные позвонки, длина которых приблизительно равна высоте, снизу тела позвонков выпуклые; остистые отростки короткие, значительно короче длины тела позвонка, наклоненные назад. Те же позвонки продиноцеров обладают удлиненными телами (их длина больше высоты), снизу тела вогнутые, остистые отростки длинные, значительно длиннее тел, наклоненные вперед.
Особенно значительные отличия от уинтатериев прослеживаются в конечностях и их поясах. Лопатка продиноцеров значительно уже и длиннее, чем лопатка уинтатериев. Она менее слонообразна, не имеет столь резко выраженной треугольной формы. По сравнению с уинтатериями плечевая кость продиноцеров короче, ее сгибательный медиальный надмыщелок значительно сильнее. Локтевая и лучевая кости, наоборот, относительно тоньше и длиннее, чем таковые уинтатериев.
Крестец продиноцеров в противоположность уинтатериям состоит не из 4, а из 3 позвонков. Таз продиноцеров более длинный и выпрямленный в отличие от короткого слонообразного таза уинтатериев. Бедро уинтатериев имеет слонообразную форму: прямое, с головкой, располагающейся почти по оси кости, что понятно при большой массе животного и при необходимости поддержания его большого веса. По сравнению с уинтатериями бедро продиноцеров сильно отлично и больше приближается к бедру тех млекопитающих, у которых задние конечности несут не только опорную функцию, но имеют значительно большее разнообразие движений. Большое сходство в строении бедра продиноцеры имеют с некоторыми аминодонтами (Amynodontidae).
Особенно велики отличия в строении стопы продиноцеров от Uintatherium и Gobiatherium. Продиноцеры имеют стопоходную медведеобразную стопу в противоположность слонообразной уинтатериев. Таранная кость совершенно отлична от таранной кости уинтатериев и напоминает медвежью или даже человеческую, в некоторых отношениях занимая промежуточное место между ними. Пяточная кость длиннее пяточной кости человека, но короче медвежьей.
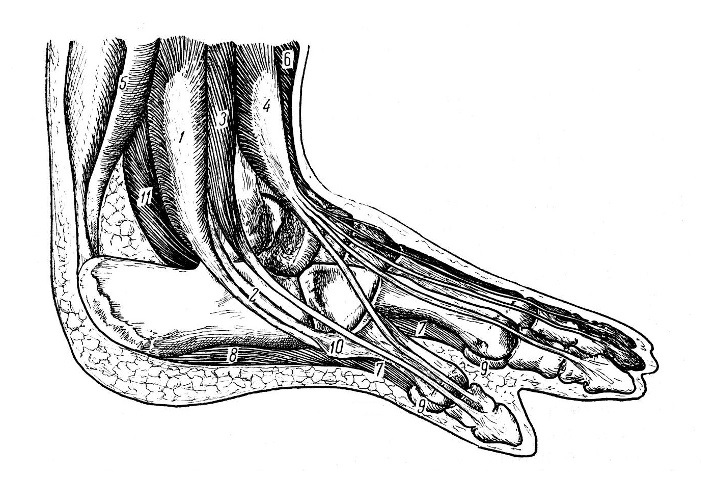
Реконструкция правой стопы продиноцера (Prodinoceras martyr); снаружи: 1 — длинный салоберцовый мускул; 2 — сухожилие короткого малоберцового мускула; 3 — боковой разгибатель пальцев; 4 — общий разгибатель пальцев; 5 — икроножная и камбаловидная мышцы; 6 — передний большеберцовый мускул; 7 — подошвенные межкостные мышцы; 8 — короткий сгибатель пальцев; 9 — сесамовидные кости; 10 — бугристость основания V-й плюсневой кости; 11 — брюшко длинного сгибателя пальцев.
В общем, посткраниальный скелет продиноцеров характеризуется рядом черт, свойственных, с одной стороны, медведям, с другой — безусловно, близко напоминающих пантодонтов типа Barylambda, а также некоторых наземных ленивцев. Несомненно, что продиноцеры обладали способностью подниматься и принимать вертикальное положение, хватаясь передними конечностями.