Терапсиды — Therapsida
Клада Амниоты (Amniota)
Клада Синапсидообразные (Synapsidomorpha)
Класс †Синапсиды (Synapsida)
Подкласс †Терапсиды (Therapsida)
Терапсиды (Therapsida Broom, 1905) — группа, объединяющая высших синапсид, условно принимаемая здесь в качестве подкласса. Известны начиная со средней перми до раннего мела. Доминируют в большинстве известных позднепермских фаун. Крайне разнообразны по величине, пищевым адаптациям и местообитаниям. В кладистических классификациях терапсиды не являются вымершими, так как млекопитающие, считающиеся их потомками, существуют до сих пор.
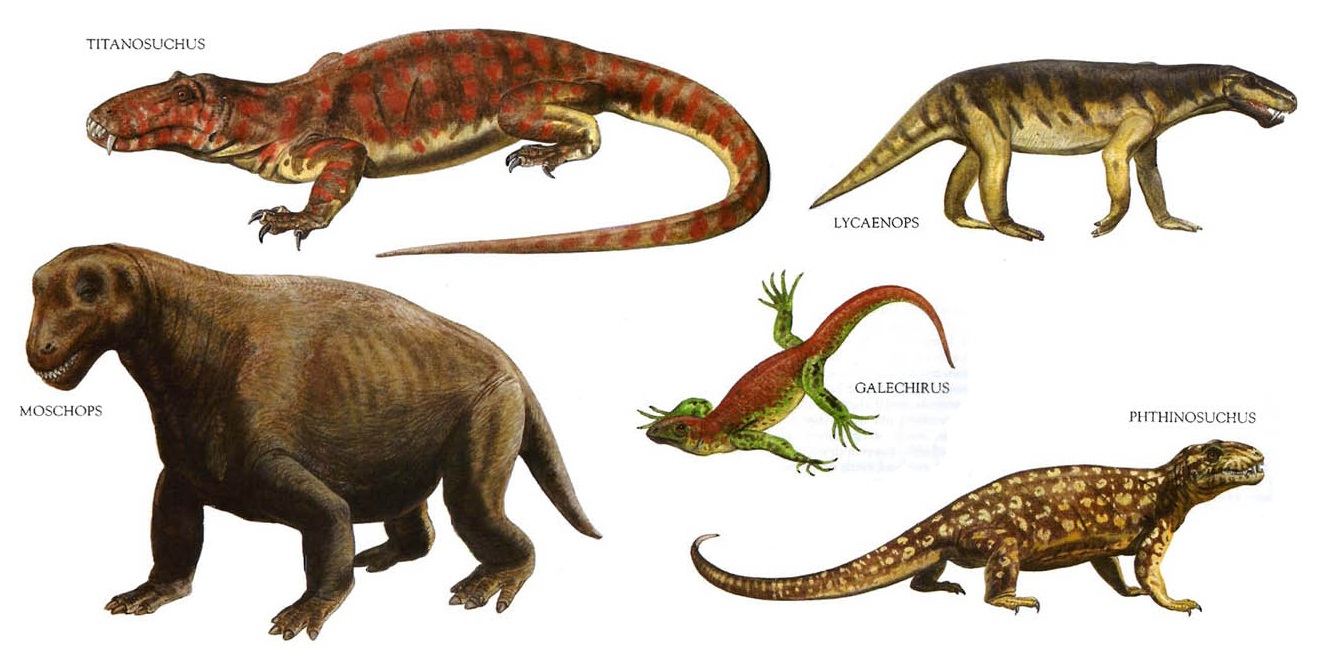
Разнообразие терапсид.
Ссылки
Маммализация териодонтов (Википедия) перейди
Терапсиды (Википедия) перейти
Териодонты (Википедия) перейти
Eutherapsida (Википедия) перейти
Литература
Богданов Д. В. Звероящеры и другие пермские монстры. — М.: Фитон XXI, 2018
Иванов А. О., Черепанов Г. О. Ископаемые высшие позвоночные. — СПб.: Издательство Санкт-Петербургского университета, 2001
Ивахненко М. Ф. Тетраподы Восточно-Европейского плаката — позднепалеозойского территориально-природного комплекса. — Пермь, 2001
Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Ч. 1 / Отв. ред. Ивахненко М. Ф. и Курочкин Е. Н. — М.: ГЕОС, 2008
Кэрролл Р. Палеонтология и эволюция позвоночных. В 3 т. Т. 2 — М.: Мир, 1993
Основы палеонтологии. В 15 т. Т. 12: Земноводные, пресмыкающиеся, птицы / Гл. ред. Орлов Ю. А. — М.: Наука, 1964
Протеро Д. Отпечатки жизни. 25 шагов эволюции и вся история планеты. — СПб.: Питер, 2016
Ромер А. Ш. Палеонтология позвоночных. — М. — Л.: Государственное научно-техническое издательство нефтяной и горно-топливной литературы, 1939
Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М.: ГЕОС, 2009