Происхождение
Терапсиды весьма разнообразны морфологически, поэтому для анализа вопроса их происхождения особый смысл имеют признаки их примитивных представителей. Среди этих признаков немало тех, которые свидетельствуют о родстве с пеликозаврами. Пеликозавры известны в основном из Северной Америки и Европы (несколько более поздних родов — из Южной Африки и России). Их наиболее полные ископаемые остатки найдены в раннепермской последовательности красноцветных слоев юго-запада США. Эти отложения продолжают накапливаться и в средней перми, но их фауна постепенно обедняется в результате возрастания аридности. Из среднепермских отложений описаны ископаемые остатки с некоторыми общими с терапсидами признаками, но они слишком фрагментарны, чтобы дать представление о переходе от одной группы к другой. Можно предположить, что древнейшие терапсиды обитали преимущественно в засушливых местностях за пределами болотистых ландшафтов, чем и объясняется их слабая представленность в палеонтологической летописи.
Древнейшие несомненные терапсиды известны из начала поздней перми Европейской части России. Здесь уже можно различить несколько групп, что говорит о длительном периоде предшествовавшей эволюции. Ранние терапсиды уже отчетливо продвинуты по сравнению с пеликозаврами, особенно чертами специализации посткраниального скелета. При этом они совпадают по времени с последним казеидовым пеликозавром из России — Ennatosaurus, но их остатки попадаются в иных местах; по-видимому, эти формы заселяли различные местообитания. В результате анатомических изменений и возможного разделения экологических ниш ранние терапсиды, вероятно, приспособились к иному образу жизни, чем пеликозавры; не исключено, что это связано и с их физиологическими особенностями. Такими различиями можно объяснить быструю радиацию терапсид, которые очень скоро достигли значительно большего разнообразия, чем их предки.
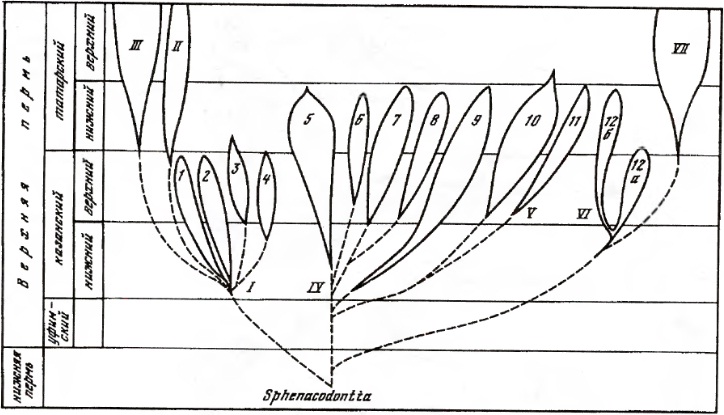
Традиционное филогенетическое древо терапсид (по Чудинову, 1983):
I — отряд Eotheriodontia, семейства: 1 — Biarmosuchidae, 2 — Eotitanosuchidae, 3 — Phthinosuchidae, 4 — Phthinosauridae; II — отряд Gorgonopia; III — отряд Eutheriodontia; надотряд Deinocephalia: IV — отряд Titanosuchia, семейства: 5 — Brithopodidae, 6 — Deuterosauridae, 7 — Anteosauridae, 8 — Titanosuchidae, 9 — Estemmenosuchidae; V — отряд Tapinocephalia, семейства: 10 — Tapinocephalidae, 11 — Styracocephalidae; надотряд Anomodontia: VI — отряд Venyukoviamorpha, семейство Venyukoviidae, подсемейства: 12a — Otsheriinae, 126 — Venyukoviinae; VII — отряд Dicynodontia.
Дж. Хопсон и Г. Баргхьюзен (1986 г.) присоединяются к авторам, особо выделяющим признаки, свидетельствующие о родстве терапсид с группой продвинутых пеликозавров — сфенакодонтами. У терапсид, по сравнению с пеликозаврами, заметно увеличено височное окно, но со сфенакодонтами они сходны и наличием на нижней челюсти отогнутой наружной пластинки угловой кости, и утратой скулового отростка квадратно-скуловой кости, замещенного отростком чешуйчатой кости, и увеличением высоты верхнечелюстной кости, отделяющей укороченную слезную кость от области ноздри; в передней части верхнечелюстной кости и в соответствующей области нижней челюсти развиваются клыки. Парокципитальный отросток удлиняется, и у более примитивных терапсид он направлен вниз и наружу. Ретроартикулярный отросток нижней челюсти у примитивных терапсид (особенно у фтинозухий и горгонопсий), как и у сфенакодонтов, массивен и отогнут вниз.
Из апоморфий собственно терапсид, помимо увеличения размеров височного окна, исследователи отмечают: утрату надвисочной кости; выраженный лицевой отросток септомаксиллярной кости, вклинивающийся между носовой и верхнечелюстной костями; более высокую, чем у пеликозавров, верхнечелюстную кость, которая в передней части, соединившись с носовой костью, отделяет слезную кость от ноздри и септомаксиллярной кости; удлинение восходящих отростков предчелюстных костей; образование вырезки в верхнем крае наружной пластинки угловой кости; уменьшение размеров межптеригоидной ямы; расширение сошника между хоанами. Для примитивных терапсид типично положение теменного отверстия на костном бугорке.
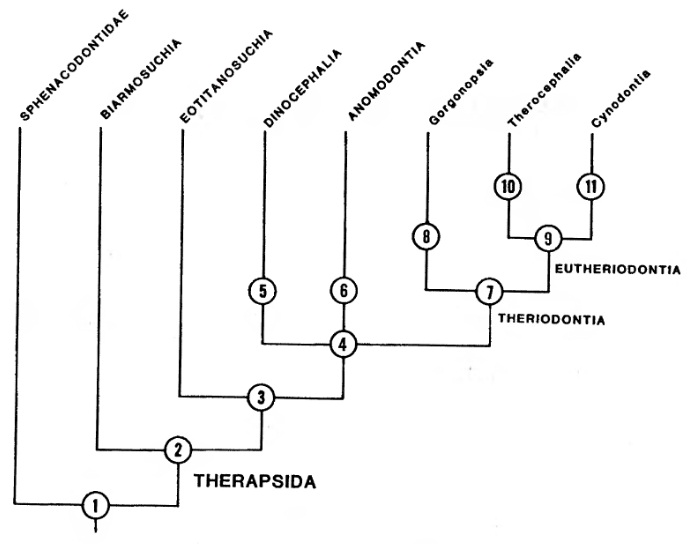
Кладограмма терапсид, включающая их сфенакодонтных предков (по Хопсону и Баргхьюзену, 1986).
Дж. Хопсон и Г. Баргхьюзен отмечали также появление на затылочной поверхности чешуйчатой кости желобка "наружного слухового прохода" и дальнейшее увеличение клыков, но первый признак не выражен у примитивных синапсид, а крупные клыки характерны, главным образом, для горгонопсий, тероцефалов и дицинодонтов. Дополнительно можно отметить тесную связь квадратно-скуловой кости с квадратной. В дальнейшей работе Дж. Хопсон (1991 г.) отмечает и некоторые признаки посткраниального скелета: сужение лопатки, укорочение межключицы, разрастание вперед подвздошной кости, стройность длинных костей конечностей, сигмовидную изогнутость бедренной кости, головка которой слегка выступает медиально, дисковидную форму "избыточных" для маммальной фаланговой формулы фаланг III-го и IV-го пальцев. Изменения в костях конечностей связаны с прогрессом локомоции. Имеются определенные признаки парасагиттальной постановки задних конечностей терапсид.
Конкретные переходные формы между сфенакодонтными пеликозаврами и терапсидами неизвестны. Dimetrodon и другие прогрессивные сфенакодонты (Sphenacodontia), очевидно, уже слишком специализированы (длинные остистые отростки, уменьшенное число зубов на предчелюстной кости), чтобы быть прямыми предками терапсид. Но более примитивный Haptodus, по мнению Р. Кэрролла (1987 г.), вполне подходит на эту роль. Согласно этой точке зрения, ведущая к терапсидам линия могла отделиться от самых примитивных сфенакодонтов, сходных с гаптодами, в любое время между поздним карбоном и средней пермью, на протяжении периода длительностью по меньшей мере 25 млн. лет.
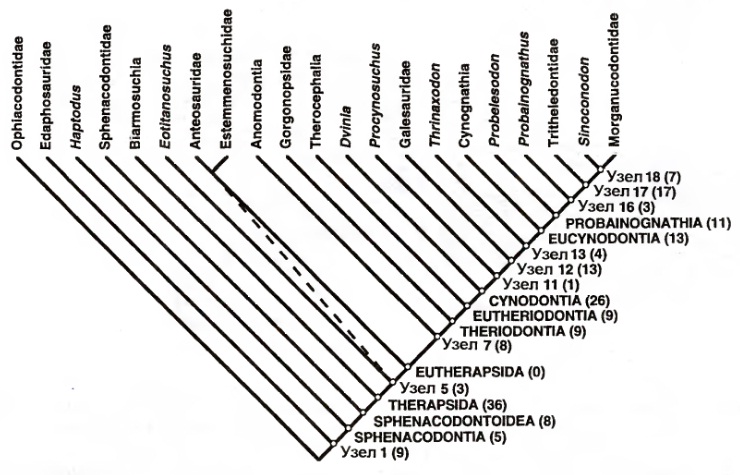
Популярная кладограмма тероморф (по Сидору и Хопсону, 1998).
В начале средней части перми выделяют так называемый провал Олсона, получивший свое название в честь американского палеонтолога Э. Олсона. Из этого промежутка времени, длившегося несколько миллионов лет, практически не известны фауны наземных позвоночных, хотя именно в это время должны были появиться первые терапсиды. Их самые древние надежные остатки происходят из слоев выше провала. Сам Олсон пытался его заполнить. В 1950–1960-х гг. из "среднепермских" слоях Техаса и Оклахомы, США, он описал довольно разнообразную фауну примитивных терапсид, которую он считал сходной с очерской фауной из России. Позднее оказалось, что описанные Э. Олсоном фрагментарные останки принадлежали казеидам и, возможно, другим пеликозаврам. Это кажется странным, учитывая тот факт, что Э. Олсон был одним из ведущих специалистов по казеидам. Вероятно, ошибка Олсона может объясняться психологическими причинами — он был вдохновлен открытием очерской фауны и верил в то, что в переходных "среднепермских" слоях Северной Америки удастся обнаружить предшественников русских терапсид. Известно, что Э. Олсон дружил с И.А. Ефремовым (они долгие годы переписывались и обменивались научной информацией). Возможно, что тут какую-то роль сыграло дружеское научное соперничество.
М. Лорин и Р. Рейс (1990, 1996 гг.) считали наиболее близким к предкам терапсид некрупного (череп длиной менее 10 см) синапсида Tetraceratops из конца ранней перми Техаса, жившего рядом с различными пеликозаврами. А. Ромер и Л. Прайс (1940 г.) условно относили этого курьезное животное с 6 рогообразными выростами на голове к эотиридидам (Eothyrididae). Верхнечелюстная кость у Tetraceratops невысокая и не отделяет слезную кость от ноздри, септомаксиллярная кость не имеет длинного, характерного для терапсид лицевого отростка. Но размеры височного окна у Tetraceratops увеличены, в передней части верхнечелюстной кости развиты клыки. М. Лорин и Р. Рейз показали, что у Tetraceratops имеется отогнутая пластинка угловой кости, зубы на эктоптеригоидах отсутствуют, а квадратно-скуловая кость маленькая. М. Лорин и Р. Рейс отнесли Tetraceratops к базальным терапсидам, хотя, вероятно, правильнее считать его аберрантным сфенакодонтом, сохранявшим невысокую верхнечелюстную кость.
Не исключено также, что реликтами подобных переходных форм могут являться Nikkasaurus из Западного Приуралья и Raranimus из Китая, датирующиеся началом средней перми. Их черепа довольно сильно напоминают черепа сфенакодонтов, хотя наблюдаются и существенные отличия. Однако сами эти некрупные хищники предками терапсид быть не могли. В фаунах, из которых они происходят, рядом с ними жили и настоящие, хотя весьма примитивные терапсиды. Остатки других наиболее ранних из известных терапсид происходят из Южной Африки и Южной Америки.